
Effects of a Cc2d1a/Freud-1 Knockdown in the Hippocampus on Behavior, the Serotonin System, and BDNF

 ,
,
Abstract
:1. Introduction
2. Results
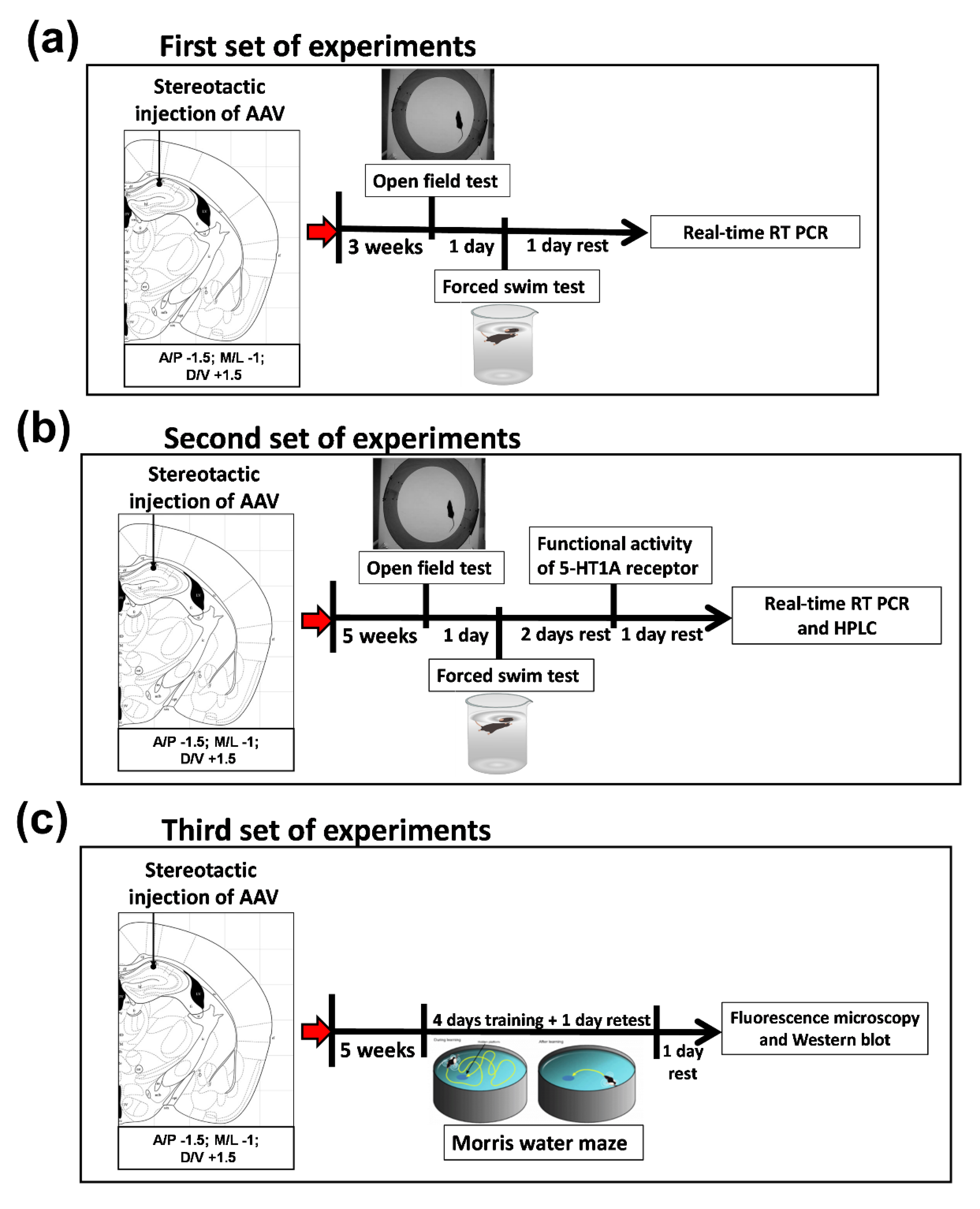
2.1. The First Set of Experiments (3 Week of Recovery after AAV Administration)
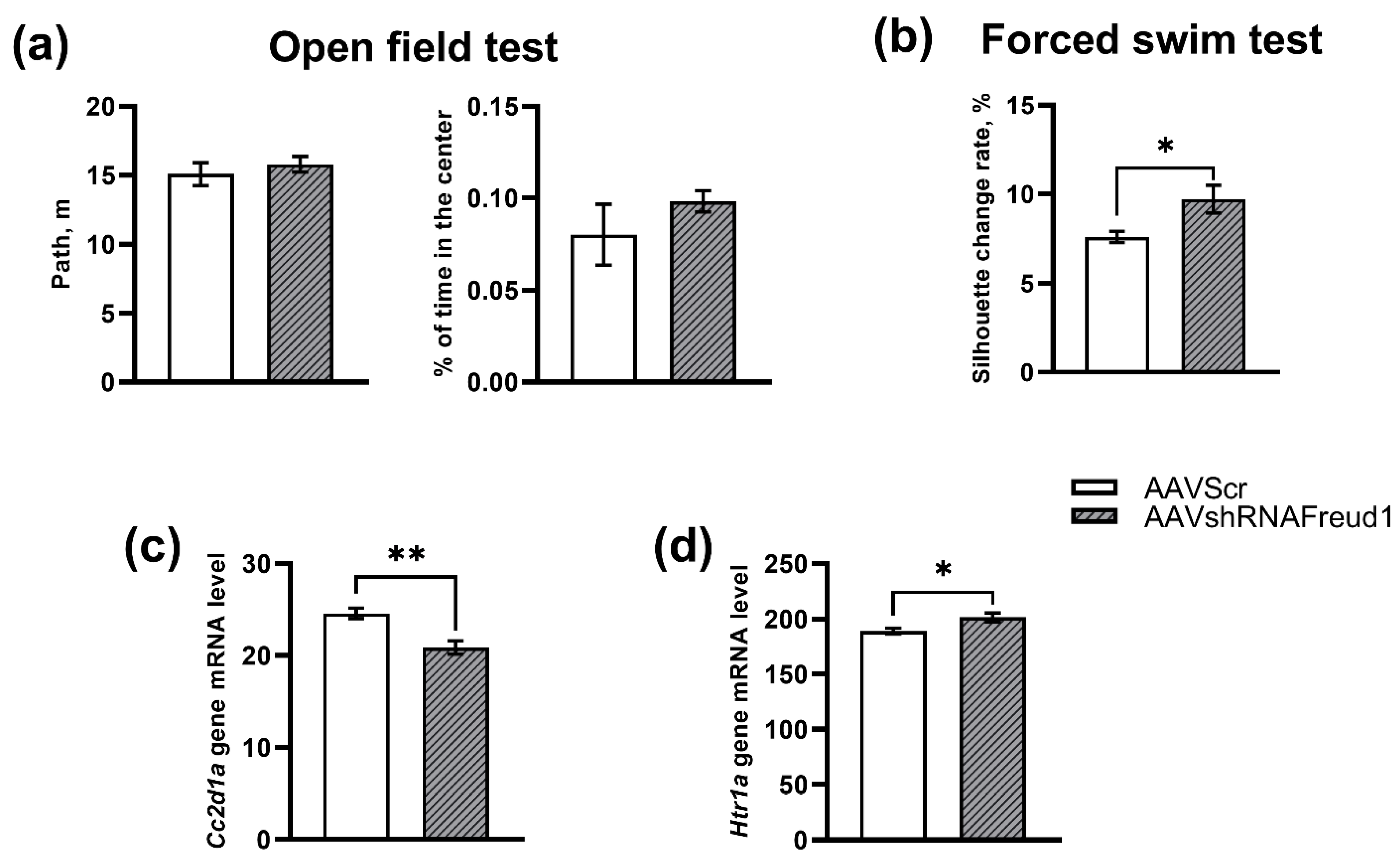
2.1.1. The Open Field Test
2.1.2. The Forced Swim Test
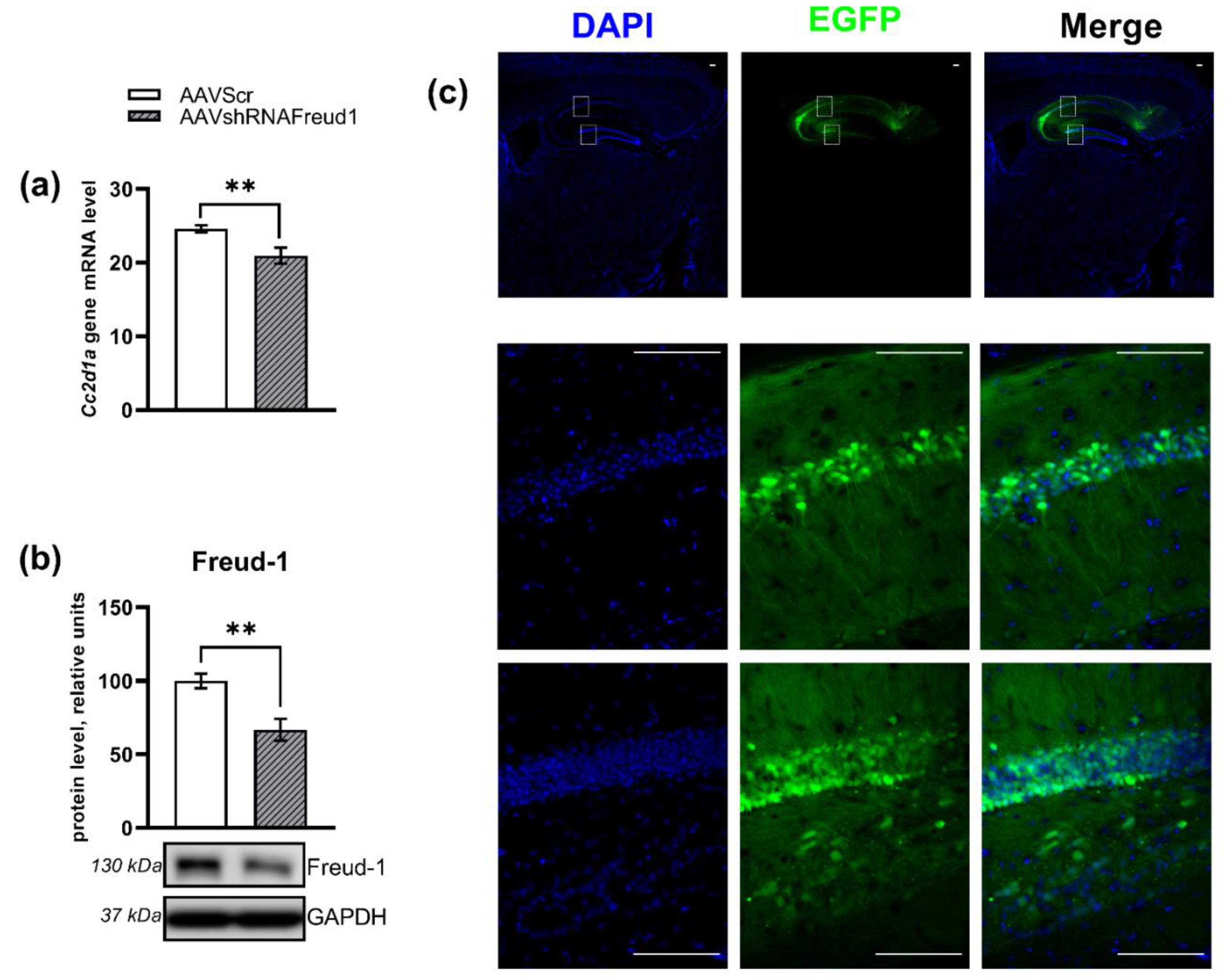
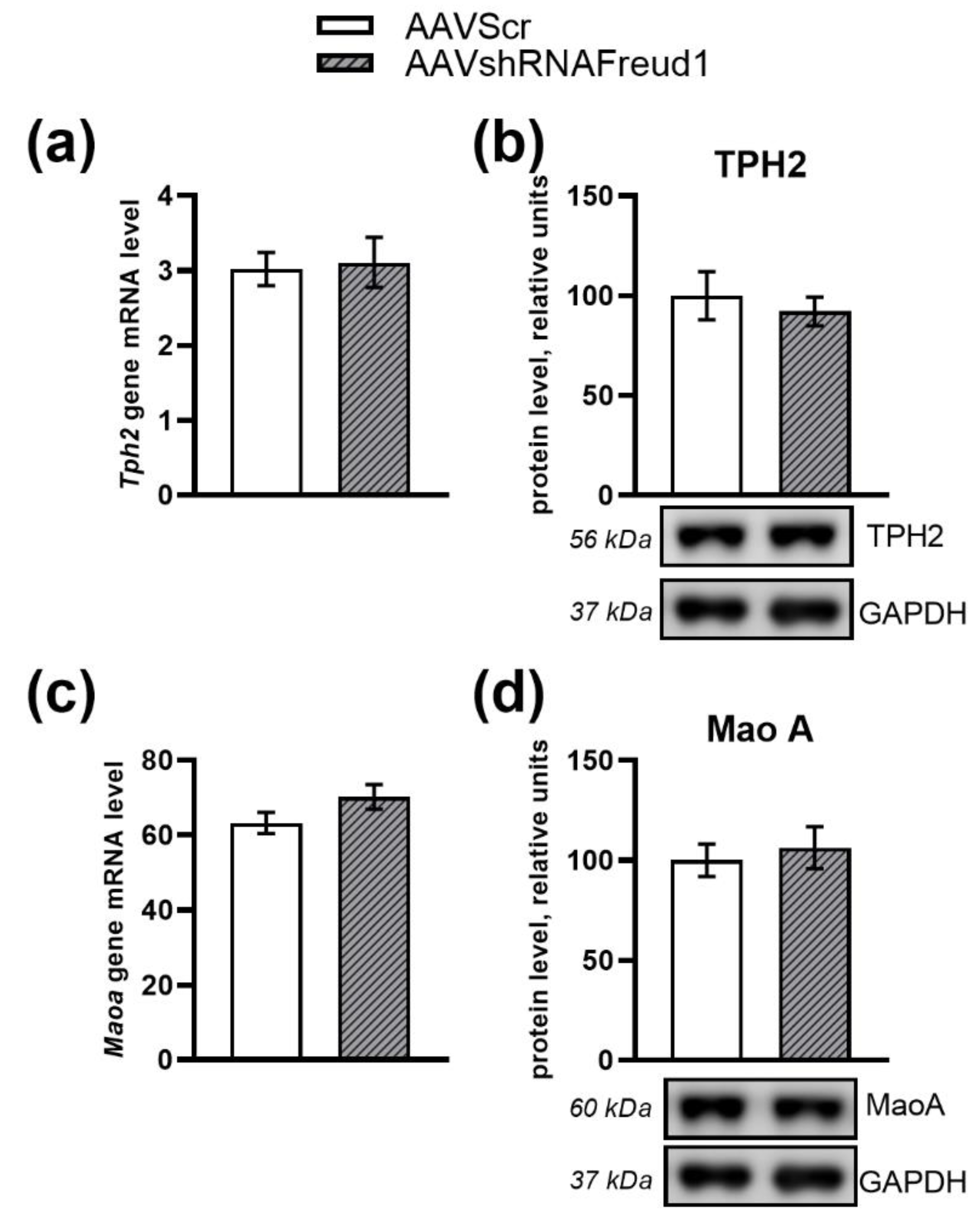
2.1.3. mRNA Levels
2.2. The Second and Third Sets of Experiments (5 Week of Recovery after AAV Administration)
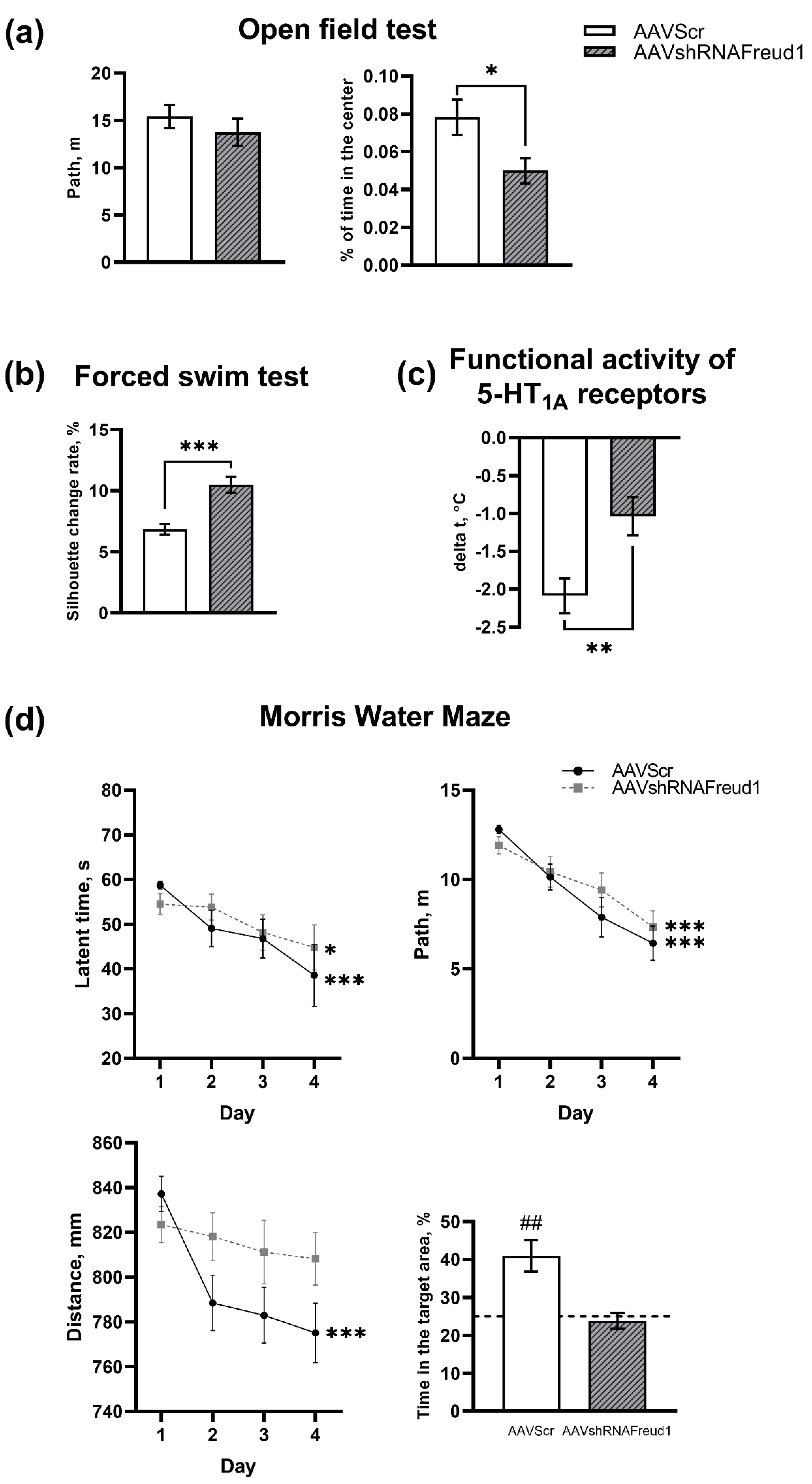
2.2.1. The Open Field Test
2.2.2. The Forced Swim Test
2.2.3. Pharmacological Analysis of the 5-HT1A Receptor Functional Activity
2.2.4. The Morris Water Maze Test
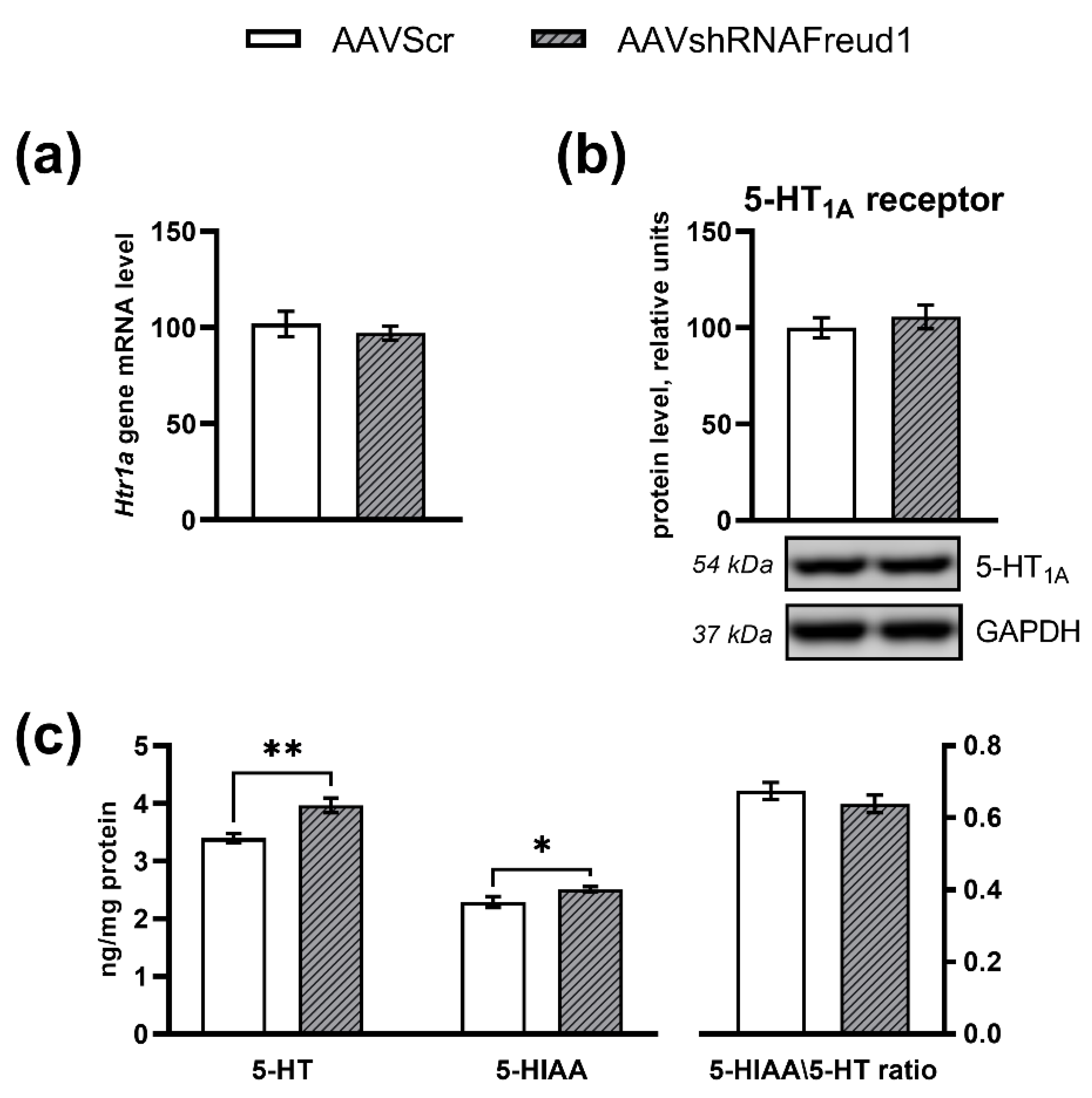
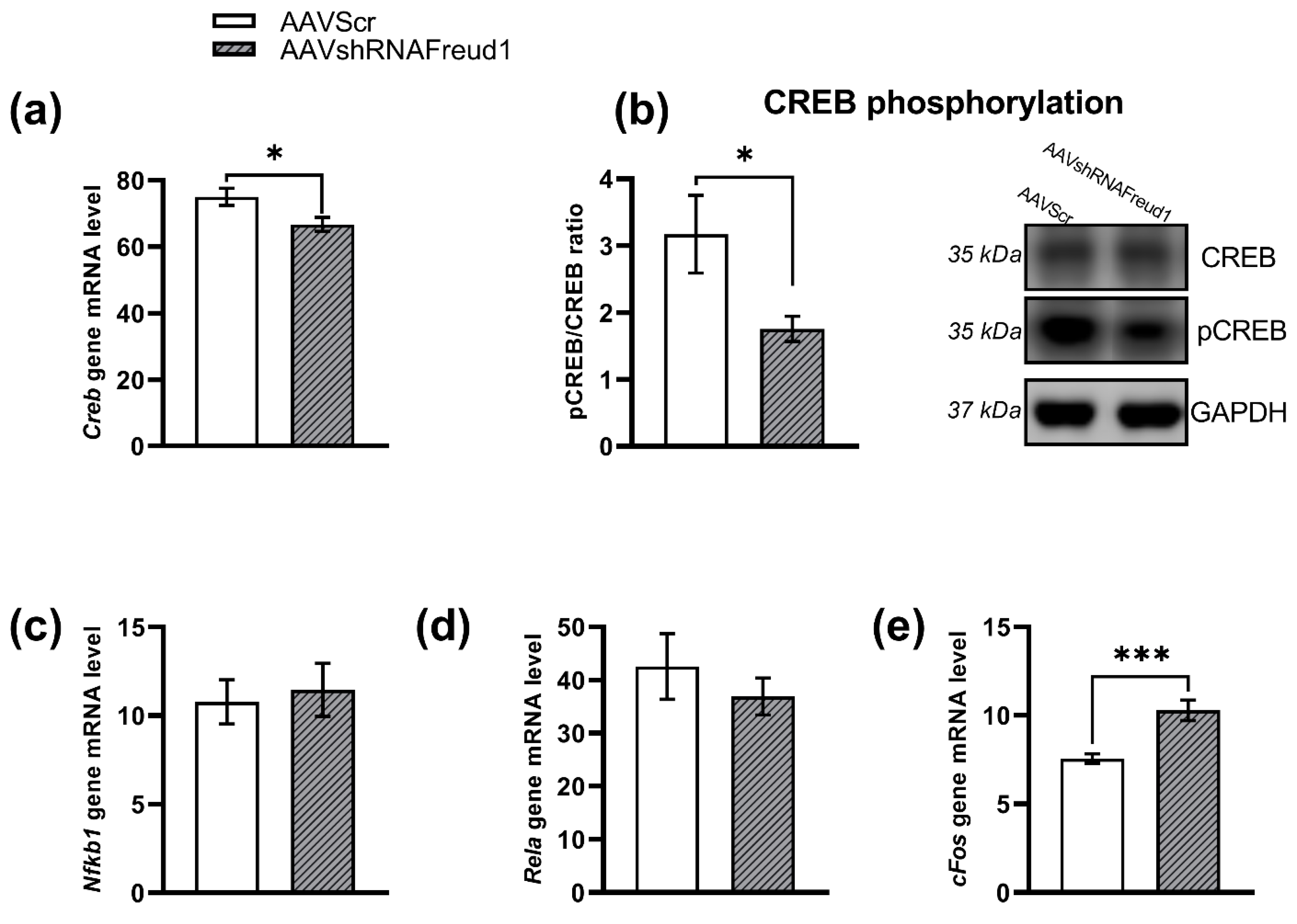
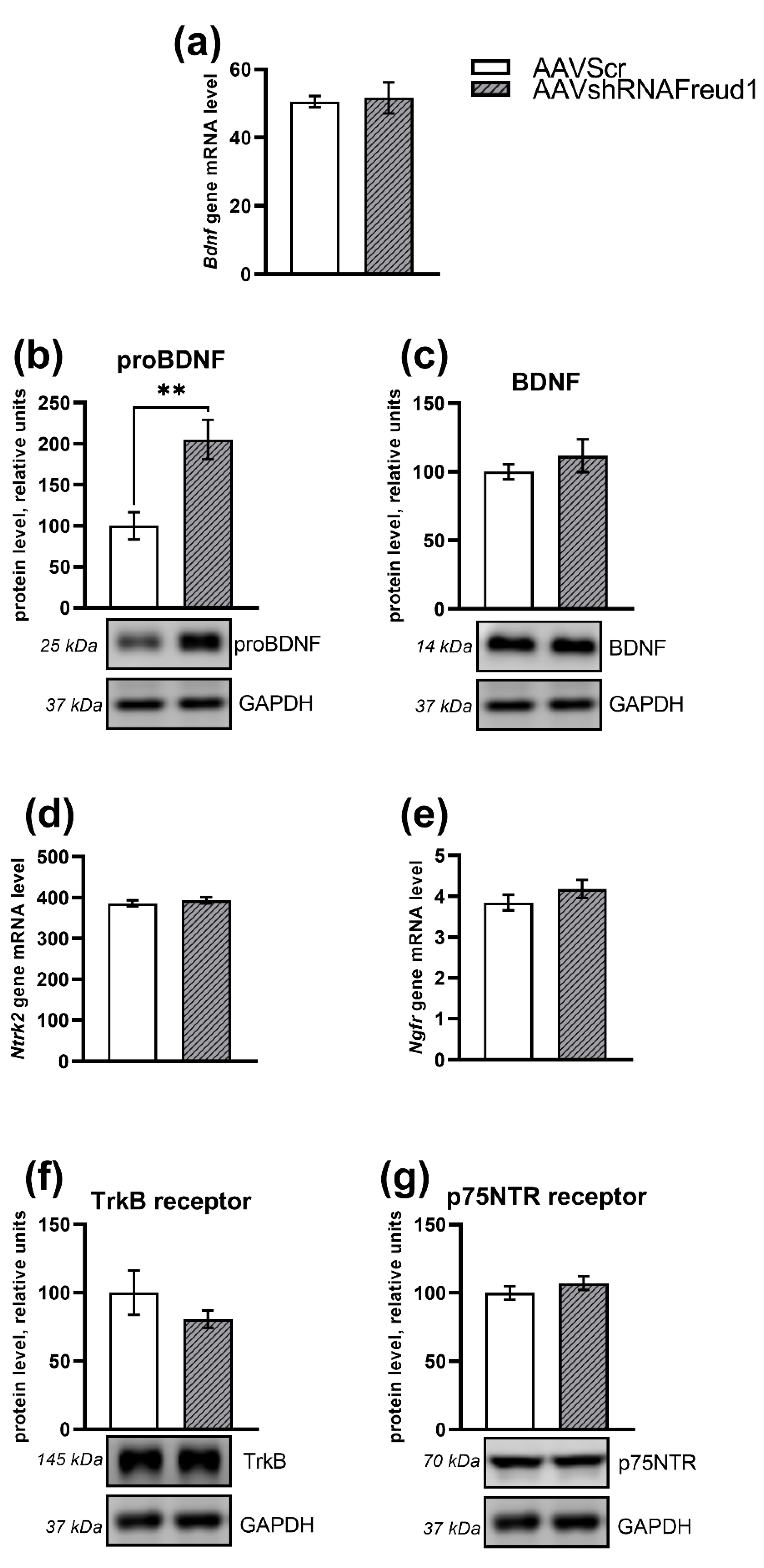
2.2.5. mRNA, Protein, and Monoamine Levels
3. Discussion
4. Materials and Methods
4.1. Experimental Design
4.1.1. Animals
4.1.2. Plasmids
4.1.3. Cell Culture and Transfection
4.1.4. AAV Production
4.1.5. Primary Neuronal Culture
4.1.6. Stereotactic Injection into the Hippocampus
4.1.7. Hippocampus Isolation
4.1.8. The Open Field Test
4.1.9. The Forced Swim Test
4.1.10. Pharmacological Analysis of 5-HT1A Receptor Functional Activity
4.1.11. The Morris Water Maze Test
4.1.12. Fluorescence Microscopy
4.1.13. qPCR
4.1.14. HPLC
4.1.15. Western Blot
4.1.16. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Harro, J.; Oreland, L. Depression as a spreading neuronal adjustment disorder. Eur. Neuropsychopharmacol. 1996, 6, 207–223. [Google Scholar] [CrossRef]
- Duman, R.S.; Heninger, G.R.; Nestler, E.J. A Molecular and Cellular Theory of Depression. Arch. Gen. Psychiatry 1997, 54, 597–606. [Google Scholar] [CrossRef]
- Jans, L.A.W.; Riedel, W.J.; Markus, C.R.; Blokland, A. Serotonergic vulnerability and depression: Assumptions, experimental evidence and implications. Mol. Psychiatry 2006, 12, 522–543. [Google Scholar] [CrossRef]
- Popova, N.K.; Naumenko, V.S. 5-HT1A receptor as a key player in the brain 5-HT system. Rev. Neurosci. 2013, 24, 1–14. [Google Scholar] [CrossRef]
- Barnes, N.M.; Sharp, T. A review of central 5-HT receptors and their function. Neuropharmacology 1999, 38, 1083–1152. [Google Scholar] [CrossRef]
- Albert, P.R.; Le François, B.; Millar, A.M. Transcriptional dysregulation of 5-HT1A autoreceptors in mental illness. Mol. Brain 2011, 4, 21. [Google Scholar] [CrossRef] [Green Version]
- Fuller, R.W. Role of serotonin in therapy of depression and related disorders. J. Clin. Psychiatry 1991, 52, 52–57. [Google Scholar] [PubMed]
- Heisler, L.K.; Chu, H.-M.; Brennan, T.J.; Danao, J.A.; Bajwa, P.; Parsons, L.H.; Tecott, L.H. Elevated anxiety and antidepressant-like responses in serotonin 5-HT1A receptor mutant mice. Proc. Natl. Acad. Sci. USA 1998, 95, 15049–15054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Overstreet, D.H.; Commissaris, R.C.; De La Garza, R., 2nd; File, S.E.; Knapp, D.J.; Seiden, L.S. Involvement of 5-HT1A receptors in animal tests of anxiety and depression: Evidence from genetic models. Stress 2003, 6, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Ramboz, S.; Oosting, R.; Amara, D.A.; Kung, H.F.; Blier, P.; Mendelsohn, M.; Mann, J.J.; Brunner, D.; Hen, R. Serotonin receptor 1A knockout: An animal model of anxiety-related disorder. Proc. Natl. Acad. Sci. USA 1998, 95, 14476–14481. [Google Scholar] [CrossRef] [Green Version]
- Richardson-Jones, J.W.; Craige, C.P.; Guiard, B.P.; Stephen, A.; Metzger, K.L.; Kung, H.F.; Gardier, A.M.; Dranovsky, A.; David, D.; Beck, S.G.; et al. 5-HT1A Autoreceptor Levels Determine Vulnerability to Stress and Response to Antidepressants. Neuron 2010, 65, 40–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maes, M.; Meltzer, H.Y. The serotonin hypothesis of major depression. In Psychopharmacology: The Fourth Generation of Progress; Bloom, E.E., Kupfer, N.N., Eds.; Raven Press: New York, NY, USA, 1995; pp. 933–944. [Google Scholar]
- Arango, V.; Huang, Y.-Y.; Underwood, M.; Mann, J. Genetics of the serotonergic system in suicidal behavior. J. Psychiatr. Res. 2003, 37, 375–386. [Google Scholar] [CrossRef]
- Stockmeier, C.A.; Shapiro, L.A.; Dilley, G.E.; Kolli, T.N.; Friedman, L.; Rajkowska, G. Increase in Serotonin-1A Autoreceptors in the Midbrain of Suicide Victims with Major Depression—Postmortem Evidence for Decreased Serotonin Activity. J. Neurosci. 1998, 18, 7394–7401. [Google Scholar] [CrossRef] [Green Version]
- Lowther, S.; De Paermentier, F.; Cheetham, S.C.; Crompton, M.; Katona, C.L.; Horton, R.W. 5-HT1A Receptor binding sites in post-mortem brain samples from depressed suicides and controls. J. Affect. Disord. 1997, 42, 199–207. [Google Scholar] [CrossRef]
- Albert, P.; Lemonde, S. 5-HT1A Receptors, Gene Repression, and Depression: Guilt by Association. Neuroscientist 2004, 10, 575–593. [Google Scholar] [CrossRef]
- Shively, C.A.; Friedman, D.P.; Gage, H.D.; Bounds, M.C.; Brown-Proctor, C.; Blair, J.B.; Henderson, J.A.; Smith, M.A.; Buchheimer, N. Behavioral Depression and Positron Emission Tomography–Determined Serotonin 1A Receptor Binding Potential in Cynomolgus Monkeys. Arch. Gen. Psychiatry 2006, 63, 396–403. [Google Scholar] [CrossRef] [Green Version]
- Sargent, P.A.; Kjaer, K.H.; Bench, C.J.; Rabiner, E.A.; Messa, C.; Meyer, J.; Gunn, R.N.; Grasby, P.M.; Cowen, P.J. Brain serotonin1A receptor binding measured by positron emission tomography with [11C]WAY-100635: Effects of depression and antidepressant treatment. Arch. Gen. Psychiatry 2000, 57, 174–180. [Google Scholar] [CrossRef] [Green Version]
- Bhagwagar, Z.; Rabiner, E.A.; Sargent, P.A.; Grasby, P.M.; Cowen, P.J. Persistent reduction in brain serotonin1A receptor binding in recovered depressed men measured by positron emission tomography with [11C]WAY-100635. Mol. Psychiatry 2004, 9, 386–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moses-Kolko, E.L.; Wisner, K.; Price, J.C.; Berga, S.L.; Drevets, W.C.; Hanusa, B.H.; Loucks, T.; Meltzer, C.C. Serotonin 1A receptor reductions in postpartum depression: A positron emission tomography study. Fertil. Steril. 2008, 89, 685–692. [Google Scholar] [CrossRef] [Green Version]
- Stockmeier, C.A.; Howley, E.; Shi, X.; Sobanska, A.; Clarke, G.; Friedman, L.; Rajkowska, G. Antagonist but not agonist labeling of serotonin-1A receptors is decreased in major depressive disorder. J. Psychiatr. Res. 2009, 43, 887–894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neumeister, A.; Bain, E.; Nugent, A.C.; Carson, R.E.; Bonne, O.; Luckenbaugh, D.A.; Eckelman, W.; Herscovitch, P.; Charney, D.S.; Drevets, W.C. Reduced serotonin type 1A receptor binding in panic disorder. J. Neurosci. 2004, 24, 589–591. [Google Scholar] [CrossRef] [Green Version]
- Sullivan, G.M.; Oquendo, M.A.; Simpson, N.; Van Heertum, R.L.; Mann, J.J.; Parsey, R.V. Brain Serotonin1A Receptor Binding in Major Depression Is Related to Psychic and Somatic Anxiety. Biol. Psychiatry 2005, 58, 947–954. [Google Scholar] [CrossRef]
- Lanzenberger, R.R.; Mitterhauser, M.; Spindelegger, C.; Wadsak, W.; Klein, N.; Mien, L.-K.; Holik, A.; Attarbaschi, T.; Mossaheb, N.; Sacher, J.; et al. Reduced Serotonin-1A Receptor Binding in Social Anxiety Disorder. Biol. Psychiatry 2007, 61, 1081–1089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drevets, W.C.; Thase, M.E.; Moses-Kolko, E.L.; Price, J.; Frank, E.; Kupfer, D.J.; Mathis, C. Serotonin-1A receptor imaging in recurrent depression: Replication and literature review. Nucl. Med. Biol. 2007, 34, 865–877. [Google Scholar] [CrossRef] [Green Version]
- Boldrini, M.; Underwood, M.; Mann, J.J.; Arango, V. Serotonin-1A autoreceptor binding in the dorsal raphe nucleus of depressed suicides. J. Psychiatr. Res. 2008, 42, 433–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ou, X.-M.; Jafar-Nejad, H.; Storring, J.M.; Meng, J.-H.; Lemonde, S.; Albert, P. Novel Dual Repressor Elements for Neuronal Cell-specific Transcription of the Rat 5-HT1A Receptor Gene. J. Biol. Chem. 2000, 275, 8161–8168. [Google Scholar] [CrossRef] [Green Version]
- Ou, X.-M.; Lemonde, S.; Jafar-Nejad, H.; Bown, C.D.; Goto, A.; Rogaeva, A.; Albert, P.R. Freud-1: A Neuronal Calcium-Regulated Repressor of the 5-HT1A Receptor Gene. J. Neurosci. 2003, 23, 7415–7425. [Google Scholar] [CrossRef] [Green Version]
- Zhao, M.; Raingo, J.; Chen, Z.J.; Kavalali, E.T. Cc2d1a, a C2 domain containing protein linked to nonsyndromic mental retardation, controls functional maturation of central synapses. J. Neurophysiol. 2011, 105, 1506–1515. [Google Scholar] [CrossRef] [Green Version]
- Al-Tawashi, A.; Jung, S.Y.; Liu, D.; Su, B.; Qin, J. Protein Implicated in Nonsyndromic Mental Retardation Regulates Protein Kinase A (PKA) Activity. J. Biol. Chem. 2012, 287, 14644–14658. [Google Scholar] [CrossRef] [Green Version]
- Chen, K.R.; Chang, C.H.; Huang, C.Y.; Lin, C.Y.; Lin, W.Y.; Lo, Y.C.; Yang, C.Y.; Hsing, E.W.; Chen, L.F.; Shih, S.R.; et al. TBK1-associated protein in endolysosomes (TAPE)/CC2D1A is a key regulator linking RIG-I-like receptors to antiviral immunity. J. Biol. Chem. 2012, 287, 32216–32221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vahid-Ansari, F.; Daigle, M.; Manzini, M.C.; Tanaka, K.F.; Hen, R.; Geddes, S.D.; Beique, J.C.; James, J.; Merali, Z.; Albert, P.R. Abrogated Freud-1/Cc2d1a Repression of 5-HT1A Autoreceptors Induces Fluoxetine-Resistant Anxiety/Depression-Like Behavior. J. Neurosci. 2017, 37, 11967–11978. [Google Scholar] [CrossRef] [Green Version]
- Albert, P.; Lembo, P.; Storring, J.M.; Charest, A.; Saucier, C. The 5-HT1A receptor: Signaling, desensitization, and gene transcription. Neuropsychopharmacology 1996, 14, 19–25. [Google Scholar] [CrossRef]
- Albert, P.R.; Francois, B.L. Modifying 5-HT1A Receptor Gene Expression as a New Target for Antidepressant Therapy. Front. Neurosci. 2010, 4, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castrén, E.; Rantamäki, T. The role of BDNF and its receptors in depression and antidepressant drug action: Reactivation of developmental plasticity. Dev. Neurobiol. 2010, 70, 289–297. [Google Scholar] [CrossRef]
- Sun, L.-J.; Zhang, L.-M.; Liu, D.; Xue, R.; Liu, Y.-Q.; Li, L.; Guo, Y.; Shang, C.; Yao, J.-Q.; Zhang, Y.-Z.; et al. The faster-onset antidepressant effects of hypidone hydrochloride (YL-0919). Metab. Brain Dis. 2019, 34, 1375–1384. [Google Scholar] [CrossRef]
- Lu, B.; Pang, P.T.; Woo, N.H. The yin and yang of neurotrophin action. Nat. Rev. Neurosci. 2005, 6, 603–614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benarroch, E.E. Brain-derived neurotrophic factor: Regulation, effects, and potential clinical relevance. Neurology 2015, 84, 1693–1704. [Google Scholar] [CrossRef]
- Kenchappa, R.S.; Tep, C.; Korade, Z.; Urra, S.; Bronfman, F.C.; Yoon, S.O.; Carter, B.D. p75 Neurotrophin Receptor-mediated Apoptosis in Sympathetic Neurons Involves a Biphasic Activation of JNK and Up-regulation of Tumor Necrosis Factor-α-converting Enzyme/ADAM17. J. Biol. Chem. 2010, 285, 20358–20368. [Google Scholar] [CrossRef] [Green Version]
- Deinhardt, K.; Chao, M. Shaping neurons: Long and short range effects of mature and proBDNF signalling upon neuronal structure. Neuropharmacology 2014, 76, 603–609. [Google Scholar] [CrossRef] [Green Version]
- Popova, N.K.; Ilchibaeva, T.V.; Naumenko, V.S. Neurotrophic factors (BDNF and GDNF) and the serotonergic system of the brain. Biochemistry 2017, 82, 308–317. [Google Scholar] [CrossRef] [PubMed]
- Popova, N.K.; Naumenko, V.S. Neuronal and behavioral plasticity: The role of serotonin and BDNF systems tandem. Expert Opin. Ther. Targets 2019, 23, 227–239. [Google Scholar] [CrossRef]
- Oeckinghaus, A.; Ghosh, S. The NF-kappaB family of transcription factors and its regulation. Cold Spring Harb. Perspect. Biol. 2009, 1, a000034. [Google Scholar] [CrossRef]
- Zamarbide, M.; Mossa, A.; Muñoz-Llancao, P.; Wilkinson, M.K.; Pond, H.L.; Oaks, A.W.; Manzini, M.C. Male-Specific cAMP Signaling in the Hippocampus Controls Spatial Memory Deficits in a Mouse Model of Autism and Intellectual Disability. Biol. Psychiatry 2019, 85, 760–768. [Google Scholar] [CrossRef] [PubMed]
- Kraemer, B.R.; Yoon, S.O.; Carter, B.D. The Biological Functions and Signaling Mechanisms of the p75 Neurotrophin Receptor. Mediat. Drugs Gastrointest. Motil. I 2014, 220, 121–164. [Google Scholar]
- Wissink, S.; Meijer, O.; Pearce, D.; van der Burg, B.; van der Saag, P.T. Regulation of the Rat Serotonin-1A Receptor Gene by Corticosteroids. J. Biol. Chem. 2000, 275, 1321–1326. [Google Scholar] [CrossRef] [Green Version]
- Ying, S.-W.; Futter, M.; Rosenblum, K.; Webber, M.J.; Hunt, S.; Bliss, T.V.P.; Bramham, C. Brain-Derived Neurotrophic Factor Induces Long-Term Potentiation in Intact Adult Hippocampus: Requirement for ERK Activation Coupled to CREB and Upregulation ofArcSynthesis. J. Neurosci. 2002, 22, 1532–1540. [Google Scholar] [CrossRef] [Green Version]
- Overstreet, D.H. Modeling Depression in Animal Models. Psychiatr. Disord. 2012, 829, 125–144. [Google Scholar]
- Khotskin, N.V.; Plyusnina, A.V.; Kulikova, E.A.; Bazhenova, E.Y.; Fursenko, D.V.; Sorokin, I.E.; Kolotygin, I.; Mormede, P.; Terenina, E.E.; Shevelev, O.B.; et al. On association of the lethal yellow (A(Y)) mutation in the agouti gene with the alterations in mouse brain and behavior. Behav. Brain Res. 2019, 359, 446–456. [Google Scholar] [CrossRef]
- Santarelli, L.; Saxe, M.; Gross, C.; Surget, A.; Battaglia, F.; Dulawa, S.; Weisstaub, N.; Lee, J.; Duman, R.; Arancio, O.; et al. Requirement of Hippocampal Neurogenesis for the Behavioral Effects of Antidepressants. Science 2003, 301, 805–809. [Google Scholar] [CrossRef] [Green Version]
- Porsolt, R.D.; Bertin, A.; Jalfre, M. “Behavioural despair” in rats and mice: Strain differences and the effects of imipramine. Eur. J. Pharmacol. 1978, 51, 291–294. [Google Scholar] [CrossRef]
- Oaks, A.W.; Zamarbide, M.; Tambunan, D.E.; Santini, E.; Di Costanzo, S.; Pond, H.L.; Johnson, M.W.; Lin, J.; Gonzalez, D.; Boehler, J.F.; et al. Cc2d1a Loss of Function Disrupts Functional and Morphological Development in Forebrain Neurons Leading to Cognitive and Social Deficits. Cereb. Cortex 2017, 27, 1670–1685. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.-Y.; Yu, T.-H.; Wen, W.-L.; Ling, P.; Hsu, K.-S. Conditional Deletion of CC2D1A Reduces Hippocampal Synaptic Plasticity and Impairs Cognitive Function through Rac1 Hyperactivation. J. Neurosci. 2019, 39, 4959–4975. [Google Scholar] [CrossRef]
- Fanselow, M.S.; Dong, H.-W. Are the Dorsal and Ventral Hippocampus Functionally Distinct Structures? Neuron 2010, 65, 7–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pothuizen, H.H.J.; Zhang, W.-N.; Jongen-Relo, A.L.; Feldon, J.; Yee, B.K. Dissociation of function between the dorsal and the ventral hippocampus in spatial learning abilities of the rat: A within-subject, within-task comparison of reference and working spatial memory. Eur. J. Neurosci. 2004, 19, 705–712. [Google Scholar] [CrossRef]
- Kheirbek, M.; Drew, L.J.; Burghardt, N.S.; Costantini, D.O.; Tannenholz, L.; Ahmari, S.E.; Zeng, H.; Fenton, A.; Hen, R. Differential Control of Learning and Anxiety along the Dorsoventral Axis of the Dentate Gyrus. Neuron 2013, 77, 955–968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanger, D.J.; Joly, D. Performance of a passive avoidance response is disrupted by compounds acting at 5HT(1A) receptors. Behav. Pharmacol. 1989, 1, 235–240. [Google Scholar]
- Carli, M.; Samanin, R. 8-Hydroxy-2-(di-n-propylamino)tetralin impairs spatial learning in a water maze: Role of postsynaptic 5-HT1A receptors. Br. J. Pharmacol. 1992, 105, 720–726. [Google Scholar] [CrossRef] [Green Version]
- Luttgen, M.; Elvander, E.; Madjid, N.; Ogren, S.O. Analysis of the role of 5-HT1A receptors in spatial and aversive learning in the rat. Neuropharmacology 2005, 48, 830–852. [Google Scholar] [CrossRef]
- Carli, M.; Lazarova, M.; Tatarczynska, E.; Samanin, R. Stimulation of 5-HT1A receptors in the dorsal hippocampus impairs acquisition and performance of a spatial task in a water maze. Brain Res. 1992, 595, 50–56. [Google Scholar] [CrossRef]
- Carli, M.; Silva, S.; Balducci, C.; Samanin, R. WAY 100635, a 5-HT1A receptor antagonist, prevents the impairment of spatial learning caused by blockade of hippocampal NMDA receptors. Neuropharmacology 1999, 38, 1165–1173. [Google Scholar] [CrossRef]
- Ögren, S.O.; Eriksson, T.M.; Elvander-Tottie, E.; D’Addario, C.; Ekström, J.C.; Svenningsson, P.; Meister, B.; Kehr, J.; Stiedl, O. The role of 5-HT1A receptors in learning and memory. Behav. Brain Res. 2008, 195, 54–77. [Google Scholar] [CrossRef]
- Bianchi, M.; Moser, C.; Lazzarini, C.; Vecchiato, E.; Crespi, F. Forced swimming test and fluoxetine treatment: In vivo evidence that peripheral 5-HT in rat platelet-rich plasma mirrors cerebral extracellular 5-HT levels, whilst 5-HT in isolated platelets mirrors neuronal 5-HT changes. Exp. Brain Res. 2002, 143, 191–197. [Google Scholar] [CrossRef]
- Caccia, S.; Anelli, M.; Codegoni, A.M.; Fracasso, C.; Garattini, S. The effects of single and repeated anorectic doses of 5-hydroxytryptamine uptake inhibitors on indole levels in rat brain. Br. J. Pharmacol. 1993, 110, 355–359. [Google Scholar] [CrossRef] [Green Version]
- Popa, D.; Cerdan, J.; Repérant, C.; Guiard, B.P.; Guilloux, J.-P.; David, D.; Gardier, A.M. A longitudinal study of 5-HT outflow during chronic fluoxetine treatment using a new technique of chronic microdialysis in a highly emotional mouse strain. Eur. J. Pharmacol. 2010, 628, 83–90. [Google Scholar] [CrossRef]
- Vaswani, M.; Linda, F.K.; Ramesh, S. Role of selective serotonin reuptake inhibitors in psychiatric disorders: A comprehensive review. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2003, 27, 85–102. [Google Scholar] [CrossRef]
- Blier, P.; Seletti, B.; Gilbert, F.; Young, S.N.; Benkelfat, C. Serotonin 1A Receptor Activation and Hypothermia in Humans Lack of Evidence for a Presynaptic Mediation. Neuropsychopharmacology 2002, 27, 301–308. [Google Scholar] [CrossRef]
- Wang, L.-E.; Zhang, X.-Q.; Yin, Y.-Q.; Zhang, Y.-H. Augmentative effect of spinosin on pentobarbital-induced loss of righting reflex in mice associated with presynaptic 5-HT1A receptor. J. Pharm. Pharmacol. 2012, 64, 277–282. [Google Scholar] [CrossRef] [PubMed]
- Lifschytz, T.; Zozulinsky, P.; Eitan, R.; Landshut, G.; Ohayon, S.; Lerer, B. Effect of Triiodothyronine on Antidepressant Screening Tests in Mice and on Presynaptic 5-HT1A Receptors: Mediation by Thyroid Hormone α Receptors. J. Pharmacol. Exp. Ther. 2011, 337, 494–502. [Google Scholar] [CrossRef]
- Overstreet, D.H.; Daws, L.C.; Schiller, G.D.; Orbach, J.; Janowsky, D.S. Cholinergic/serotonergic interactions in hypothermia: Implications for rat models of depression. Pharmacol. Biochem. Behav. 1998, 59, 777–785. [Google Scholar] [CrossRef]
- Overstreet, D.H.; Rezvani, A.H.; Knapp, D.J.; Crews, F.T.; Janowsky, D.S. Further selection of rat lines differing in 5-HT-1A receptor sensitivity: Behavioral and functional correlates. Psychiatr. Genet. 1996, 6, 107–117. [Google Scholar] [CrossRef]
- File, S.E.; Ouagazzal, A.-M.; Gonzalez, L.E.; Overstreet, D.H. Chronic Fluoxetine in Tests of Anxiety in Rat Lines Selectively Bred for Differential 5-HT1A Receptor Function. Pharmacol. Biochem. Behav. 1999, 62, 695–701. [Google Scholar] [CrossRef]
- Gonzalez, L.E.; File, S.E.; Overstreet, D.H. Selectively Bred Lines of Rats Differ in Social Interaction and Hippocampal 5-HT1A Receptor Function: A Link Between Anxiety and Depression? Pharmacol. Biochem. Behav. 1998, 59, 787–792. [Google Scholar] [CrossRef]
- Hensler, J.G.; Kovachich, G.B.; Frazer, A. A quantitative autoradiographic study of serotonin1A receptor regulation. Effect of 5,7-dihydroxytryptamine and antidepressant treatments. Neuropsychopharmacology 1991, 4, 131–144. [Google Scholar]
- Kelly, J.; Leonard, B. The effect of tianeptine and sertraline in three animal models of depression. Neuropharmacology 1994, 33, 1011–1016. [Google Scholar] [CrossRef]
- Riad, M.; Rbah, L.; Verdurand, M.; Aznavour, N.; Zimmer, L.; Descarries, L. Unchanged density of 5-HT1A autoreceptors on the plasma membrane of nucleus raphe dorsalis neurons in rats chronically treated with fluoxetine. Neuroscience 2008, 151, 692–700. [Google Scholar] [CrossRef]
- Gorinski, N.; Bijata, M.; Prasad, S.; Wirth, A.; Galil, D.A.; Zeug, A.; Bazovkina, D.; Kondaurova, E.; Kulikova, E.; Ilchibaeva, T.; et al. Attenuated palmitoylation of serotonin receptor 5-HT1A affects receptor function and contributes to depression-like behaviors. Nat. Commun. 2019, 10, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naumenko, V.S.; Bazovkina, D.V.; Kondaurova, E.M. On the Functional Cross-Talk between Brain 5-HT1A and 5-HT2A Receptors. Zhurnal Vyss. Nervn. deiatelnosti Im. IP Pavlov. 2015, 65, 240–247. [Google Scholar]
- Naumenko, V.S.; Bazovkina, D.V.; Kondaurova, E.M.; Zubkov, E.A.; Kulikov, A.V. The role of 5-HT2A receptor and 5-HT2A/5-HT1A receptor interaction in the suppression of catalepsy. Genes Brain Behav. 2010, 9, 519–524. [Google Scholar]
- Naumenko, V.S.; Popova, N.K.; Lacivita, E.; Leopoldo, M.; Ponimaskin, E.G. Interplay between Serotonin 5-HT1Aand 5-HT7Receptors in Depressive Disorders. CNS Neurosci. Ther. 2014, 20, 582–590. [Google Scholar] [CrossRef]
- Liang, T.; Chen, Q.; Li, Q.; Li, R.; Tang, J.; Hu, R.; Zhong, J.; Ge, H.; Liu, X.; Feng, H. 5-HT1a activation in PO/AH area induces therapeutic hypothermia in a rat model of intracerebral hemorrhage. Oncotarget 2017, 8, 73613–73626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nibuya, M.; Morinobu, S.; Duman, R.S. Regulation of BDNF and trkB mRNA in rat brain by chronic electroconvulsive seizure and antidepressant drug treatments. J. Neurosci. 1995, 15, 7539–7547. [Google Scholar] [CrossRef]
- Itoh, T.; Tokumura, M.; Abe, K. Effects of rolipram, a phosphodiesterase 4 inhibitor, in combination with imipramine on depressive behavior, CRE-binding activity and BDNF level in learned helplessness rats. Eur. J. Pharmacol. 2004, 498, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Rogóz, Z.; Legutko, B. Combined treatment with imipramine and metyrapone induces hippocampal and cortical brain-derived neurotrophic factor gene expression in rats. Pharmacol. Rep. 2005, 57, 840–844. [Google Scholar] [PubMed]
- Brunoni, A.R.; Lopes, M.; Fregni, F. A systematic review and meta-analysis of clinical studies on major depression and BDNF levels: Implications for the role of neuroplasticity in depression. Int. J. Neuropsychopharmacol. 2008, 11, 1169–1180. [Google Scholar] [CrossRef]
- Hellweg, R.; Ziegenhorn, A.; Heuser, I.; Deuschle, M. Serum Concentrations of Nerve Growth Factor and Brain-Derived Neurotrophic Factor in Depressed Patients before and after Antidepressant Treatment. Pharmacopsychiatry 2008, 41, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-Y.; Kim, Y.-K. Plasma Brain-Derived Neurotrophic Factor as a Peripheral Marker for the Action Mechanism of Antidepressants. Neuropsychobiology 2008, 57, 194–199. [Google Scholar] [CrossRef] [PubMed]
- Sen, S.; Duman, R.; Sanacora, G. Serum Brain-Derived Neurotrophic Factor, Depression, and Antidepressant Medications: Meta-Analyses and Implications. Biol. Psychiatry 2008, 64, 527–532. [Google Scholar] [CrossRef] [Green Version]
- Changelian, P.S.; Feng, P.; King, T.C.; Milbrandt, J. Structure of the NGFI-A gene and detection of upstream sequences responsible for its transcriptional induction by nerve growth factor. Proc. Natl. Acad. Sci. USA 1989, 86, 377–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinnock, S.B.; Blake, A.M.; Platt, N.J.; Herbert, J. The Roles of BDNF, pCREB and Wnt3a in the Latent Period Preceding Activation of Progenitor Cell Mitosis in The Adult Dentate Gyrus by Fluoxetine. PLoS ONE 2010, 5, e13652. [Google Scholar] [CrossRef] [Green Version]
- Bourtchuladze, R.; Frenguelli, B.; Blendy, J.; Cioffi, D.; Schutz, G.; Silva, A.J. Deficient long-term memory in mice with a targeted mutation of the cAMP-responsive element-binding protein. Cell 1994, 79, 59–68. [Google Scholar] [CrossRef]
- Okuno, H. Regulation and function of immediate-early genes in the brain: Beyond neuronal activity markers. Neurosci. Res. 2011, 69, 175–186. [Google Scholar] [CrossRef]
- Rioja, J.; Santin, L.J.; Doña, A.; De Pablos, L.; Minano, F.J.; González-Barón, S.; Aguirre, J.A.; Sánchez, J.M. 5-HT1A receptor activation counteracts c-Fos immunoreactivity induced in serotonin neurons of the raphe nuclei after immobilization stress in the male rat. Neurosci. Lett. 2006, 397, 190–195. [Google Scholar] [CrossRef]
- Nakamura, A.; Naito, M.; Tsuruo, T.; Fujita, N. Freud-1/Aki1, a novel PDK1-interacting protein, functions as a scaffold to activate the PDK1/Akt pathway in epidermal growth factor signaling. Mol. Cell. Biol. 2008, 28, 5996–6009. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.-H.; Lai, L.-C.; Cheng, H.-C.; Chen, K.-R.; Syue, Y.-Z.; Lu, H.-C.; Lin, W.-Y.; Chen, S.-H.; Huang, H.-S.; Shiau, A.-L.; et al. TBK1-associated Protein in Endolysosomes (TAPE) Is an Innate Immune Regulator Modulating the TLR3 and TLR4 Signaling Pathways. J. Biol. Chem. 2011, 286, 7043–7051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, M.; Li, X.D.; Chen, Z. CC2D1A, a DM14 and C2 domain protein, activates NF-kappaB through the canonical pathway. J. Biol. Chem. 2010, 285, 24372–24380. [Google Scholar] [CrossRef] [Green Version]
- Hsiung, S.C.; Tamir, H.; Franke, T.F.; Liu, K.P. Roles of extracellular signal-regulated kinase and Akt signaling in coordinating nuclear transcription factor-kappaB-dependent cell survival after serotonin 1A receptor activation. J. Neurochem. 2005, 95, 1653–1666. [Google Scholar] [CrossRef]
- Manzini, M.C.; Xiong, L.; Shaheen, R.; Tambunan, D.E.; Di Costanzo, S.; Mitisalis, V.; Tischfield, D.J.; Cinquino, A.; Ghaziuddin, M.; Christian, M.; et al. CC2D1A regulates human intellectual and social function as well as NF-kappaB signaling homeostasis. Cell Rep. 2014, 8, 647–655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popova, N.K.; Naumenko, E.V. Dominance relations and the pituitary-adrenal system in rats. Anim. Behav. 1972, 20, 108–111. [Google Scholar] [CrossRef]
- Serova, L.I.; Naumenko, E.V. The brain catecholamine systems in the regulation of dominance. Zhurnal Vyss. Nervn. Deiatelnosti Im. IP Pavlov. 1990, 40, 490–496. [Google Scholar]
- Wirth, A.; Chen-Wacker, C.; Wu, Y.-W.; Gorinski, N.; Filippov, M.A.; Pandey, G.; Ponimaskin, E. Dual lipidation of the brain-specific Cdc42 isoform regulates its functional properties. Biochem. J. 2013, 456, 311–322. [Google Scholar] [CrossRef] [Green Version]
- Grimm, D.; Kay, M.A.; Kleinschmidt, J.A. Helper virus-free, optically controllable, and two-plasmid-based production of adeno-associated virus vectors of serotypes 1 to 6. Mol. Ther. 2003, 7, 839–850. [Google Scholar] [CrossRef]
- Kobe, F.; Guseva, P.D.; Jensen, T.; Wirth, A.; Renner, U.; Hess, D.; Müller, M.; Medrihan, L.; Zhang, W.; Zhang, M.; et al. 5-HT7R/G12 Signaling Regulates Neuronal Morphology and Function in an Age-Dependent Manner. J. Neurosci. 2012, 32, 2915–2930. [Google Scholar] [CrossRef]
- Slotnick, B.M.; Leonard, C.M. A Stereotaxic Atlas of the Albino Mouse Forebrain; U.S. Department of Health, Education and Welfare: Rockville, MD, USA, 1975; p. 174. [Google Scholar]
- Kulikov, A.V.; Tikhonova, M.A.; Kulikov, V.A. Automated measurement of spatial preference in the open field test with transmitted lighting. J. Neurosci. Methods 2008, 170, 345–351. [Google Scholar] [CrossRef]
- Kulikov, A.V.; Morozova, M.V.; Kulikov, V.A.; Kirichuk, V.S.; Popova, N.K. Automated analysis of antidepressants’ effect in the forced swim test. J. Neurosci. Methods 2010, 191, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Naumenko, V.S.; Kondaurova, E.; Popova, N.K. On the role of brain 5-HT7 receptor in the mechanism of hypothermia: Comparison with hypothermia mediated via 5-HT1A and 5-HT3 receptor. Neuropharmacology 2011, 61, 1360–1365. [Google Scholar] [CrossRef]
- Kulikov, A.V.; Fursenko, D.V.; Khotskin, N.V.; Bazovkina, D.V.; Kulikov, V.A.; Naumenko, V.S.; Bazhenova, E.Y.; Popova, N.K. Spatial learning in the Morris water maze in mice genetically different in the predisposition to catalepsy: The effect of intraventricular treatment with brain-derived neurotrophic factor. Pharmacol. Biochem. Behav. 2014, 122, 266–272. [Google Scholar] [CrossRef]
- Dalm, S.; Grootendorst, J.; De Kloet, E.R.; Oitzl, M.S. Quantification of swim patterns in the morris water maze. Behav. Res. Methods Instrum. Comput. 2000, 32, 134–139. [Google Scholar] [CrossRef] [PubMed]
- Kulikov, A.V.; Naumenko, V.S.; Voronova, I.P.; Tikhonova, M.A.; Popova, N.K. Quantitative RT-PCR assay of 5-HT1A and 5-HT2A serotonin receptor mRNAs using genomic DNA as an external standard. J. Neurosci. Methods 2005, 141, 97–101. [Google Scholar] [CrossRef]
- Naumenko, V.S.; Kulikov, A.V. Quantitative assay of 5-HT(1A) serotonin receptor gene expression in the brain. Mol. Biol. 2006, 40, 37–44. [Google Scholar] [CrossRef]
- Naumenko, V.S.; Osipova, D.V.; Kostina, E.V.; Kulikov, A.V. Utilization of a two-standard system in real-time PCR for quantification of gene expression in the brain. J. Neurosci. Methods 2008, 170, 197–203. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence | Annealing Temperature, °C | Product Length, bp |
|---|---|---|---|
| Htr1a | F 5′-ctgtgacctgtttatcgccctg-3′ R 5′-gtagtctatagggtcggtgattgc-3′ | 62 | 200 |
| Cc2d1a | F 5′-gcaaagccgggcaacatcatc-3′ R 5′-tagcagaggtgggtgtagtgg-3′ | 60 | 181 |
| Bdnf | F 5′-tagcaaaaagagaattggctg-3′ R 5′-tttcaggtcatggatatgtcc-3′ | 59 | 255 |
| Ntrk2 | F 5′-cattcactgtgagaggcaacc-3′ F 5′-atcagggtgtagtctccgttatt-3′ | 63 | 175 |
| Ngfr | F 5′-acaacacccagcacccagga-3′ R 5′-cacaaccacagcagccaaga-3′ | 62 | 171 |
| Rela | F 5′-gcacctgttccaaagagcac-3′ R 5′-gagttgtccacagatgccagg-3′ | 64 | 93 |
| Nfkb1 | F 5′-cgtctgtctgctctctctcgac-3′ R 5′-ctcgcctcggtacttctctctc-3′ | 64 | 152 |
| Creb | F 5′-gctggctaacaatggtacggat-3′ R 5′-tggttgctgggcactagaat-3′ | 64 | 140 |
| cFos | F 5′-aaagagaaggaaaaactggag-3′ R 5′-cggaaacaagaagtcatcaa-3′ | 58 | 264 |
| Tph2 | F 5′-cattcctcgcacaattccagtcg-3′ R 5′-cttgacatattcaactagacgctc-3′ | 61 | 239 |
| Maoa | F 5′-atgaggatgttaaatgggtagatgttggt-3′ R 5′-cttgacatattcaactagacgctc- | 61 | 138 |
| Polr2a | F 5′-tgtgacaactccatacaatgc-3′ R 5′-ctctcttagtgaatttgcgtact-3′ | 60 | 194 |
| Antibodies, Manufacturer, Cat. No. | Dilution | Incubation Time, Conditions |
|---|---|---|
| Rabbit polyclonal antibody to 5-HT1A protein, Abcam, Cambridge, UK, ab85615 | 1:1000 in TBST supplemented with 5% milk powder | Night at 4 °C, as a primary antibody |
| Rabbit monoclonal antibody to CC2D1A/Freud-1 protein, Abcam, Cambridge, UK, ab191472 | 1:2000 in TBST supplemented with 5% milk powder | Night at 4 °C, as a primary antibody |
| Rabbit antibody to BDNF protein, Abcam, Cambridge, UK, ab46176 | 1:1000 in TBST supplemented with 5% milk powder | Night at 4 °C, as a primary antibody |
| Mouse antibody to pro-BDNF protein, Santa Cruz Biotechnology, Dallas, TX, USA, sc65513 | 1:250 in TBST supplemented with 5% milk powder | Night at 4 °C, as a primary antibody |
| Rabbit antibody to p75NTR, Abcam, Cambridge, UK, ab38335 | 1:500 in TBST supplemented with 5% milk powder | 2 h at RT, as a primary antibody |
| Rabbit antibody to TrkB, Abcam, Cambridge, UK, ab18987 | 1:400 in TBST supplemented with 3% BSA | Night at 4 °C, as a primary antibody |
| Rabbit monoclonal [E113] antibody to CREB protein (phospho S133), Abcam, Cambridge, UK, ab32096 | 1:1000 in TBST supplemented with 5% BSA | Night at 4 °C, as a primary antibody |
| Rabbit polyclonal antibody to CREB protein, Abcam, Cambridge, UK, ab31387 | 1:1000 in TBST supplemented with 5% BSA | Night at 4 °C, as a primary antibody |
| Rabbit polyclonal antibody to GAPDH protein, Abcam, Cambridge, UK, ab9485 | 1:2500 in TBST supplemented with 5% BSA | Night at 4 °C, as a primary antibody |
| Rabbit polyclonal antibody to MAOA Abcam, Cambridge, UK, ab126751 | 1:500 in TBST supplemented with 5% milk powder | Night at 4 °C, as a primary antibody |
| Rabbit polyclonal antibody to TPH2 Abcam, Cambridge, UK, ab184505 | 1:1000 in TBST supplemented with 5% milk powder | Night at 4 °C, as a primary antibody |
| Goat anti-rabbit IgG antibody conjugated to horseradish peroxidase, Invitrogen, Waltham, MA, USA, G-21234 | 1:10000 in TBST supplemented with 5% milk powder | 1 h at RT, as a secondary antibody |
| Goat anti-mouse IgG antibody conjugated to horseradish peroxidase, Abcam, Cambridge, UK, ab6728 | 1:2000 in TBST supplemented with 5% milk powder | 1 h at RT, as a secondary antibody |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kondaurova, E.M.; Plyusnina, A.V.; Ilchibaeva, T.V.; Eremin, D.V.; Rodnyy, A.Y.; Grygoreva, Y.D.; Naumenko, V.S. Effects of a Cc2d1a/Freud-1 Knockdown in the Hippocampus on Behavior, the Serotonin System, and BDNF. Int. J. Mol. Sci. 2021, 22, 13319. https://doi.org/10.3390/ijms222413319
Kondaurova EM, Plyusnina AV, Ilchibaeva TV, Eremin DV, Rodnyy AY, Grygoreva YD, Naumenko VS. Effects of a Cc2d1a/Freud-1 Knockdown in the Hippocampus on Behavior, the Serotonin System, and BDNF. International Journal of Molecular Sciences. 2021; 22(24):13319. https://doi.org/10.3390/ijms222413319
Chicago/Turabian StyleKondaurova, Elena M., Alexandra V. Plyusnina, Tatiana V. Ilchibaeva, Dmitry V. Eremin, Alexander Ya. Rodnyy, Yulia D. Grygoreva, and Vladimir S. Naumenko. 2021. "Effects of a Cc2d1a/Freud-1 Knockdown in the Hippocampus on Behavior, the Serotonin System, and BDNF" International Journal of Molecular Sciences 22, no. 24: 13319. https://doi.org/10.3390/ijms222413319