
Tks5 Regulates Synaptic Podosome Formation and Stabilization of the Postsynaptic Machinery at the Neuromuscular Junction

 and
and 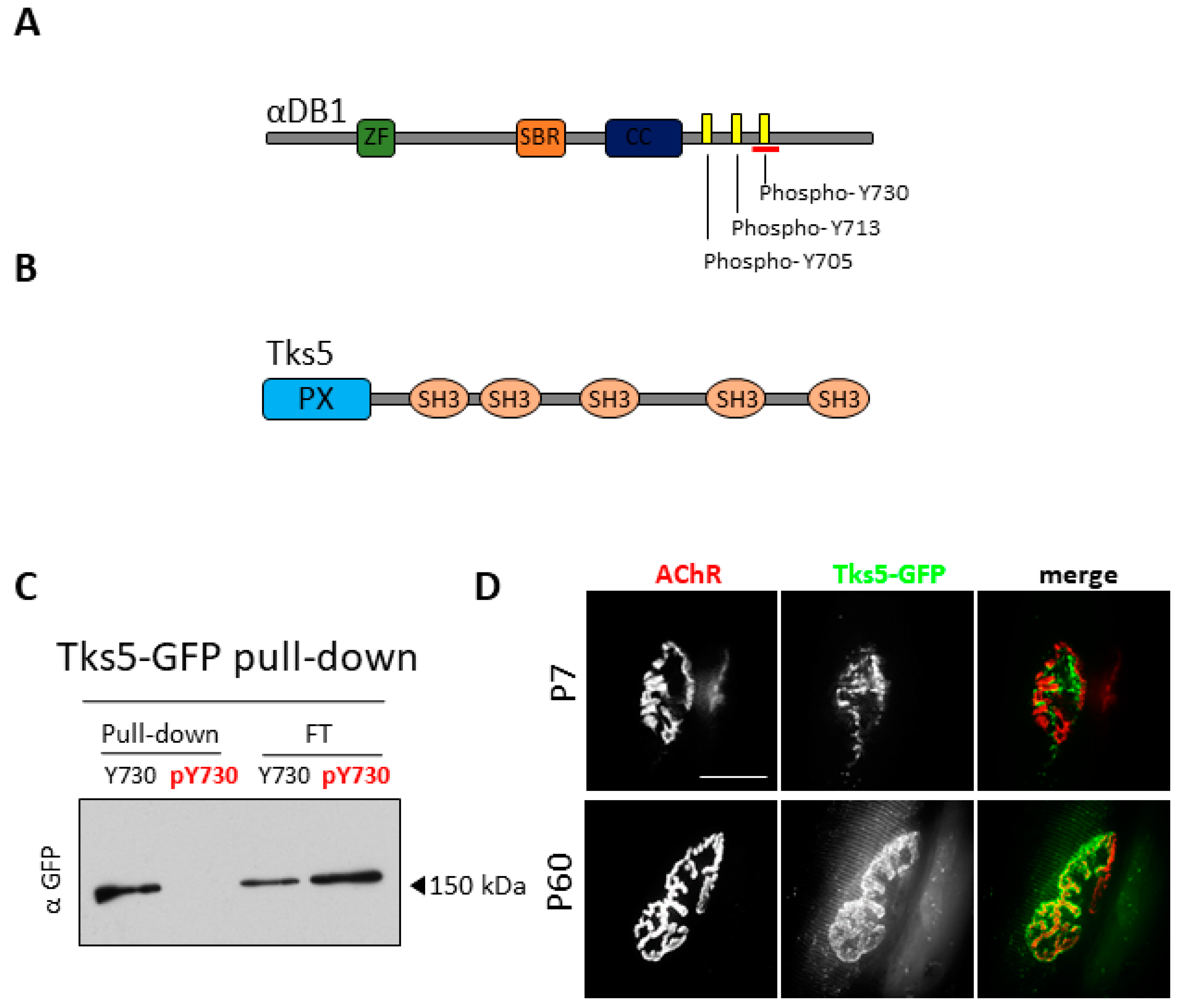{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Tks5 Interacts with Unphosphorylated Adystrobrevin-1 and Localizes to the NMJ
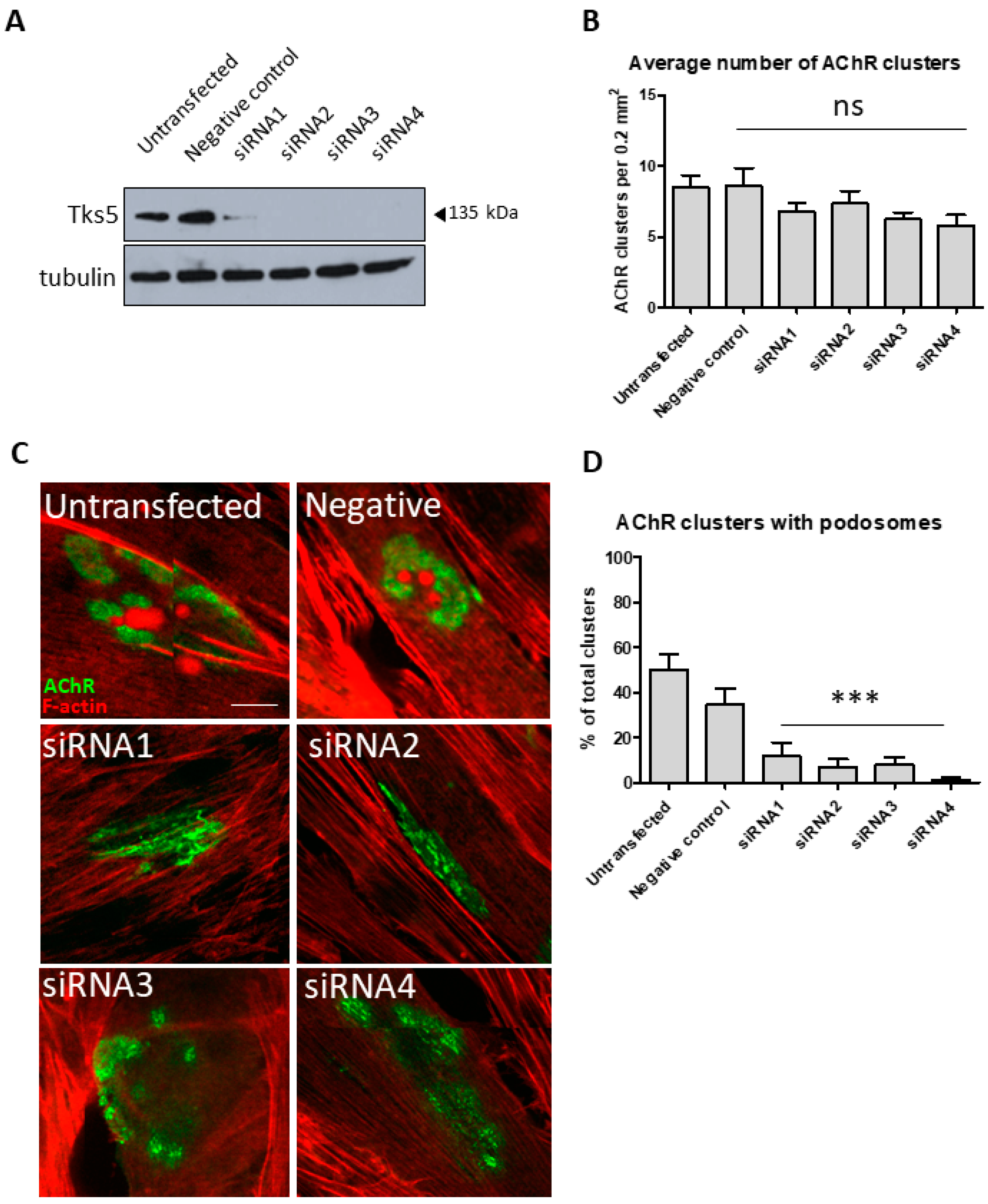
2.2. Tks5 Is Essential for Synaptic Podosome Formation
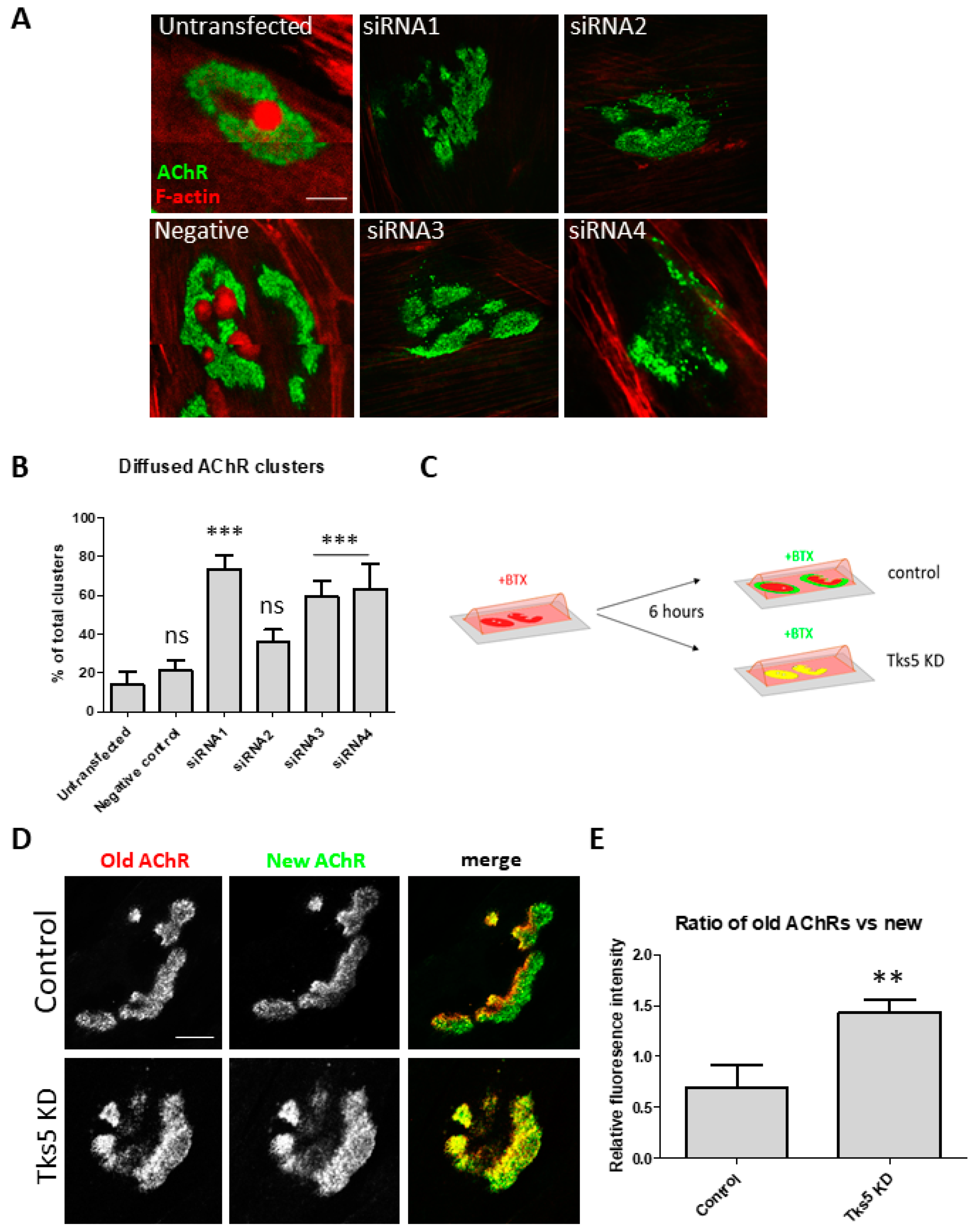
2.3. Absence of Tks5 Leads to Instability of AChR Clusters and Compromised Turnover of the Receptors
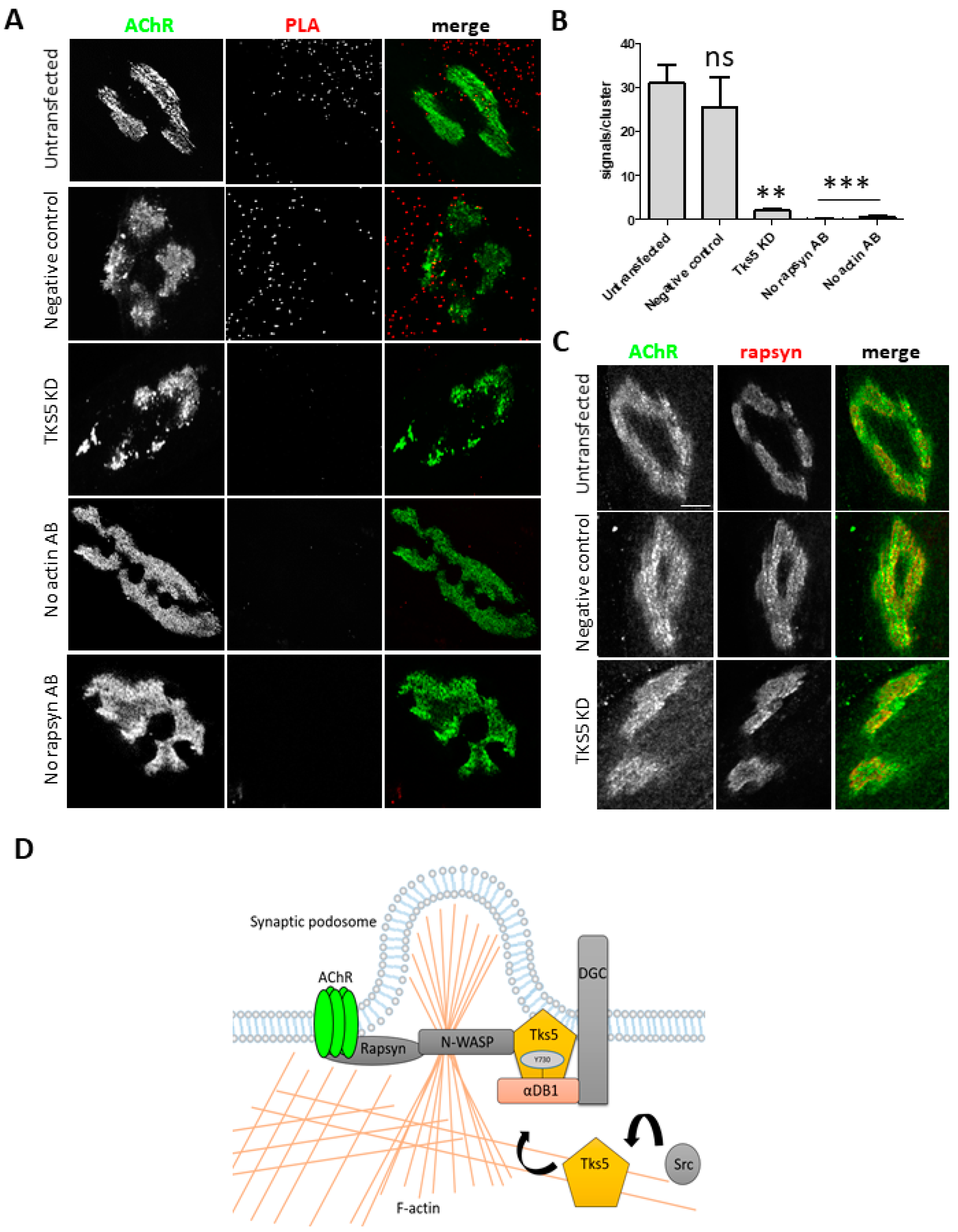
2.4. Tks5 Is Required for Association of Actin with the Postsynaptic Machinery
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Cell Transfection
4.3. Immunocytochemistry and Immunohistochemistry
4.4. Microscopy and Image Processing
4.5. Proximity Ligation Assay (PLA)
4.6. Protein Pulldown
4.7. Mass Spectrometry
4.8. Mice Sacrifice and Tissue Extraction
4.9. Muscle Electroporation
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nishimune, H.; Shigemoto, K. Practical Anatomy of the Neuromuscular Junction in Health and Disease. Neurol. Clin. 2018, 36, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Slater, C.R. The functional organization of motor nerve terminals. Prog. Neurobiol. 2015, 134, 55–103. [Google Scholar] [CrossRef] [PubMed]
- Verschuuren, J.; Strijbos, E.; Vincent, A. Neuromuscular junction disorders. Handb. Clin. Neurol. 2016, 133, 447–466. [Google Scholar] [CrossRef]
- Steinbach, J.H. Developmental changes in acetylcholine receptor aggregates at rat skeletal neuromuscular junctions. Dev. Biol. 1981, 84, 267–276. [Google Scholar] [CrossRef]
- Marques, M.J.; Conchello, J.-A.; Lichtman, J.W. From Plaque to Pretzel: Fold Formation and Acetylcholine Receptor Loss at the Developing Neuromuscular Junction. J. Neurosci. 2000, 20, 3663–3675. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.I.; Mikesh, M.; Smith, I.; Rimer, M.; Thompson, W. Muscles in a mouse model of spinal muscular atrophy show profound defects in neuromuscular development even in the absence of failure in neuromuscular transmission or loss of motor neurons. Dev. Biol. 2011, 356, 432–444. [Google Scholar] [CrossRef] [Green Version]
- Kang, H.; Tian, L.; Son, Y.-J.; Zuo, Y.; Procaccino, D.; Love, F.; Hayworth, C.; Trachtenberg, J.; Mikesh, M.; Sutton, L.; et al. Regulation of the Intermediate Filament Protein Nestin at Rodent Neuromuscular Junctions by Innervation and Activity. J. Neurosci. 2007, 27, 5948–5957. [Google Scholar] [CrossRef]
- Pęziński, M.; Prószyński, T. Domain organization of synaptic components. Curr. Opin. Physiol. 2018, 4, 7–14. [Google Scholar] [CrossRef]
- Proszynski, T.J.; Gingras, J.; Valdez, G.; Krzewski, K.; Sanes, J.R. Podosomes are present in a postsynaptic apparatus and participate in its maturation. Proc. Natl. Acad. Sci. USA 2009, 106, 18373–18378. [Google Scholar] [CrossRef] [Green Version]
- Bernadzki, K.M.; Rojek, K.O.; Prószyński, T.J. Podosomes in muscle cells and their role in the remodeling of neuromuscular postsynaptic machinery. Eur. J. Cell Biol. 2014, 93, 478–485. [Google Scholar] [CrossRef]
- Chan, Z.C.-K.; Kwan, H.L.R.; Wong, Y.S.; Jiang, Z.; Zhou, Z.; Tam, K.W.; Chan, Y.-S.; Chan, C.B.; Lee, C.W. Site-directed MT1-MMP trafficking and surface insertion regulate AChR clustering and remodeling at developing NMJs. eLife 2020, 9, 9. [Google Scholar] [CrossRef]
- Linder, S.; Cervero, P. The podosome cap: Past, present, perspective. Eur. J. Cell Biol. 2020, 99, 151087. [Google Scholar] [CrossRef]
- Proszynski, T.J.; Sanes, J.R. Amotl2 interacts with LL5β, localizes to podosomes and regulates postsynaptic differentiation in muscle. J. Cell Sci. 2013, 126, 2225–2235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kishi, M.; Kummer, T.T.; Eglen, S.J.; Sanes, J.R. LL5beta: A regulator of postsynaptic differentiation identified in a screen for synaptically enriched transcripts at the neuromuscular junction. J. Cell Biol. 2005, 169, 355–366. [Google Scholar] [CrossRef]
- Gawor, M.; Prószyński, T.J. The molecular cross talk of the dystrophin-glycoprotein complex. Ann. N. Y. Acad. Sci. 2017, 1412, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Nishimune, H.; Valdez, G.; Jarad, G.; Moulson, C.L.; Müller, U.; Miner, J.H.; Sanes, J.R. Laminins promote postsynaptic maturation by an autocrine mechanism at the neuromuscular junction. J. Cell Biol. 2008, 182, 1201–1215. [Google Scholar] [CrossRef] [Green Version]
- Belhasan, D.C.; Akaaboune, M. The role of the dystrophin glycoprotein complex on the neuromuscular system. Neurosci. Lett. 2020, 722, 134833. [Google Scholar] [CrossRef]
- Nakamori, M.; Kimura, T.; Kubota, T.; Matsumura, T.; Sumi, H.; Fujimura, H.; Takahashi, M.P.; Sakoda, S. Aberrantly spliced -dystrobrevin alters -syntrophin binding in myotonic dystrophy type 1. Neurology 2008, 70, 677–685. [Google Scholar] [CrossRef] [PubMed]
- Grady, R.M.; Grange, R.W.; Lau, K.S.; Maimone, M.M.; Nichol, M.C.; Stull, J.T.; Sanes, J.R. Role for α-dystrobrevin in the pathogenesis of dystrophin-dependent muscular dystrophies. Nat. Cell Biol. 1999, 1, 215–220. [Google Scholar] [CrossRef]
- Bernadzki, K.M.; Daszczuk, P.; Rojek, K.O.; Pęziński, M.; Gawor, M.; Pradhan, B.; De Cicco, T.; Bijata, M.; Bijata, K.; Włodarczyk, J.; et al. Arhgef5 Binds α-Dystrobrevin 1 and Regulates Neuromuscular Junction Integrity. Front. Mol. Neurosci. 2020, 13, 104. [Google Scholar] [CrossRef] [PubMed]
- Gingras, J.; Gawor, M.; Bernadzki, K.M.; Grady, R.M.; Hallock, P.; Glass, D.J.; Sanes, J.R.; Prószyński, T.J. Alpha-Dystrobrevin-1 recruits Grb2 and alpha-catulin to organize neurotransmitter receptors at the neuromuscular junction. J. Cell Sci. 2016, 129, 898–911. [Google Scholar] [PubMed] [Green Version]
- Bernadzki, K.M.; Gawor, M.; Pęziński, M.; Mazurek, P.; Niewiadomski, P.; Rędowicz, M.J.; Prószyński, T.J. Liprin-α-1 is a novel component of the murine neuromuscular junction and is involved in the organization of the postsynaptic machinery. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grady, R.M.; Akaaboune, M.; Cohen, A.L.; Maimone, M.M.; Lichtman, J.W.; Sanes, J.R. Tyrosine-phosphorylated and nonphosphorylated isoforms of alpha-dystrobrevin: Roles in skeletal muscle and its neuromuscular and myotendinous junctions. J. Cell Biol. 2003, 160, 741–752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pawlikowski, B.T.; Maimone, M.M. Formation of complex AChR aggregates in vitro requires alpha-dystrobrevin. Dev. Neurobiol. 2009, 69, 326–338. [Google Scholar] [CrossRef]
- Seals, D.F.; Azucena, E.F.; Pass, I.; Tesfay, L.; Gordon, R.; Woodrow, M.; Resau, J.H.; Courtneidge, S.A. The adaptor protein Tks5/Fish is required for podosome formation and function, and for the protease-driven invasion of cancer cells. Cancer Cell 2005, 7, 155–165. [Google Scholar] [CrossRef] [Green Version]
- Courtneidge, S.A.; Azucena, E.F.; Pass, I.; Seals, D.F.; Tesfay, L. The SRC substrate Tks5, podosomes (invadopodia), and cancer cell invasion. Cold Spring Harb. Symp. Quant. Biol. 2005, 70, 167–171. [Google Scholar] [CrossRef] [Green Version]
- Thompson, O.; Kleino, I.; Crimaldi, L.; Gimona, M.; Saksela, K.; Winder, S.J. Dystroglycan, Tks5 and Src Mediated Assembly of Podosomes in Myoblasts. PLoS ONE 2008, 3, e3638. [Google Scholar] [CrossRef] [Green Version]
- Oikawa, T.; Itoh, T.; Takenawa, T. Sequential signals toward podosome formation in NIH-src cells. J. Cell Biol. 2008, 182, 157–169. [Google Scholar] [CrossRef]
- Muntoni, F.; Torelli, S.; Ferlini, A. Dystrophin and mutations: One gene, several proteins, multiple phenotypes. Lancet Neurol. 2003, 2, 731–740. [Google Scholar] [CrossRef]
- Kummer, T.; Misgeld, T.; Lichtman, J.W.; Sanes, J.R. Nerve-independent formation of a topologically complex postsynaptic apparatus. J. Cell Biol. 2004, 164, 1077–1087. [Google Scholar] [CrossRef] [Green Version]
- Alam, M.S. Proximity Ligation Assay (PLA). Curr. Protoc. Immunol. 2018, 123, e58. [Google Scholar] [CrossRef]
- Lin, M.; Xiong, W.-C.; Mei, L. Neuromuscular Junction Formation, Aging, and Disorders. Annu. Rev. Physiol. 2018, 80, 159–188. [Google Scholar] [CrossRef]
- Murphy, D.A.; Courtneidge, S.A. The ‘ins’ and ‘outs’ of podosomes and invadopodia: Characteristics, formation and function. Nat. Rev. Mol. Cell Biol. 2011, 12, 413–426. [Google Scholar] [CrossRef] [Green Version]
- Gimona, M.; Buccione, R.; Courtneidge, S.A.; Linder, S. Assembly and biological role of podosomes and invadopodia. Curr. Opin. Cell Biol. 2008, 20, 235–241. [Google Scholar] [CrossRef]
- Abram, C.L.; Seals, D.F.; Pass, I.; Salinsky, D.; Maurer, L.; Roth, T.M.; Courtneidge, S.A. The Adaptor Protein Fish Associates with Members of the ADAMs Family and Localizes to Podosomes of Src-transformed Cells. J. Biol. Chem. 2003, 278, 16844–16851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cartaud, A.; Maoui, A.; Cartaud, J.; Stetzkowski-Marden, F. Agrin triggers the clustering of raft-associated acetylcholine receptors through actin cytoskeleton reorganization. Biol. Cell 2011, 103, 287–301. [Google Scholar] [CrossRef]
- Simon, A.M.; Hoppe, P.; Burden, S.J. Spatial restriction of AChR gene expression to subsynaptic nuclei. Development 1992, 114, 545–553. [Google Scholar] [CrossRef]
- Vale, R.D. Intracellular transport using microtubule-based motors. Annu. Rev. Cell Biol. 1987, 3, 347–378. [Google Scholar] [CrossRef]
- Kudlik, G.; Takács, T.; Radnai, L.; Kurilla, A.; Szeder, B.; Koprivanacz, K.; Merő, B.; Buday, L.; Vas, V. Advances in Understanding TKS4 and TKS5: Molecular Scaffolds Regulating Cellular Processes from Podosome and Invadopodium Formation to Differentiation and Tissue Homeostasis. Int. J. Mol. Sci. 2020, 21, 8117. [Google Scholar] [CrossRef]
- Boateng, L.R.; Huttenlocher, A. Spatiotemporal regulation of Src and its substrates at invadosomes. Eur. J. Cell Biol. 2012, 91, 878–888. [Google Scholar] [CrossRef] [Green Version]
- García, E.; E Jones, G.; Machesky, L.; Antón, I.M. WIP: WASP-interacting proteins at invadopodia and podosomes. Eur. J. Cell Biol. 2012, 91, 869–877. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pęziński, M.; Maliszewska-Olejniczak, K.; Daszczuk, P.; Mazurek, P.; Niewiadomski, P.; Rędowicz, M.J. Tks5 Regulates Synaptic Podosome Formation and Stabilization of the Postsynaptic Machinery at the Neuromuscular Junction. Int. J. Mol. Sci. 2021, 22, 12051. https://doi.org/10.3390/ijms222112051
Pęziński M, Maliszewska-Olejniczak K, Daszczuk P, Mazurek P, Niewiadomski P, Rędowicz MJ. Tks5 Regulates Synaptic Podosome Formation and Stabilization of the Postsynaptic Machinery at the Neuromuscular Junction. International Journal of Molecular Sciences. 2021; 22(21):12051. https://doi.org/10.3390/ijms222112051
Chicago/Turabian StylePęziński, Marcin, Kamila Maliszewska-Olejniczak, Patrycja Daszczuk, Paula Mazurek, Paweł Niewiadomski, and Maria Jolanta Rędowicz. 2021. "Tks5 Regulates Synaptic Podosome Formation and Stabilization of the Postsynaptic Machinery at the Neuromuscular Junction" International Journal of Molecular Sciences 22, no. 21: 12051. https://doi.org/10.3390/ijms222112051