
Role of Oxytocin and Vasopressin in Neuropsychiatric Disorders: Therapeutic Potential of Agonists and Antagonists

 ,
,  , and
, and
Abstract
:1. Introduction
2. Receptors: Brain Distribution, Expression Regulation, and Signaling
3. Neuropsychiatric Conditions: OT and AVP Role
3.1. Depression, Anxiety, and Stress
3.2. Substance Abuse
3.3. Social Behaviors
3.3.1. Social Interactions
3.3.2. Aggression
3.3.3. Maternal Behavior
3.3.4. Social Play Behavior
3.3.5. Social Behavior Dysfunction and Neuropsychiatric Conditions
4. Preclinical and Clinical Studies and Human Research with OT and AVP: The Good, the Bad, and the Ugly
5. Agonist and Antagonist of OT-R and AVP-R: Peptides and Non-Peptides Alternatives
5.1. OT-R
5.2. V1AR and V1BR
6. Concluding Remarks and Future Projections
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AC | adenylate cyclase |
| ACC | anterior cingulate cortex |
| AMPH | amphetamine |
| ADHD | attentional deficit hyperactive disorder |
| ASD | autism spectrum disorder |
| AVP | arginine vasopressin |
| AVP-R | arginine vasopressin receptor |
| BLA | basolateral amygdala |
| BNST | bed nucleus of stria terminalis |
| CPP | conditioned place preference |
| CeA | central amygdala |
| CNS | central nervous system |
| CSF | cerebrospinal fluid |
| DA | dopamine |
| DAG | 1,2 diacylglycerol |
| ERα | estrogen receptor α |
| ERβ | estrogen receptor β |
| GABA | gamma-aminobutyric acid |
| GAD | generalized anxiety disorder |
| GPCR | protein coupled receptor |
| HPA | hypothalamic-pituitary-adrenal |
| icv | intracerebroventricular |
| ip | intraperitoneal |
| IP3 | inositol triphosphate |
| IRK | inward rectifying K+ |
| KO | knockout |
| LS | lateral septum |
| MDD | major depressive disorder |
| MeA | medial amygdala |
| α-MSH | α-melanocyte stimulating hormone |
| NPs | neuropeptides |
| NAc | nucleus accumbens |
| NAcc | nucleus accumbens core |
| NAcSh | nucleus accumbens shell |
| PKC | protein kinase C |
| PLC | phospholipase C |
| po | pers or |
| PVN | parvocellular nucleus |
| PFC | prefrontal cortex |
| mPOA | medial preoptic area |
| OT | oxytocin |
| OT-R | oxytocin receptor |
| sc | subcutaneous injection |
| SN | substantia nigra |
| SON | supraoptic nucleus |
| VTA | ventral tegmental area |
| V1AR | vasopressin receptor type 1A |
| V1BR | vasopressin receptor type 1B |
| V2R | vasopressin receptor type 2 |
References
- Blackburn, T.P. Depressive Disorders: Treatment Failures and Poor Prognosis over the Last 50 Years. Pharmacol. Res. Perspect. 2019, 7, e00472. [Google Scholar] [CrossRef]
- Cuijpers, P. Measuring Success in the Treatment of Depression: What Is Most Important to Patients? Expert Rev. Neurother. 2020, 20, 123–125. [Google Scholar] [CrossRef]
- Caldwell, H.K.; Albers, H.E. Oxytocin, Vasopressin, and the Motivational Forces That Drive Social Behaviors. Curr. Top. Behav. Neurosci. 2016, 27, 51–103. [Google Scholar]
- Caldwell, H.K. Oxytocin and Vasopressin: Powerful Regulators of Social Behavior. Neuroscientist 2017, 23, 517–528. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, W.E.; Warach, S.; Hatton, G.I.; McNeill, T.H. Subnuclei in the Rat Hypothalamic Paraventricular Nucleus: A Cytoarchitectural, Horseradish Peroxidase and Immunocytochemical Analysis. Neuroscience 1980, 5, 1931–1958. [Google Scholar] [CrossRef]
- Armstrong, W.E. Hypothalamic Supraoptic and Paraventricular Nuclei. In The Rat Nervous System; Elsevier: Amsterdam, The Netherlands, 2015; pp. 295–314. ISBN 9780123742452. [Google Scholar]
- Grinevich, V.; Knobloch-Bollmann, H.S.; Eliava, M.; Busnelli, M.; Chini, B. Assembling the Puzzle: Pathways of Oxytocin Signaling in the Brain. Biol. Psychiatry 2016, 79, 155–164. [Google Scholar] [CrossRef] [Green Version]
- Meyer-Lindenberg, A.; Domes, G.; Kirsch, P.; Heinrichs, M. Oxytocin and Vasopressin in the Human Brain: Social Neuropeptides for Translational Medicine. Nat. Rev. Neurosci. 2011, 12, 524–538. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, W.E.; Stern, J.E. Electrophysiological Distinctions Between Oxytocin and Vasopressin Neurons in the Supraoptic Nucleus. Adv. Exp. Med. Biol. 1998, 449, 67–77. [Google Scholar] [PubMed]
- Stern, J.E.; Armstrong, W.E. Electrophysiological Differences between Oxytocin and Vasopressin Neurones Recorded from Female Rats in Vitro. J. Physiol. 1995, 488, 701–708. [Google Scholar] [CrossRef] [Green Version]
- Rood, B.D.; De Vries, G.J. Vasopressin Innervation of the Mouse (Mus Musculus) Brain and Spinal Cord. J. Comp. Neurol. 2011, 519, 2434–2474. [Google Scholar] [CrossRef] [Green Version]
- Busnelli, M.; Saulière, A.; Manning, M.; Bouvier, M.; Galés, C.; Chini, B. Functional Selective Oxytocin-Derived Agonists Discriminate between Individual G Protein Family Subtypes. J. Biol. Chem. 2012, 287, 3617–3629. [Google Scholar] [CrossRef] [Green Version]
- Matsuzaki, M.; Matsushita, H.; Tomizawa, K.; Matsui, H. Oxytocin: A Therapeutic Target for Mental Disorders. J. Physiol. Sci. 2012, 62, 441–444. [Google Scholar] [CrossRef]
- Acevedo-Rodriguez, A.; Mani, S.K.; Handa, R.J. Oxytocin and Estrogen Receptor β in the Brain: An Overview. Front. Endocrinol. 2015, 6, 160. [Google Scholar] [CrossRef] [Green Version]
- Knepper, M.A.; Kwon, T.-H.; Nielsen, S. Molecular Physiology of Water Balance. N. Engl. J. Med. 2015, 372, 1349–1358. [Google Scholar] [CrossRef]
- Baribeau, D.A.; Anagnostou, E. Oxytocin and Vasopressin: Linking Pituitary Neuropeptides and Their Receptors to Social Neurocircuits. Front. Neurosci. 2015, 9, 335. [Google Scholar] [CrossRef] [Green Version]
- Koshimizu, T.-A.; Nakamura, K.; Egashira, N.; Hiroyama, M.; Nonoguchi, H.; Tanoue, A. Vasopressin V1a and V1b Receptors: From Molecules to Physiological Systems. Physiol. Rev. 2012, 92, 1813–1864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacDonald, K.; Feifel, D. Oxytocin’s Role in Anxiety: A Critical Appraisal. Brain Res. 2014, 1580, 22–56. [Google Scholar] [CrossRef] [PubMed]
- Neumann, I.D.; Landgraf, R. Balance of Brain Oxytocin and Vasopressin: Implications for Anxiety, Depression, and Social Behaviors. Trends Neurosci. 2012, 35, 649–659. [Google Scholar] [CrossRef]
- Kirsch, P. Oxytocin in the Socioemotional Brain: Implications for Psychiatric Disorders. Dialogues Clin. Neurosci. 2015, 17, 463–476. [Google Scholar]
- Sasaki, T.; Hashimoto, K.; Oda, Y.; Ishima, T.; Kurata, T.; Takahashi, J.; Kamata, Y.; Kimura, H.; Niitsu, T.; Komatsu, H.; et al. Decreased Levels of Serum Oxytocin in Pediatric Patients with Attention Deficit/Hyperactivity Disorder. Psychiatry Res. 2015, 228, 746–751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalyoncu, T.; Özbaran, B.; Köse, S.; Onay, H. Variation in the Oxytocin Receptor Gene Is Associated with Social Cognition and ADHD. J. Atten. Disord. 2019, 23, 702–711. [Google Scholar] [CrossRef] [PubMed]
- Taurines, R.; Schwenck, C.; Lyttwin, B.; Schecklmann, M.; Jans, T.; Reefschläger, L.; Geissler, J.; Gerlach, M.; Romanos, M. Oxytocin Plasma Concentrations in Children and Adolescents with Autism Spectrum Disorder: Correlation with Autistic Symptomatology. Atten. Defic. Hyperact. Disord. 2014, 6, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Oztan, O.; Garner, J.P.; Constantino, J.N.; Parker, K.J. Neonatal CSF Vasopressin Concentration Predicts Later Medical Record Diagnoses of Autism Spectrum Disorder. Proc. Natl. Acad. Sci. USA 2020, 117, 10609–10613. [Google Scholar] [CrossRef] [PubMed]
- Lukas, M.; Bredewold, R.; Landgraf, R.; Neumann, I.D.; Veenema, A.H. Early Life Stress Impairs Social Recognition due to a Blunted Response of Vasopressin Release within the Septum of Adult Male Rats. Psychoneuroendocrinology 2011, 36, 843–853. [Google Scholar] [CrossRef]
- Choleris, E.; Pfaff, D.; Kavaliers, M. Oxytocin, vasopressin, and related peptides in the regulation of behavior: Where next? In Oxytocin, Vasopressin and Related Peptides in the Regulation of Behavior; Cambridge University Press: Cambridge, UK, 2013; pp. 379–381. [Google Scholar]
- Shalev, I.; Ebstein, R.P. Social Hormones and Human Behavior: What Do We Know and Where Do We Go from Here; Frontiers Media SA: Lausanne, Switzerland, 2015; ISBN 9782889194070. [Google Scholar]
- Ring, R.H.; Schechter, L.E.; Leonard, S.K.; Dwyer, J.M.; Platt, B.J.; Graf, R.; Grauer, S.; Pulicicchio, C.; Resnick, L.; Rahman, Z.; et al. Receptor and Behavioral Pharmacology of WAY-267464, a Non-Peptide Oxytocin Receptor Agonist. Neuropharmacology 2010, 58, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Albers, H.E. Species, Sex and Individual Differences in the Vasotocin/vasopressin System: Relationship to Neurochemical Signaling in the Social Behavior Neural Network. Front. Neuroendocrinol. 2015, 36, 49–71. [Google Scholar] [CrossRef] [Green Version]
- Song, Z.; Elliott Albers, H. Cross-Talk among Oxytocin and Arginine-Vasopressin Receptors: Relevance for Basic and Clinical Studies of the Brain and Periphery. Front. Neuroendocrinol. 2018, 51, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Ebstein, R.P.; Shalev, I.; Israel, S.; Uzefovsky, F.; Avinun, R.; Knafo, A.; Yirmiya, N.; Mankuta, D. Oxytocin and Vasopressin in Human Sociality and Social Psychopathologies. In Oxytocin, Vasopressin and Related Peptides in the Regulation of Behavior; Cambridge University Press: Cambridge, UK, 2013; pp. 343–366. [Google Scholar]
- Wilkie, T.M.; Scherle, P.A.; Strathmann, M.P.; Slepak, V.Z.; Simon, M.I. Characterization of G-Protein Alpha Subunits in the Gq Class: Expression in Murine Tissues and in Stromal and Hematopoietic Cell Lines. Proc. Natl. Acad. Sci. USA 1991, 88, 10049–10053. [Google Scholar] [CrossRef] [Green Version]
- Boccia, M.L.; Petrusz, P.; Suzuki, K.; Marson, L.; Pedersen, C.A. Immunohistochemical Localization of Oxytocin Receptors in Human Brain. Neuroscience 2013, 253, 155–164. [Google Scholar] [CrossRef]
- Rault, J.-L.; Carter, C.S.; Garner, J.P.; Marchant-Forde, J.N.; Richert, B.T.; Lay, D.C., Jr. Repeated Intranasal Oxytocin Administration in Early Life Dysregulates the HPA Axis and Alters Social Behavior. Physiol. Behav. 2013, 112–113, 40–48. [Google Scholar] [CrossRef]
- Skuse, D.H.; Gallagher, L. Dopaminergic-Neuropeptide Interactions in the Social Brain. Trends in Cognitive Sciences 2009, 13, 27–35. [Google Scholar] [CrossRef]
- Garibay, J.L.; Kozasa, T.; Itoh, H.; Tsukamoto, T.; Matsuoka, M.; Kaziro, Y. Analysis by mRNA Levels of the Expression of Six G Protein Alpha-Subunit Genes in Mammalian Cells and Tissues. Biochim. Biophys. Acta 1991, 1094, 193–199. [Google Scholar] [CrossRef]
- Berrada, K.; Plesnicher, C.L.; Luo, X.; Thibonnier, M. Dynamic Interaction of Human Vasopressin/oxytocin Receptor Subtypes with G Protein-Coupled Receptor Kinases and Protein Kinase C after Agonist Stimulation. J. Biol. Chem. 2000, 275, 27229–27237. [Google Scholar] [CrossRef]
- Gravati, M.; Busnelli, M.; Bulgheroni, E.; Reversi, A.; Spaiardi, P.; Parenti, M.; Toselli, M.; Chini, B. Dual Modulation of Inward Rectifier Potassium Currents in Olfactory Neuronal Cells by Promiscuous G Protein Coupling of the Oxytocin Receptor. J. Neurochem. 2010, 114, 1424–1435. [Google Scholar] [CrossRef]
- Thibonnier, M.; Schork, N.J. The Genetics of Hypertension. Curr. Opin. Genet. Dev. 1995, 5, 362–370. [Google Scholar] [CrossRef]
- Wettschureck, N.; Moers, A.; Hamalainen, T.; Lemberger, T.; Schütz, G.; Offermanns, S. Heterotrimeric G Proteins of the Gq/11 Family Are Crucial for the Induction of Maternal Behavior in Mice. Mol. Cell. Biol. 2004, 24, 8048–8054. [Google Scholar] [CrossRef] [Green Version]
- De Dreu, C.K.W. Oxytocin Modulates Cooperation within and Competition between Groups: An Integrative Review and Research Agenda. Horm. Behav. 2012, 61, 419–428. [Google Scholar] [CrossRef]
- Song, Z.; Borland, J.M.; Larkin, T.E.; O’Malley, M.; Albers, H.E. Activation of Oxytocin Receptors, but Not Arginine-Vasopressin V1a Receptors, in the Ventral Tegmental Area of Male Syrian Hamsters Is Essential for the Reward-like Properties of Social Interactions. Psychoneuroendocrinology 2016, 74, 164–172. [Google Scholar] [CrossRef]
- Voglis, G.; Tavernarakis, N. The Role of Synaptic Ion Channels in Synaptic Plasticity. EMBO Rep. 2006, 7, 1104–1110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waltenspühl, Y.; Schöppe, J.; Ehrenmann, J.; Kummer, L.; Plückthun, A. Crystal Structure of the Human Oxytocin Receptor. Sci. Adv. 2020, 6, eabb5419. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Seuthe, A.B.; Ehrler, O.T.; Zhang, X.; Wyttenbach, T.; Hsu, J.F.; Bowers, M.T. Oxytocin-Receptor Binding: Why Divalent Metals Are Essential. J. Am. Chem. Soc. 2005, 127, 2024–2025. [Google Scholar] [CrossRef]
- Alshanski, I.; Shalev, D.E.; Yitzchaik, S.; Hurevich, M. Determining the Structure and Binding Mechanism of Oxytocin-Cu Complex Using Paramagnetic Relaxation Enhancement NMR Analysis. J. Biol. Inorg. Chem. 2021, 26, 809–815. [Google Scholar] [CrossRef]
- Birnbaumer, M. Vasopressin Receptors. Trends Endocrinol Metab. 2000, 11, 406–410. [Google Scholar] [CrossRef]
- Koshimizu, T.-A.; Nasa, Y.; Tanoue, A.; Oikawa, R.; Kawahara, Y.; Kiyono, Y.; Adachi, T.; Tanaka, T.; Kuwaki, T.; Mori, T.; et al. V1a Vasopressin Receptors Maintain Normal Blood Pressure by Regulating Circulating Blood Volume and Baroreflex Sensitivity. Proc. Natl. Acad. Sci. USA 2006, 103, 7807–7812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antoni, F.A. Vasopressinergic Control of Pituitary Adrenocorticotropin Secretion Comes of Age. Front. Neuroendocrinol. 1993, 14, 76–122. [Google Scholar] [CrossRef]
- Aguilera, G.; Pham, Q.; Rabadan-Diehl, C. Regulation of Pituitary Vasopressin Receptors during Chronic Stress: Relationship to Corticotroph Responsiveness. J. Neuroendocrinol. 1994, 6, 299–304. [Google Scholar] [CrossRef]
- Litvin, Y.; Murakami, G.; Pfaff, D.W. Effects of Chronic Social Defeat on Behavioral and Neural Correlates of Sociality: Vasopressin, Oxytocin and the Vasopressinergic V1b Receptor. Physiol. Behav. 2011, 103, 393–403. [Google Scholar] [CrossRef] [PubMed]
- Overstreet, D.H.; Griebel, G. Antidepressant-like Effects of the Vasopressin V1b Receptor Antagonist SSR149415 in the Flinders Sensitive Line Rat. Pharmacol. Biochem. Behav. 2005, 82, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Roos, K.P.; Strait, K.A.; Raphael, K.L.; Blount, M.A.; Kohan, D.E. Collecting Duct-Specific Knockout of Adenylyl Cyclase Type VI Causes a Urinary Concentration Defect in Mice. Am. J. Physiol. Ren. Physiol. 2012, 302, F78–F84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vargas, K.J.; Sarmiento, J.M.; Ehrenfeld, P.; Añazco, C.C.; Villanueva, C.I.; Carmona, P.L.; Brenet, M.; Navarro, J.; Müller-Esterl, W.; González, C.B. Postnatal Expression of V2 Vasopressin Receptor Splice Variants in the Rat Cerebellum. Differentiation 2009, 77, 377–385. [Google Scholar] [CrossRef] [Green Version]
- Krieg, S.M.; Sonanini, S.; Plesnila, N.; Trabold, R. Effect of Small Molecule Vasopressin V1a and V2 Receptor Antagonists on Brain Edema Formation and Secondary Brain Damage Following Traumatic Brain Injury in Mice. J. Neurotrauma 2015, 32, 221–227. [Google Scholar] [CrossRef]
- Okhovat, M.; Maguire, S.M.; Phelps, S.M. Methylation of in the Cortex of Wild Prairie Voles: Effects of CpG Position and Polymorphism. R. Soc. Open Sci. 2017, 4, 160646. [Google Scholar] [CrossRef] [Green Version]
- Okhovat, M.; Chen, I.C.; Dehghani, Z.; Zheng, D.J.; Ikpatt, J.E.; Momoh, H.; Phelps, S.M. Genetic Variation in the Developmental Regulation of Cortical avpr1a among Prairie Voles. Genes Brain Behav. 2018, 17, 36–48. [Google Scholar] [CrossRef] [Green Version]
- Murgatroyd, C.; Patchev, A.V.; Wu, Y.; Micale, V.; Bockmühl, Y.; Fischer, D.; Holsboer, F.; Wotjak, C.T.; Almeida, O.F.X.; Spengler, D. Dynamic DNA Methylation Programs Persistent Adverse Effects of Early-Life Stress. Nat. Neurosci. 2009, 12, 1559–1566. [Google Scholar] [CrossRef] [PubMed]
- Roper, J.; O’Carroll, A.-M.; Young, W., 3rd; Lolait, S. The Vasopressin Avpr1b Receptor: Molecular and Pharmacological Studies. Stress 2011, 14, 98–115. [Google Scholar] [CrossRef]
- Anisman, H.; Hayley, S.; Kelly, O.; Borowski, T.; Merali, Z. Psychogenic, Neurogenic, and Systemic Stressor Effects on Plasma Corticosterone and Behavior: Mouse Strain-Dependent Outcomes. Behav. Neurosci. 2001, 115, 443–454. [Google Scholar] [CrossRef] [PubMed]
- Wersinger, S.R.; Caldwell, H.K.; Christiansen, M.; Young, W.S., 3rd. Disruption of the Vasopressin 1b Receptor Gene Impairs the Attack Component of Aggressive Behavior in Mice. Genes Brain Behav. 2007, 6, 653–660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wersinger, S.R.; Ginns, E.I.; O’Carroll, A.-M.; Lolait, S.J.; Young, W.S., III. Vasopressin V1b Receptor Knockout Reduces Aggressive Behavior in Male Mice. Mol. Psychiatry 2002, 7, 975–984. [Google Scholar] [CrossRef] [Green Version]
- Kalin, N.H. The Critical Relationship between Anxiety and Depression. Am. J. Psychiatry 2020, 177, 365–367. [Google Scholar] [CrossRef]
- Selye, H. Stress and Distress. Compr. Ther. 1975, 1, 9–13. [Google Scholar]
- Selye, H. Stress and the General Adaptation Syndrome. Br. Med. J. 1950, 1, 1383–1392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slattery, D.A.; Neumann, I.D. Chronic Icv Oxytocin Attenuates the Pathological High Anxiety State of Selectively Bred Wistar Rats. Neuropharmacology 2010, 58, 56–61. [Google Scholar] [CrossRef]
- Mak, P.; Broussard, C.; Vacy, K.; Broadbear, J.H. Modulation of Anxiety Behavior in the Elevated plus Maze Using Peptidic Oxytocin and Vasopressin Receptor Ligands in the Rat. J. Psychopharmacol. 2012, 26, 532–542. [Google Scholar] [CrossRef] [PubMed]
- Lancel, M.; Krömer, S.; Neumann, I.D. Intracerebral Oxytocin Modulates Sleep-Wake Behaviour in Male Rats. Regul. Pept. 2003, 114, 145–152. [Google Scholar] [CrossRef]
- Slattery, D.A.; Neumann, I.D. Oxytocin and Major Depressive Disorder: Experimental and Clinical Evidence for Links to Aetiology and Possible Treatment. Pharmaceuticals 2010, 3, 702–724. [Google Scholar] [CrossRef] [Green Version]
- Leuner, B.; Caponiti, J.M.; Gould, E. Oxytocin Stimulates Adult Neurogenesis Even under Conditions of Stress and Elevated Glucocorticoids. Hippocampus 2012, 22, 861–868. [Google Scholar] [CrossRef] [Green Version]
- Keck, M.E.; Welt, T.; Müller, M.B.; Uhr, M.; Ohl, F.; Wigger, A.; Toschi, N.; Holsboer, F.; Landgraf, R. Reduction of Hypothalamic Vasopressinergic Hyperdrive Contributes to Clinically Relevant Behavioral and Neuroendocrine Effects of Chronic Paroxetine Treatment in a Psychopathological Rat Model. Neuropsychopharmacology 2003, 28, 235–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bao, A.-M.; Swaab, D.F. Corticotropin-releasing hormone and arginine vasopressin in depression focus on the human postmortem Hypothalamus. Vitam. Horm. 2010, 82, 339–365. [Google Scholar]
- Koob, G.F. The Dark Side of Emotion: The Addiction Perspective. Eur. J. Pharmacol. 2015, 753, 73–87. [Google Scholar] [CrossRef] [Green Version]
- Koob, G.F. Addiction Is a Reward Deficit and Stress Surfeit Disorder. Front. Psychiatry 2013, 4, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blum, K.; Femino, J.; Teitelbaum, S.; Giordano, J.; Oscar-Berman, M.; Gold, M. Molecular Neurobiology of Addiction Recovery: The 12 Steps Program and Fellowship; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2013; ISBN 9781461472308. [Google Scholar]
- Koob, G.F. Circuits, Drugs, and Drug Addiction. Adv. Pharmacol. 1997, 42, 978–982. [Google Scholar]
- Koob, G.F.; Volkow, N.D. Neurocircuitry of Addiction. Neuropsychopharmacology 2010, 35, 217–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grant, J.E.; Brewer, J.A.; Potenza, M.N. The Neurobiology of Substance and Behavioral Addictions. CNS Spectr. 2006, 11, 924–930. [Google Scholar] [CrossRef] [PubMed]
- Meyers, R.A.; Zavala, A.R.; Neisewander, J.L. Dorsal, but Not Ventral, Hippocampal Lesions Disrupt Cocaine Place Conditioning. NeuroReport 2003, 14, 2127–2131. [Google Scholar] [CrossRef] [PubMed]
- Sarnyai, Z.; Kovács, G.L. Role of Oxytocin in the Neuroadaptation to Drugs of Abuse. Psychoneuroendocrinology 1994, 19, 85–117. [Google Scholar] [CrossRef]
- Zhou, Y.; Litvin, Y.; Piras, A.P.; Pfaff, D.W.; Kreek, M.J. Persistent Increase in Hypothalamic Arginine Vasopressin Gene Expression during Protracted Withdrawal from Chronic Escalating-Dose Cocaine in Rodents. Neuropsychopharmacology 2011, 36, 2062–2075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarnyai, Z. Neurobiology of Stress and Cocaine Addiction. Studies on Corticotropin-Releasing Factor in Rats, Monkeys, and Humans. Ann. N. Y. Acad. Sci. 1998, 851, 371–387. [Google Scholar] [CrossRef] [PubMed]
- Ahumada, C.; Bahamondes, C.; Cerda, C.A.; Silva, R.A.; Cruz, G.; Moya, P.R.; Sotomayor-Zárate, R.; Renard, G.M. Amphetamine Treatment Affects the Extra-Hypothalamic Vasopressinergic System in a Sex- and Nucleus-Dependent Manner. J. Neuroendocrinol. 2017, 29. [Google Scholar] [CrossRef] [PubMed]
- Gárate-Pérez, M.F.; Méndez, A.; Bahamondes, C.; Sanhueza, C.; Guzmán, F.; Reyes-Parada, M.; Sotomayor-Zárate, R.; Renard, G.M. Vasopressin in the Lateral Septum Decreases Conditioned Place Preference to Amphetamine and Nucleus Accumbens Dopamine Release. Addict. Biol. 2021, 26, e12851. [Google Scholar] [CrossRef] [PubMed]
- You, Z.D.; Li, J.H.; Song, C.Y.; Wang, C.H.; Lu, C.L. Chronic Morphine Treatment Inhibits Oxytocin Synthesis in Rats. Neuroreport 2000, 11, 3113–3116. [Google Scholar] [CrossRef]
- Zhou, Y.; Leri, F.; Cummins, E.; Hoeschele, M.; Kreek, M.J. Involvement of Arginine Vasopressin and V1b Receptor in Heroin Withdrawal and Heroin Seeking Precipitated by Stress and by Heroin. Neuropsychopharmacology 2008, 33, 226–236. [Google Scholar] [CrossRef] [Green Version]
- Mennella, J.A.; Pepino, M.Y.; Teff, K.L. Acute Alcohol Consumption Disrupts the Hormonal Milieu of Lactating Women. J. Clin. Endocrinol. Metab. 2005, 90, 1979–1985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchesi, C.; Chiodera, P.; Brusamonti, E.; Volpi, R.; Coiro, V. Abnormal Plasma Oxytocin and β-Endorphin Levels in Alcoholics after Short and Long Term Abstinence. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 1997, 21, 797–807. [Google Scholar] [CrossRef]
- Paula-Barbosa, M.M.; Silva, S.M.; Andrade, J.P.; Cadete-Leite, A.; Madeira, M.D. Nerve Growth Factor Restores mRNA Levels and the Expression of Neuropeptides in the Suprachiasmatic Nucleus of Rats Submitted to Chronic Ethanol Treatment and Withdrawal. J. Neurocytol. 2001, 30, 195–207. [Google Scholar] [CrossRef]
- Silva, S.M.; Paula-Barbosa, M.M.; Madeira, M.D. Prolonged Alcohol Intake Leads to Reversible Depression of Corticotropin-Releasing Hormone and Vasopressin Immunoreactivity and mRNA Levels in the Parvocellular Neurons of the Paraventricular Nucleus. Brain Res. 2002, 954, 82–93. [Google Scholar] [CrossRef]
- Bowen, M.T.; Neumann, I.D. Rebalancing the Addicted Brain: Oxytocin Interference with the Neural Substrates of Addiction. Trends Neurosci. 2017, 40, 691–708. [Google Scholar] [CrossRef]
- Sarnyai, Z.; Babarczy, E.; Kriván, M.; Szabó, G.; Kovács, G.L.; Barth, T.; Telegdy, G. Selective Attenuation of Cocaine-Induced Stereotyped Behaviour by Oxytocin: Putative Role of Basal Forebrain Target Sites. Neuropeptides 1991, 19, 51–56. [Google Scholar] [CrossRef]
- Qi, J.; Yang, J.-Y.; Song, M.; Li, Y.; Wang, F.; Wu, C.-F. Inhibition by Oxytocin of Methamphetamine-Induced Hyperactivity Related to Dopamine Turnover in the Mesolimbic Region in Mice. Naunyn. Schmiedebergs. Arch. Pharmacol. 2008, 376, 441–448. [Google Scholar] [CrossRef]
- Xiao, L.; Priest, M.F.; Nasenbeny, J.; Lu, T.; Kozorovitskiy, Y. Biased Oxytocinergic Modulation of Midbrain Dopamine Systems. Neuron 2017, 95, 368–384.e5. [Google Scholar] [CrossRef] [Green Version]
- Peris, J.; Steck, M.R.; Krause, E.G. Oxytocin Treatment for Alcoholism: Potential Neurocircuitry Targets. Neuropharmacology 2020, 171, 108091. [Google Scholar] [CrossRef]
- Sarnyai, Z.; Szabó, G.; Kovács, G.L.; Telegdy, G. Oxytocin Attenuates the Cocaine-Induced Exploratory Hyperactivity in Mice. Neuroreport 1990, 1, 200–202. [Google Scholar] [CrossRef] [PubMed]
- Carson, D.S.; Cornish, J.L.; Guastella, A.J.; Hunt, G.E.; McGregor, I.S. Oxytocin Decreases Methamphetamine Self-Administration, Methamphetamine Hyperactivity, and Relapse to Methamphetamine-Seeking Behaviour in Rats. Neuropharmacology 2010, 58, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Peters, S.T.; Bowen, M.T.; Bohrer, K.; McGregor, I.S.; Neumann, I.D. Oxytocin Inhibits Ethanol Consumption and Ethanol-Induced Dopamine Release in the Nucleus Accumbens. Addict. Biol. 2017, 22, 702–711. [Google Scholar] [CrossRef]
- Hung, C.R.; Tabakoff, B.; Melchior, C.L.; Hoffman, P.L. Intraventricular Arginine Vasopressin Maintains Ethanol Tolerance. Eur. J. Pharmacol. 1984, 106, 645–648. [Google Scholar] [CrossRef]
- Hoffman, P.L.; Tabakoff, B. Neurohypophyseal Peptides Maintain Tolerance to the Incoordinating Effects of Ethanol. Pharmacol. Biochem. Behav. 1984, 21, 539–543. [Google Scholar] [CrossRef]
- Post, R.M.; Contel, N.R.; Gold, P. Impaired Behavioral Sensitization to Cocaine in Vasopressin Deficient Rats. Life Sci. 1982, 31, 2745–2750. [Google Scholar] [CrossRef]
- Chen, P.; Hong, W. Neural Circuit Mechanisms of Social Behavior. Neuron 2018, 98, 16–30. [Google Scholar] [CrossRef] [Green Version]
- Pedersen, C.A.; Prange, A.J., Jr. Oxytocin and Mothering Behavior in the Rat. Pharmacol. Ther. 1985, 28, 287–302. [Google Scholar] [CrossRef]
- McCarthy, M.M.; Bare, J.E.; vom Saal, F.S. Infanticide and Parental Behavior in Wild Female House Mice: Effects of Ovariectomy, Adrenalectomy and Administration of Oxytocin and Prostaglandin F2 Alpha. Physiol. Behav. 1986, 36, 17–23. [Google Scholar] [CrossRef]
- Leshner, A.I.; Roche, K.E. Comparison of the Effects of ACTH and Lysine Vasopressin on Avoidance-of-Attack in Mice. Physiol. Behav. 1977, 18, 879–883. [Google Scholar] [CrossRef]
- Roche, K.E.; Leshner, A.I. ACTH and Vasopressin Treatments Immediately after a Defeat Increase Future Submissiveness in Male Mice. Science 1979, 204, 1343–1344. [Google Scholar] [CrossRef]
- Crawley, J.N.; Hays, S.E.; O’Donohue, T.L.; Paul, S.M.; Goodwin, F.K. Neuropeptide Modulation of Social and Exploratory Behaviors in Laboratory Rodents. Peptides 1981, 2 (Suppl. S1), 123–129. [Google Scholar] [CrossRef]
- Kohli, S.; King, M.V.; Williams, S.; Edwards, A.; Ballard, T.M.; Steward, L.J.; Alberati, D.; Fone, K.C.F. Oxytocin Attenuates Phencyclidine Hyperactivity and Increases Social Interaction and Nucleus Accumben Dopamine Release in Rats. Neuropsychopharmacology 2019, 44, 295–305. [Google Scholar] [CrossRef] [PubMed]
- Berridge, K.C. Evolving Concepts of Emotion and Motivation. Front. Psychol. 2018, 9, 1647. [Google Scholar] [CrossRef] [Green Version]
- Robinson, D.L.; Zitzman, D.L.; Smith, K.J.; Spear, L.P. Fast Dopamine Release Events in the Nucleus Accumbens of Early Adolescent Rats. Neuroscience 2011, 176, 296–307. [Google Scholar] [CrossRef] [Green Version]
- Dölen, G.; Darvishzadeh, A.; Huang, K.W.; Malenka, R.C. Social Reward Requires Coordinated Activity of Nucleus Accumbens Oxytocin and Serotonin. Nature 2013, 501, 179–184. [Google Scholar] [CrossRef] [Green Version]
- Johnson, Z.V.; Walum, H.; Xiao, Y.; Riefkohl, P.C.; Young, L.J. Oxytocin Receptors Modulate a Social Salience Neural Network in Male Prairie Voles. Horm. Behav. 2017, 87, 16–24. [Google Scholar] [CrossRef] [Green Version]
- De Keyzer, Y.; Auzan, C.; Lenne, F.; Beldjord, C.; Thibonnier, M.; Bertagna, X.; Clauser, E. Cloning and Characterization of the Human V3 Pituitary Vasopressin Receptor. FEBS Lett. 1994, 356, 215–220. [Google Scholar] [CrossRef] [Green Version]
- Sugimoto, T.; Saito, M.; Mochizuki, S.; Watanabe, Y.; Hashimoto, S.; Kawashima, H. Molecular Cloning and Functional Expression of a cDNA Encoding the Human V1b Vasopressin Receptor. J. Biol. Chem. 1994, 269, 27088–27092. [Google Scholar] [CrossRef]
- Song, Z.; McCann, K.E.; McNeill, J.K., 4th; Larkin, T.E., 2nd; Huhman, K.L.; Albers, H.E. Oxytocin Induces Social Communication by Activating Arginine-Vasopressin V1a Receptors and Not Oxytocin Receptors. Psychoneuroendocrinology 2014, 50, 14–19. [Google Scholar] [CrossRef] [Green Version]
- Taylor, J.H.; Cavanaugh, J.; French, J.A. Neonatal Oxytocin and Vasopressin Manipulation Alter Social Behavior during the Juvenile Period in Mongolian Gerbils. Dev. Psychobiol. 2017, 59, 653–657. [Google Scholar] [CrossRef] [PubMed]
- Tan, O.; Musullulu, H.; Raymond, J.S.; Wilson, B.; Langguth, M.; Bowen, M.T. Oxytocin and Vasopressin Inhibit Hyper-Aggressive Behaviour in Socially Isolated Mice. Neuropharmacology 2019, 156, 107573. [Google Scholar] [CrossRef]
- Ross, A.P.; McCann, K.E.; Larkin, T.E.; Song, Z.; Grieb, Z.A.; Huhman, K.L.; Albers, H.E. Sex-Dependent Effects of Social Isolation on the Regulation of Arginine-Vasopressin (AVP) V1a, Oxytocin (OT) and Serotonin (5HT) 1a Receptor Binding and Aggression. Horm. Behav. 2019, 116, 104578. [Google Scholar] [CrossRef] [PubMed]
- Terranova, J.I.; Song, Z.; Larkin, T.E., 2nd; Hardcastle, N.; Norvelle, A.; Riaz, A.; Albers, H.E. Serotonin and Arginine-Vasopressin Mediate Sex Differences in the Regulation of Dominance and Aggression by the Social Brain. Proc. Natl. Acad. Sci. USA 2016, 113, 13233–13238. [Google Scholar] [CrossRef] [Green Version]
- Pagani, J.H.; Zhao, M.; Cui, Z.; Avram, S.K.W.; Caruana, D.A.; Dudek, S.M.; Young, W.S. Role of the Vasopressin 1b Receptor in Rodent Aggressive Behavior and Synaptic Plasticity in Hippocampal Area CA2. Mol. Psychiatry 2015, 20, 490–499. [Google Scholar]
- Bayerl, D.S.; Bosch, O.J. Brain Vasopressin Signaling Modulates Aspects of Maternal Behavior in Lactating Rats. Genes Brain Behav. 2019, 18, e12517. [Google Scholar] [CrossRef] [Green Version]
- Bosch, O.J.; Neumann, I.D. Vasopressin Released within the Central Amygdala Promotes Maternal Aggression. Eur. J. Neurosci. 2010, 31, 883–891. [Google Scholar] [CrossRef]
- Bosch, O.J. Maternal Aggression in Rodents: Brain Oxytocin and Vasopressin Mediate Pup Defence. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2013, 368, 20130085. [Google Scholar] [CrossRef] [Green Version]
- Kessler, M.S.; Bosch, O.J.; Bunck, M.; Landgraf, R.; Neumann, I.D. Maternal Care Differs in Mice Bred for High vs. Low Trait Anxiety: Impact of Brain Vasopressin and Cross-Fostering. Soc. Neurosci. 2011, 6, 156–168. [Google Scholar] [CrossRef]
- Mitre, M.; Marlin, B.J.; Schiavo, J.K.; Morina, E.; Norden, S.E.; Hackett, T.A.; Aoki, C.J.; Chao, M.V.; Froemke, R.C. A Distributed Network for Social Cognition Enriched for Oxytocin Receptors. J. Neurosci. 2016, 36, 2517–2535. [Google Scholar] [CrossRef] [PubMed]
- Marlin, B.J.; Mitre, M.; D’amour, J.A.; Chao, M.V.; Froemke, R.C. Oxytocin Enables Maternal Behaviour by Balancing Cortical Inhibition. Nature 2015, 520, 499–504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakermans-Kranenburg, M.J.; van Ijzendoorn, M.H. Oxytocin Receptor (OXTR) and Serotonin Transporter (5-HTT) Genes Associated with Observed Parenting. Soc. Cogn. Affect. Neurosci. 2008, 3, 128–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiss, I.C.; Pryce, C.R.; Jongen-Rêlo, A.L.; Nanz-Bahr, N.I.; Feldon, J. Effect of Social Isolation on Stress-Related Behavioural and Neuroendocrine State in the Rat. Behav. Brain Res. 2004, 152, 279–295. [Google Scholar] [CrossRef] [PubMed]
- Park, G.; Ryu, C.; Kim, S.; Jeong, S.J.; Koo, J.W.; Lee, Y.-S.; Kim, S.J. Social Isolation Impairs the Prefrontal-Nucleus Accumbens Circuit Subserving Social Recognition in Mice. Cell Rep. 2021, 35, 109104. [Google Scholar] [CrossRef]
- Veenema, A.H.; Bredewold, R.; De Vries, G.J. Sex-Specific Modulation of Juvenile Social Play by Vasopressin. Psychoneuroendocrinology 2013, 38, 2554–2561. [Google Scholar] [CrossRef] [Green Version]
- Paul, M.J.; Terranova, J.I.; Probst, C.K.; Murray, E.K.; Ismail, N.I.; de Vries, G.J. Sexually Dimorphic Role for Vasopressin in the Development of Social Play. Front. Behav. Neurosci. 2014, 8, 58. [Google Scholar] [CrossRef] [Green Version]
- Bredewold, R.; Smith, C.J.W.; Dumais, K.M.; Veenema, A.H. Sex-Specific Modulation of Juvenile Social Play Behavior by Vasopressin and Oxytocin Depends on Social Context. Front. Behav. Neurosci. 2014, 8, 216. [Google Scholar] [CrossRef] [Green Version]
- Lai, M.-C.; Lombardo, M.V.; Baron-Cohen, S. Autism. Lancet 2014, 383, 896–910. [Google Scholar] [CrossRef]
- Modi, M.E.; Young, L.J. The Oxytocin System in Drug Discovery for Autism: Animal Models and Novel Therapeutic Strategies. Horm. Behav. 2012, 61, 340–350. [Google Scholar] [CrossRef] [Green Version]
- Schaafsma, S.M.; Pfaff, D.W. Etiologies Underlying Sex Differences in Autism Spectrum Disorders. Front. Neuroendocrinol. 2014, 35, 255–271. [Google Scholar] [CrossRef]
- Murakami, G.; Hunter, R.G.; Fontaine, C.; Ribeiro, A.; Pfaff, D. Relationships among Estrogen Receptor, Oxytocin and Vasopressin Gene Expression and Social Interaction in Male Mice. Eur. J. Neurosci. 2011, 34, 469–477. [Google Scholar] [CrossRef]
- Willcutt, E.G. The Prevalence of DSM-IV Attention-Deficit/hyperactivity Disorder: A Meta-Analytic Review. Neurotherapeutics 2012, 9, 490–499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Işık, Ü.; Bilgiç, A.; Toker, A.; Kılınç, I. Serum Levels of Cortisol, Dehydroepiandrosterone, and Oxytocin in Children with Attention-Deficit/hyperactivity Disorder Combined Presentation with and without Comorbid Conduct Disorder. Psychiatry Res. 2018, 261, 212–219. [Google Scholar] [CrossRef] [PubMed]
- Enzi, B.; Doering, S.; Faber, C.; Hinrichs, J.; Bahmer, J.; Northoff, G. Reduced Deactivation in Reward Circuitry and Midline Structures during Emotion Processing in Borderline Personality Disorder. World J. Biol. Psychiatry 2013, 14, 45–56. [Google Scholar] [CrossRef]
- Bartz, J.A.; Zaki, J.; Bolger, N.; Ochsner, K.N. Social Effects of Oxytocin in Humans: Context and Person Matter. Trends Cogn. Sci. 2011, 15, 301–309. [Google Scholar] [CrossRef]
- Herpertz, S.C.; Bertsch, K. A New Perspective on the Pathophysiology of Borderline Personality Disorder: A Model of the Role of Oxytocin. Am. J. Psychiatry 2015, 172, 840–851. [Google Scholar] [CrossRef]
- Gunaydin, L.A.; Grosenick, L.; Finkelstein, J.C.; Kauvar, I.V.; Fenno, L.E.; Adhikari, A.; Lammel, S.; Mirzabekov, J.J.; Airan, R.D.; Zalocusky, K.A.; et al. Natural Neural Projection Dynamics Underlying Social Behavior. Cell 2014, 157, 1535–1551. [Google Scholar] [CrossRef] [Green Version]
- Yoshihara, C.; Numan, M.; Kuroda, K.O. Oxytocin and Parental Behaviors. Curr. Top. Behav. Neurosci. 2018, 35, 119–153. [Google Scholar] [PubMed]
- Tapp, D.N.; Singstock, M.D.; Gottliebson, M.S.; McMurray, M.S. Central but Not Peripheral Oxytocin Administration Reduces Risk-Based Decision-Making in Male Rats. Horm. Behav. 2020, 125, 104840. [Google Scholar] [CrossRef]
- Kovàcs, G.L.; Sarnyai, Z.; Barbarczi, E.; Szabó, G.; Telegdy, G. The Role of Oxytocin-Dopamine Interactions in Cocaine-Induced Locomotor Hyperactivity. Neuropharmacology 1990, 29, 365–368. [Google Scholar] [CrossRef]
- Bowen, M.T.; Peters, S.T.; Absalom, N.; Chebib, M.; Neumann, I.D.; McGregor, I.S. Oxytocin Prevents Ethanol Actions at δ Subunit-Containing GABAA Receptors and Attenuates Ethanol-Induced Motor Impairment in Rats. Proc. Natl. Acad. Sci. USA 2015, 112, 3104–3109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burkett, J.P.; Young, L.J. The Behavioral, Anatomical and Pharmacological Parallels between Social Attachment, Love and Addiction. Psychopharmacology 2012, 224, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Heinrichs, M.; Baumgartner, T.; Kirschbaum, C.; Ehlert, U. Social Support and Oxytocin Interact to Suppress Cortisol and Subjective Responses to Psychosocial Stress. Biol. Psychiatry 2003, 54, 1389–1398. [Google Scholar] [CrossRef]
- Ditzen, B.; Schaer, M.; Gabriel, B.; Bodenmann, G.; Ehlert, U.; Heinrichs, M. Intranasal Oxytocin Increases Positive Communication and Reduces Cortisol Levels during Couple Conflict. Biol. Psychiatry 2009, 65, 728–731. [Google Scholar] [CrossRef]
- Yamasue, H.; Okada, T.; Munesue, T.; Kuroda, M.; Fujioka, T.; Uno, Y.; Matsumoto, K.; Kuwabara, H.; Mori, D.; Okamoto, Y.; et al. Effect of Intranasal Oxytocin on the Core Social Symptoms of Autism Spectrum Disorder: A Randomized Clinical Trial. Mol. Psychiatry 2020, 25, 1849–1858. [Google Scholar] [CrossRef] [PubMed]
- Woolley, J.D.; Chuang, B.; Lam, O.; Lai, W.; O’Donovan, A.; Rankin, K.P.; Mathalon, D.H.; Vinogradov, S. Oxytocin Administration Enhances Controlled Social Cognition in Patients with Schizophrenia. Psychoneuroendocrinology 2014, 47, 116–125. [Google Scholar] [CrossRef] [Green Version]
- Parr, L.A. Intranasal Oxytocin Enhances Socially-Reinforced Learning in Rhesus Monkeys. Front. Behav. Neurosci. 2014. [Google Scholar] [CrossRef] [Green Version]
- Gimpl, G.; Fahrenholz, F. The Oxytocin Receptor System: Structure, Function, and Regulation. Physiol. Rev. 2001, 81, 629–683. [Google Scholar] [CrossRef] [Green Version]
- Landgraf, R. Simultaneous Measurement of Arginine Vasopressin and Oxytocin in Plasma and Neurohypophyses by Radioimmunoassay. Endokrinologie 1981, 78, 191–204. [Google Scholar]
- Egashira, N.; Tanoue, A.; Matsuda, T.; Koushi, E.; Harada, S.; Takano, Y.; Tsujimoto, G.; Mishima, K.; Iwasaki, K.; Fujiwara, M. Impaired Social Interaction and Reduced Anxiety-Related Behavior in Vasopressin V1a Receptor Knockout Mice. Behav. Brain Res. 2007, 178, 123–127. [Google Scholar] [CrossRef]
- Manning, M.; Stoev, S.; Chini, B.; Durroux, T.; Mouillac, B.; Guillon, G. Peptide and Non-Peptide Agonists and Antagonists for the Vasopressin and Oxytocin V1a, V1b, V2 and OT Receptors: Research Tools and Potential Therapeutic Agents. Prog. Brain Res. 2008, 170, 473–512. [Google Scholar]
- Xiao, X.; Zhu, Y.; Zhen, D.; Chen, X.M.; Yue, W.; Liu, L.; Li, T. Beneficial and Side Effects of Arginine Vasopressin and Terlipressin for Septic Shock. J. Surg. Res. 2015, 195, 568–579. [Google Scholar] [CrossRef] [PubMed]
- Borie, A.M.; Dromard, Y.; Guillon, G.; Olma, A.; Manning, M.; Muscatelli, F.; Desarménien, M.G.; Jeanneteau, F. Correction of Vasopressin Deficit in the Lateral Septum Ameliorates Social Deficits of Mouse Autism Model. J. Clin. Investig. 2021, 131, e144450. [Google Scholar] [CrossRef] [PubMed]
- Simmons, T.C.; Balland, J.F.; Dhauna, J.; Yang, S.Y.; Traina, J.L.; Vazquez, J.; Bales, K.L. Early Intranasal Vasopressin Administration Impairs Partner Preference in Adult Male Prairie Voles (Microtus ochrogaster). Front. Endocrinol. 2017, 8, 145. [Google Scholar] [CrossRef] [Green Version]
- Price, D.; Burris, D.; Cloutier, A.; Thompson, C.B.; Rilling, J.K.; Thompson, R.R. Dose-Dependent and Lasting Influences of Intranasal Vasopressin on Face Processing in Men. Front. Endocrinol. 2017, 8, 220. [Google Scholar] [CrossRef] [PubMed]
- Parker, K.J.; Oztan, O.; Libove, R.A.; Mohsin, N.; Karhson, D.S.; Sumiyoshi, R.D.; Summers, J.E.; Hinman, K.E.; Motonaga, K.S.; Phillips, J.M.; et al. A Randomized Placebo-Controlled Pilot Trial Shows That Intranasal Vasopressin Improves Social Deficits in Children with Autism. Sci. Transl. Med. 2019, 11, eaau7356. [Google Scholar] [CrossRef]
- Cady, R. A Novel Intranasal Breath-Powered Delivery System for Sumatriptan: A Review of Technology and Clinical Application of the Investigational Product AVP-825 in the Treatment of Migraine. Expert Opin. Drug Deliv. 2015, 12, 1565–1577. [Google Scholar] [CrossRef] [Green Version]
- Bolognani, F.; Del Valle Rubido, M.; Squassante, L.; Wandel, C.; Derks, M.; Murtagh, L.; Sevigny, J.; Khwaja, O.; Umbricht, D.; Fontoura, P. A Phase 2 Clinical Trial of a Vasopressin V1a Receptor Antagonist Shows Improved Adaptive Behaviors in Men with Autism Spectrum Disorder. Sci. Transl. Med. 2019, 11, eaat7838. [Google Scholar] [CrossRef]
- Chao, Y.-S.; McCormack, S. Carbetocin for the Prevention of Post-Partum Hemorrhage: A Review of Clinical Effectiveness, Cost-Effectiveness, and Guidelines; Canadian Agency for Drugs and Technologies in Health: Ottawa, ON, Canada, 2019. [Google Scholar]
- Zanos, P.; Georgiou, P.; Wright, S.R.; Hourani, S.M.; Kitchen, I.; Winsky-Sommerer, R.; Bailey, A. The Oxytocin Analogue Carbetocin Prevents Emotional Impairment and Stress-Induced Reinstatement of Opioid-Seeking in Morphine-Abstinent Mice. Neuropsychopharmacology 2014, 39, 855–865. [Google Scholar] [CrossRef] [Green Version]
- Mairesse, J.; Zinni, M.; Pansiot, J.; Hassan-Abdi, R.; Demene, C.; Colella, M.; Charriaut-Marlangue, C.; Rideau Batista Novais, A.; Tanter, M.; Maccari, S.; et al. Oxytocin Receptor Agonist Reduces Perinatal Brain Damage by Targeting Microglia. Glia 2019, 67, 345–359. [Google Scholar] [CrossRef]
- Moy, S.S.; Teng, B.L., Nikolova; Riddick, N.V.; Simpson, C.D.; Van Deusen, A.; Janzen, W.P.; Sassano, M.F.; Pedersen, C.A.; Jarstfer, M.B. Prosocial Effects of an Oxytocin Metabolite, but Not Synthetic Oxytocin Receptor Agonists, in a Mouse Model of Autism. Neuropharmacology 2019, 144, 301–311. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, A.; Cherepanov, S.M.; Kikuchi, Y.; Fakhrul, A.A.; Akther, S.; Deguchi, K.; Yoshihara, T.; Ishihara, K.; Shuto, S.; Higashida, H. Lipo-Oxytocin-1, a Novel Oxytocin Analog Conjugated with Two Palmitoyl Groups, Has Long-Lasting Effects on Anxiety-Related Behavior and Social Avoidance in CD157 Knockout Mice. Brain Sci. 2015, 5, 3–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cherepanov, S.M.; Akther, S.; Nishimura, T.; Shabalova, A.A.; Mizuno, A.; Ichinose, W.; Shuto, S.; Yamamoto, Y.; Yokoyama, S.; Higashida, H. Effects of Three Lipidated Oxytocin Analogs on Behavioral Deficits in CD38 Knockout Mice. Brain Sci. 2017, 7, 132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Modi, M.E.; Majchrzak, M.J.; Fonseca, K.R.; Doran, A.; Osgood, S.; Vanase-Frawley, M.; Feyfant, E.; McInnes, H.; Darvari, R.; Buhl, D.L.; et al. Peripheral Administration of a Long-Acting Peptide Oxytocin Receptor Agonist Inhibits Fear-Induced Freezing. J. Pharmacol. Exp. Ther. 2016, 358, 164–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huber, D.; Veinante, P.; Stoop, R. Vasopressin and Oxytocin Excite Distinct Neuronal Populations in the Central Amygdala. Science 2005, 308, 245–248. [Google Scholar] [CrossRef] [Green Version]
- Gozzi, A.; Jain, A.; Giovannelli, A.; Bertollini, C.; Crestan, V.; Schwarz, A.J.; Tsetsenis, T.; Ragozzino, D.; Gross, C.T.; Bifone, A. A Neural Switch for Active and Passive Fear. Neuron 2010, 67, 656–666. [Google Scholar] [CrossRef] [Green Version]
- Busnelli, M.; Bulgheroni, E.; Manning, M.; Kleinau, G.; Chini, B. Selective and Potent Agonists and Antagonists for Investigating the Role of Mouse Oxytocin Receptors. J. Pharmacol. Exp. Ther. 2013, 346, 318–327. [Google Scholar] [CrossRef] [Green Version]
- Sala, M.; Braida, D.; Donzelli, A.; Martucci, R.; Busnelli, M.; Bulgheroni, E.; Rubino, T.; Parolaro, D.; Nishimori, K.; Chini, B. Mice Heterozygous for the Oxytocin Receptor Gene (Oxtr(+/-)) Show Impaired Social Behaviour but Not Increased Aggression or Cognitive Inflexibility: Evidence of a Selective Haploinsufficiency Gene Effect. J. Neuroendocrinol. 2013, 25, 107–118. [Google Scholar] [CrossRef]
- Manturewicz, M.; Grzonka, Z.; Borovicková, L.; Slaninová, J. Oxytocin Analogues with Amide Groups Substituted by Tetrazole Groups in Position 4, 5 or 9. Acta Biochim. Pol. 2007, 54, 805–811. [Google Scholar] [CrossRef] [Green Version]
- Sciabola, S.; Goetz, G.H.; Bai, G.; Rogers, B.N.; Gray, D.L.; Duplantier, A.; Fonseca, K.R.; Vanase-Frawley, M.A.; Kablaoui, N.M. Systematic N-Methylation of Oxytocin: Impact on Pharmacology and Intramolecular Hydrogen Bonding Network. Bioorg. Med. Chem. 2016, 24, 3513–3520. [Google Scholar] [CrossRef]
- Pflimlin, E.; Zhou, Z.; Amso, Z.; Fu, Q.; Lee, C.; Muppiddi, A.; Joseph, S.B.; Nguyen-Tran, V.; Shen, W. Engineering a Potent, Long-Acting, and Periphery-Restricted Oxytocin Receptor Agonist with Anorexigenic and Body Weight Reducing Effects. J. Med. Chem. 2020, 63, 382–390. [Google Scholar] [CrossRef] [PubMed]
- Pitt, G.R.W.; Batt, A.R.; Haigh, R.M.; Penson, A.M.; Robson, P.A.; Rooker, D.P.; Tartar, A.L.; Trim, J.E.; Yea, C.M.; Roe, M.B. Non-Peptide Oxytocin Agonists. Bioorg. Med. Chem. Lett. 2004, 14, 4585–4589. [Google Scholar] [CrossRef] [PubMed]
- Becker, J.A.J.; Clesse, D.; Spiegelhalter, C.; Schwab, Y.; Le Merrer, J.; Kieffer, B.L. Autistic-like Syndrome in Mu Opioid Receptor Null Mice Is Relieved by Facilitated mGluR4 Activity. Neuropsychopharmacology 2014, 39, 2049–2060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frantz, M.-C.; Pellissier, L.P.; Pflimlin, E.; Loison, S.; Gandía, J.; Marsol, C.; Durroux, T.; Mouillac, B.; Becker, J.A.J.; Le Merrer, J.; et al. LIT-001, the First Nonpeptide Oxytocin Receptor Agonist That Improves Social Interaction in a Mouse Model of Autism. J. Med. Chem. 2018, 61, 8670–8692. [Google Scholar] [CrossRef]
- Olszewski, P.K.; Ulrich, C.; Ling, N.; Allen, K.; Levine, A.S. A Non-Peptide Oxytocin Receptor Agonist, WAY-267,464, Alleviates Novelty-Induced Hypophagia in Mice: Insights into Changes in c-Fos Immunoreactivity. Pharmacol. Biochem. Behav. 2014, 124, 367–372. [Google Scholar] [CrossRef]
- Hicks, C.; Jorgensen, W.; Brown, C.; Fardell, J.; Koehbach, J.; Gruber, C.W.; Kassiou, M.; Hunt, G.E.; McGregor, I.S. The Nonpeptide Oxytocin Receptor Agonist WAY 267,464: Receptor-Binding Profile, Prosocial Effects and Distribution of c-Fos Expression in Adolescent Rats. J. Neuroendocrinol. 2012, 24, 1012–1029. [Google Scholar] [CrossRef]
- Resendez, S.L.; Namboodiri, V.M.K.; Otis, J.M.; Eckman, L.E.H.; Rodriguez-Romaguera, J.; Ung, R.L.; Basiri, M.L.; Kosyk, O.; Rossi, M.A.; Dichter, G.S.; et al. Social Stimuli Induce Activation of Oxytocin Neurons Within the Paraventricular Nucleus of the Hypothalamus to Promote Social Behavior in Male Mice. J. Neurosci. 2020, 40, 2282–2295. [Google Scholar] [CrossRef]
- Scott, J.E.; Grigsby, P.L.; Hirst, J.J.; Jenkin, G. Inhibition of Prostaglandin Synthesis and Its Effect on Uterine Activity during Established Premature Labor in Sheep. J. Soc. Gynecol. Investig. 2001, 8, 266–276. [Google Scholar] [CrossRef]
- Rasie Abdullahi, P.; Eskandarian, S.; Ghanbari, A.; Rashidy-Pour, A. Oxytocin Receptor Antagonist Atosiban Impairs Consolidation, but Not Reconsolidation of Contextual Fear Memory in Rats. Brain Res. 2018, 1695, 31–36. [Google Scholar] [CrossRef]
- Liu, R.; Yuan, X.; Chen, K.; Jiang, Y.; Zhou, W. Perception of Social Interaction Compresses Subjective Duration in an Oxytocin-Dependent Manner. Elife 2018, 7, e32100. [Google Scholar] [CrossRef]
- Erfanparast, A.; Tamaddonfard, E.; Henareh-Chareh, F. Intra-Hippocampal Microinjection of Oxytocin Produced Antiepileptic Effect on the Pentylenetetrazol-Induced Epilepsy in Rats. Pharmacol. Rep. 2017, 69, 757–763. [Google Scholar] [CrossRef]
- Lukmanji, S.; Manji, S.A.; Kadhim, S.; Sauro, K.M.; Wirrell, E.C.; Kwon, C.-S.; Jetté, N. The Co-Occurrence of Epilepsy and Autism: A Systematic Review. Epilepsy Behav. 2019, 98, 238–248. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.C.; Shapiro, L.; Thelin, J.T.; Heaton, E.C.; Zaman, R.U.; D’Souza, M.J.; Murnane, K.S.; Escayg, A. Nanoparticle Encapsulated Oxytocin Increases Resistance to Induced Seizures and Restores Social Behavior in Scn1a-Derived Epilepsy. Neurobiol. Dis. 2021, 147, 105147. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Hou, W.; Hao, X.; Dong, N.; Du, P.; Yuan, W.; Yang, J.; Jia, R.; Tai, F. Oxytocin Receptor Antagonist Treatments Alter Levels of Attachment to Mothers and Central Dopamine Activity in Pre-Weaning Mandarin Vole Pups. Psychoneuroendocrinology 2017, 84, 124–134. [Google Scholar] [CrossRef] [PubMed]
- Lubin, D.A.; Elliott, J.C.; Black, M.C.; Johns, J.M. An Oxytocin Antagonist Infused into the Central Nucleus of the Amygdala Increases Maternal Aggressive Behavior. Behav. Neurosci. 2003, 117, 195–201. [Google Scholar] [CrossRef] [Green Version]
- Johns, J.M.; Noonan, L.R.; Zimmerman, L.I.; Li, L.; Pedersen, C.A. Effects of Short- and Long-Term Withdrawal from Gestational Cocaine Treatment on Maternal Behavior and Aggression in Sprague-Dawley Rats. Dev. Neurosci. 1997, 19, 368–374. [Google Scholar] [CrossRef]
- Yu, C.J.; Zhang, S.W.; Tai, F.D. Effects of Nucleus Accumbens Oxytocin and Its Antagonist on Social Approach Behavior. Behav. Pharmacol. 2016, 27, 672–680. [Google Scholar] [CrossRef]
- Boccia, M.L.; Goursaud, A.P.; Bachevalier, J.; Anderson, K.D.; Pedersen, C.A. Peripherally Administered Non-Peptide Oxytocin Antagonist, L368899®, Accumulates in Limbic Brain Areas: A New Pharmacological Tool for the Study of Social Motivation in Non-Human Primates. Horm. Behav. 2007, 52, 344–351. [Google Scholar] [CrossRef] [Green Version]
- Blitzer, D.S.; Wells, T.E.; Hawley, W.R. Administration of an Oxytocin Receptor Antagonist Attenuates Sexual Motivation in Male Rats. Horm. Behav. 2017, 94, 33–39. [Google Scholar] [CrossRef]
- Kim, E.J.; Pellman, B.; Kim, J.J. Stress Effects on the Hippocampus: A Critical Review. Learn. Mem. 2015, 22, 411–416. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.-Y.; Park, S.-H.; Chung, C.; Kim, J.J.; Choi, S.-Y.; Han, J.-S. Oxytocin Protects Hippocampal Memory and Plasticity from Uncontrollable Stress. Sci. Rep. 2015, 5, 18540. [Google Scholar] [CrossRef] [PubMed]
- Cragg, B.; Ji, G.; Neugebauer, V. Differential Contributions of Vasopressin V1A and Oxytocin Receptors in the Amygdala to Pain-Related Behaviors in Rats. Mol. Pain 2016, 12, 1744806916676491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gross, P.A.; Wagner, A.; Decaux, G. Vaptans Are Not the Mainstay of Treatment in Hyponatremia: Perhaps Not yet. Kidney Int. 2011, 80, 594–600. [Google Scholar] [CrossRef] [PubMed]
- Manning, M.; Misicka, A.; Olma, A.; Bankowski, K.; Stoev, S.; Chini, B.; Durroux, T.; Mouillac, B.; Corbani, M.; Guillon, G. Oxytocin and Vasopressin Agonists and Antagonists as Research Tools and Potential Therapeutics. J. Neuroendocrinol. 2012, 24, 609–628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serradeil-Le Gal, C.; Wagnon, J.; Simiand, J.; Griebel, G.; Lacour, C.; Guillon, G.; Barberis, C.; Brossard, G.; Soubrié, P.; Nisato, D.; et al. Characterization of (2S,4R)-1-[5-Chloro-1-[(2,4-Dimethoxyphenyl)sulfonyl]-3-(2-Methoxy-Phenyl)-2-Oxo-2,3-Dihydro-1H-Indol-3-Yl]-4-Hydroxy-N,N-Dimethyl-2-Pyrrolidine Carboxamide (SSR149415), a Selective and Orally Active Vasopressin V1b Receptor Antagonist. J. Pharmacol. Exp. Ther. 2002, 300, 1122–1130. [Google Scholar]
- Griebel, G.; Beeské, S.; Stahl, S.M. The Vasopressin V(1b) Receptor Antagonist SSR149415 in the Treatment of Major Depressive and Generalized Anxiety Disorders: Results from 4 Randomized, Double-Blind, Placebo-Controlled Studies. J. Clin. Psychiatry 2012, 73, 1403–1411. [Google Scholar] [CrossRef]
- Kirchhoff, V.D.; Nguyen, H.T.T.; Soczynska, J.K.; Woldeyohannes, H.; McIntyre, R.S. Discontinued Psychiatric Drugs in 2008. Expert Opin. Investig. Drugs 2009, 18, 1431–1443. [Google Scholar]
- AbbVie Deutschland GmbH & Co., KG. 5,6-Disubstituted Oxindole-Derivatives and Use Thereof for Treating Vasopress Independent Diseases. U.S. Patent 8546401, 1 October 2013. [Google Scholar]
- AbbVie Deutschland GmbH & Co., KG. Substituted Oxindole-Derivatives and the Use Thereof for the Treatment of Vasopressin-dependent Illnesses. U.S. Patent 8629148, 14 January 2014. [Google Scholar]
- Abbott Gmbh & Co., KG. Substituted Oxindole Derivatives and their Use as Vasopressin Receptor Ligands. U.S. Patent 0322809, 14 October 2014. [Google Scholar]
- Abbott Gmbh & Co., KG. Heteroaryl-Substituted 1,3-Dihydroindol-2-One Derivatives and Medicaments Containing Them. U.S. Patent 8580842, 12 November 2013. [Google Scholar]
- Porsolt, R.D.; Bertin, A.; Jalfre, M. “Behavioural Despair” in Rats and Mice: Strain Differences and the Effects of Imipramine. Eur. J. Pharmacol. 1978, 51, 291–294. [Google Scholar] [CrossRef]
- Ferris, C.F.; Lu, S.-F.; Messenger, T.; Guillon, C.D.; Heindel, N.; Miller, M.; Koppel, G.; Robert Bruns, F.; Simon, N.G. Orally Active Vasopressin V1a Receptor Antagonist, SRX251, Selectively Blocks Aggressive Behavior. Pharmacol. Biochem. Behav. 2006, 83, 169–174. [Google Scholar] [CrossRef]
- Lago, T.R.; Brownstein, M.J.; Page, E.; Beydler, E.; Manbeck, A.; Beale, A.; Roberts, C.; Balderston, N.; Damiano, E.; Pineles, S.L.; et al. The Novel Vasopressin Receptor (V1aR) Antagonist SRX246 Reduces Anxiety in an Experimental Model in Humans: A Randomized Proof-of-Concept Study. Psychopharmacology 2021, 238, 2393–2403. [Google Scholar] [CrossRef]
- Roth, B.L.; Sheffler, D.J.; Kroeze, W.K. Magic Shotguns versus Magic Bullets: Selectively Non-Selective Drugs for Mood Disorders and Schizophrenia. Nat. Rev. Drug Discov. 2004, 3, 353–359. [Google Scholar] [CrossRef] [Green Version]
- Reyes-Parada, M.; Iturriaga-Vasquez, P. The Development of Novel Polypharmacological Agents Targeting the Multiple Binding Sites of Nicotinic Acetylcholine Receptors. Expert Opin. Drug Discov. 2016, 11, 969–981. [Google Scholar] [CrossRef] [PubMed]
- Medina-Franco, J.L.; Giulianotti, M.A.; Welmaker, G.S.; Houghten, R.A. Shifting from the Single to the Multitarget Paradigm in Drug Discovery. Drug Discov. Today 2013, 18, 495–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravikumar, B.; Aittokallio, T. Improving the Efficacy-Safety Balance of Polypharmacology in Multi-Target Drug Discovery. Expert Opin. Drug Discov. 2018, 13, 179–192. [Google Scholar] [CrossRef]
- Mei, Y.; Yang, B. Rational Application of Drug Promiscuity in Medicinal Chemistry. Future Med. Chem. 2018, 10, 1835–1851. [Google Scholar] [CrossRef]
- Iturriaga-Vásquez, P.; Alzate-Morales, J.; Bermudez, I.; Varas, R.; Reyes-Parada, M. Multiple Binding Sites in the Nicotinic Acetylcholine Receptors: An Opportunity for Polypharmacolgy. Pharmacol. Res. 2015, 101, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Papke, R.L.; Lindstrom, J.M. Nicotinic Acetylcholine Receptors: Conventional and Unconventional Ligands and Signaling. Neuropharmacology 2020, 168, 108021. [Google Scholar] [CrossRef]
- Hoyer, D.; Bartfai, T. Neuropeptides and Neuropeptide Receptors: Drug Targets, and Peptide and Non-Peptide Ligands: A Tribute to Prof. Dieter Seebach. Chem. Biodivers. 2012, 9, 2367–2387. [Google Scholar] [CrossRef]
- Eiden, L.E.; Gundlach, A.L.; Grinevich, V.; Lee, M.R.; Mecawi, A.S.; Chen, D.; Buijs, R.M.; Hernandez, V.S.; Fajardo-Dolci, G.; Zhang, L. Regulatory Peptides and Systems Biology: A New Era of Translational and Reverse-Translational Neuroendocrinology. J. Neuroendocrinol. 2020, 32, e12844. [Google Scholar] [CrossRef]
- Rodríguez, B.; Nani, J.V.; Almeida, P.G.C.; Brietzke, E.; Lee, R.S.; Hayashi, M.A.F. Neuropeptides and Oligopeptidases in Schizophrenia. Neurosci. Biobehav. Rev. 2020, 108, 679–693. [Google Scholar] [CrossRef] [PubMed]
- Griebel, G.; Simiand, J.; Serradeil-Le Gal, C.; Wagnon, J.; Pascal, M.; Scatton, B.; Maffrand, J.-P.; Soubrie, P. Anxiolytic- and Antidepressant-like Effects of the Non-Peptide Vasopressin V1b Receptor Antagonist, SSR149415, Suggest an Innovative Approach for the Treatment of Stress-Related Disorders. Proc. Natl. Acad. Sci. USA 2002, 99, 6370–6375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griebel, G.; Simiand, J.; Stemmelin, J.; Gal, C.S.-L.; Steinberg, R. The Vasopressin V1b Receptor as a Therapeutic Target in Stress-Related Disorders. Curr. Drug Targets CNS Neurol. Disord. 2003, 2, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Tunstall, B.J.; Kirson, D.; Zallar, L.J.; McConnell, S.A.; Vendruscolo, J.C.M.; Ho, C.P.; Oleata, C.S.; Khom, S.; Manning, M.; Lee, M.R.; et al. Oxytocin Blocks Enhanced Motivation for Alcohol in Alcohol Dependence and Blocks Alcohol Effects on GABAergic Transmission in the Central Amygdala. PLoS Biol. 2019, 17, e2006421. [Google Scholar] [CrossRef] [PubMed]
- Everett, N.A.; McGregor, I.S.; Baracz, S.J.; Cornish, J.L. The Role of the Vasopressin V1A Receptor in Oxytocin Modulation of Methamphetamine Primed Reinstatement. Neuropharmacology 2018, 133, 1–11. [Google Scholar] [CrossRef]


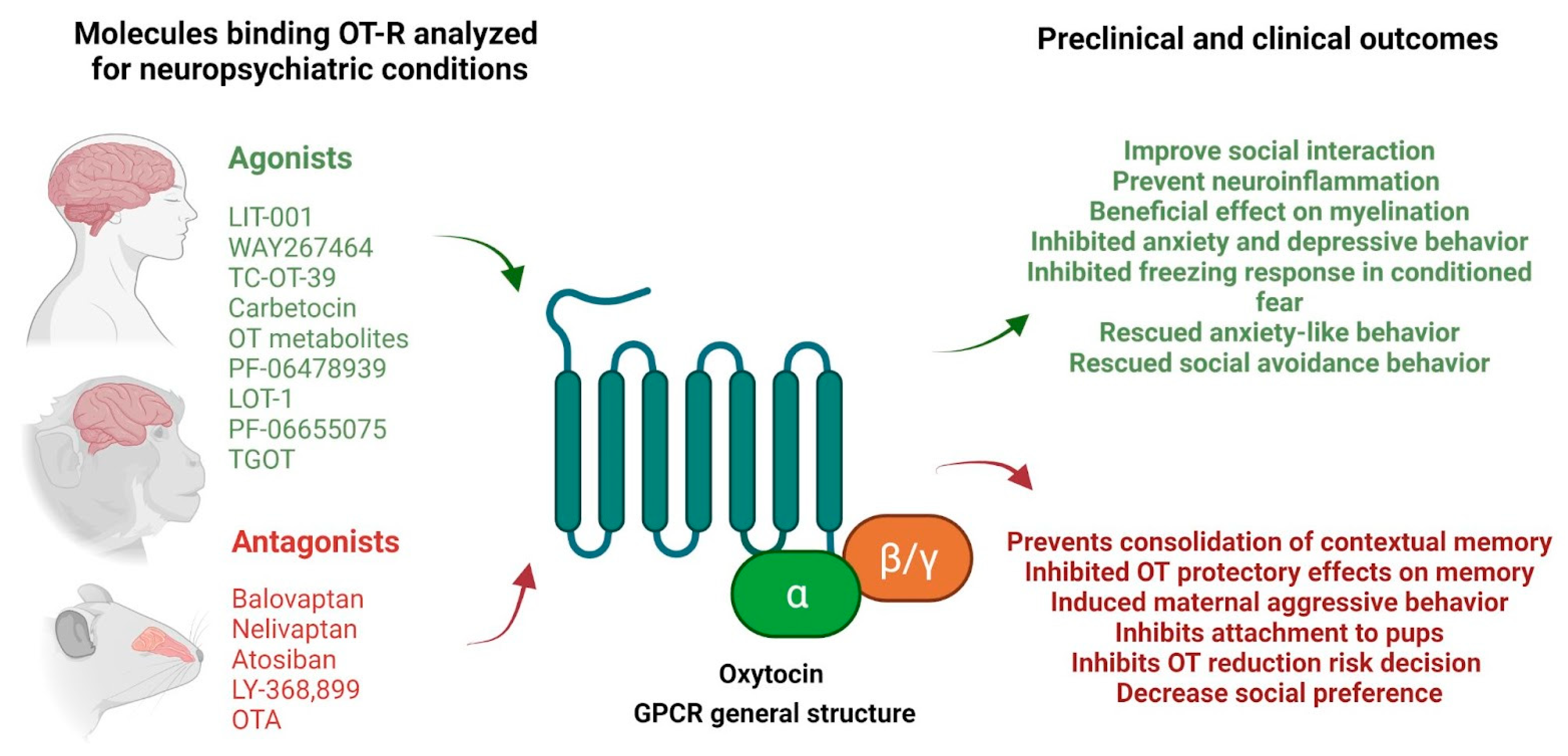{kind=link}
| Molecule | OTR | V1A | V1B | Principal Outcomes Reported | Model/Disease/Condition | Refs. |
|---|---|---|---|---|---|---|
| RG7314 or RO5285119 or RAX5D5AGV6 or balovaptan | Antagonist (np) | Antagonist (np) | Antagonist (np) | Improvements in the Vineland-II socialization and communication scores. | Humans with ASD | [163] |
| SSR149415 or nelivaptan | antagonist (+) | antagonist (+) | antagonist (++) | Antagonized exogenous AVP-induced corticotropin secretion. Rats pretreated with nelivaptan induced inhibition in plasma corticotropin secretion and 30 min before the stress period caused a 50% inhibition of plasma corticotropin elevation. | Male Sprague Dawley Rats | [201] |
| The forced swimming test produced a lower immobility time than the vehicle. | Wistar rats | [221] | ||||
| After a chronic mild stress test, degradation of the physical state of the animal’s coat was significantly improved by nelivaptan after 2 weeks of treatment. | BALB/c mice | [221,222] | ||||
| In generalized anxiety (GAD) and major depressive disorder (MDD), the treated patients did not show significant improvement from the baseline or did not separate from the placebo. | Patients with DSM criteria of GAD and MDD | [202] | ||||
| SRX251 | (-) | Antagonist (np) | (-) | Dose-dependent reduction in aggression | Male Syrian golden hamsters; resident-intruder model of aggression | [209] |
| SRX246 | (-) | Antagonist (np) | (-) | Decreased anxiety-potentiated startle independent of fear-potentiated startle | Healthy volunteers | [210] |
| LIT-001 | agonist (no-peptide) (++) | Antagonist (+) | Agonist to (-) high concentration | Improves social interaction. The activation of V1BR does not alter social behavior | Rat | [180] |
| WAY267464 | agonist (+++) | Antagonist (+) | (-) | Improves social behavior in the SHANK-3B model Antagonism V1BRfacilitates social interaction | Mice model of ASD (OPRM1 -/-) | [183] |
| TC-0T-39 | Agonist (+++) | (-) | (-) | Prosocial efficacy | BALB/cByJ model mice (ASD model) | [167] |
| Carbetocin | Agonist (+++) | (-) | (-) | No prosocial efficacy | BALB/cByJ model mice (ASD model) | [167] |
| Prevent neuroinflammation-induced brain damage of perinatal origin Beneficial effect on myelination, intrinsic brain connectivity | Rat model of perinatal brain injury (low gestational protein diet LPD) | [166] | ||||
| Administered peripherally, inhibited the development of anxiety and depressive behaviors during morphine withdrawal | Addiction mouse model | [165] | ||||
| Metabolites OT (4-9) and (5-9) | Oxytocin analogs | (-) | (-) | OT (4-9). Improves social preference, dose-dependent manner | BALB/cByJ (mice model of ASD) | [167] |
| PF-06478939 | Agonist (+++) | (-) | (-) | Peripheral administration inhibited freezing in response to the conditioned fear stimulus | Rats, conditioned fear paradigm | [170] |
| LOT-1 | Agonist (+++) | (-) | (-) | Rescued anxiety-like behavior and social avoidance in the open field test | CD157 knockout model mouse of the non-motor psychiatric symptoms of Parkinson’s disease | [168] |
| Atosiban | Antagonist (++) | Antagonist | (-) | OT-R. Prevents the consolidation of contextual memory to fear in rats | Rats, contextual fear memory paradigm | [185] |
| V1AR. The microinjection into the hippocampus prevents the antiepileptic effect induced by diazepam | [187] | |||||
| PF-06655075 | Agonist (+++) | (-) | (-) | Decreased alcohol drinking | Rat model of alcohol dependence | [223] |
| SR49059 or relcovaptan | (-) | Antagonist | (-) | Inhibited vocalizations and anxiety-like behavior (elevated plus maze) in arthritic, but not normal, rats and conveyed anxiolytic properties to arginine vasopressin | Rats. Arthritic model (kaolin/carrageenan knee joint pain model) | [198] |
| Inhibited anti-aggressive effects of OT in mice after 6 weeks of isolation | male Swiss mice | [117] | ||||
| Blocks the effect of OT in the inhibitory on METH-primed reinstatement of METH-seeking behavior | Male Rats trained to self-administer | [224] | ||||
| LY371257 | Antagonist (+++) | (-) | (-) | Facilitatory effects on vocalizations | Rats. Arthritic model (kaolin/carrageenan knee joint pain model) | [198] |
| LY-368,899 | Antagonist (+++) | (-) | (-) | Reduced interest in the infant (primate maternal interest) and sexual behavior | Rhesus Monkeys | [194,195] |
| Inhibited the oxytocin’s protective effects on hippocampal memory to stress | Model stress in rats | [197] | ||||
| OTA | Antagonist (+++) | (-) | (-) | Induced maternal aggressive behavior | Female Rats | [191] |
| Inhibits the attachment of pre-weaning pups to mothers | Male and female pre-weaning mandarin voles (Microtus mandarinus) | [190] | ||||
| Inhibits the role of oxytocin in selectively reducing risk decisions in male rats | Rats. Probability discounting task | [144] | ||||
| Decreased social preferences in volved in a dose-dependent manner | Female and male monogamous mandarin voles (Microtus mandarinus) using the social preference paradigm | [193] | ||||
| TGOT | Agonist (+++) | (-) | (-) | Reduced isolation-induced aggression | Mice, social isolation induced aggression paradigm | [117] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cid-Jofré, V.; Moreno, M.; Reyes-Parada, M.; Renard, G.M. Role of Oxytocin and Vasopressin in Neuropsychiatric Disorders: Therapeutic Potential of Agonists and Antagonists. Int. J. Mol. Sci. 2021, 22, 12077. https://doi.org/10.3390/ijms222112077
Cid-Jofré V, Moreno M, Reyes-Parada M, Renard GM. Role of Oxytocin and Vasopressin in Neuropsychiatric Disorders: Therapeutic Potential of Agonists and Antagonists. International Journal of Molecular Sciences. 2021; 22(21):12077. https://doi.org/10.3390/ijms222112077
Chicago/Turabian StyleCid-Jofré, Valeska, Macarena Moreno, Miguel Reyes-Parada, and Georgina M. Renard. 2021. "Role of Oxytocin and Vasopressin in Neuropsychiatric Disorders: Therapeutic Potential of Agonists and Antagonists" International Journal of Molecular Sciences 22, no. 21: 12077. https://doi.org/10.3390/ijms222112077