DNA Methylation Levels in Mononuclear Leukocytes from the Mother and Her Child Are Associated with IgE Sensitization to Allergens in Early Life

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
2.1. DNA Methylation Differences in ALADDIN
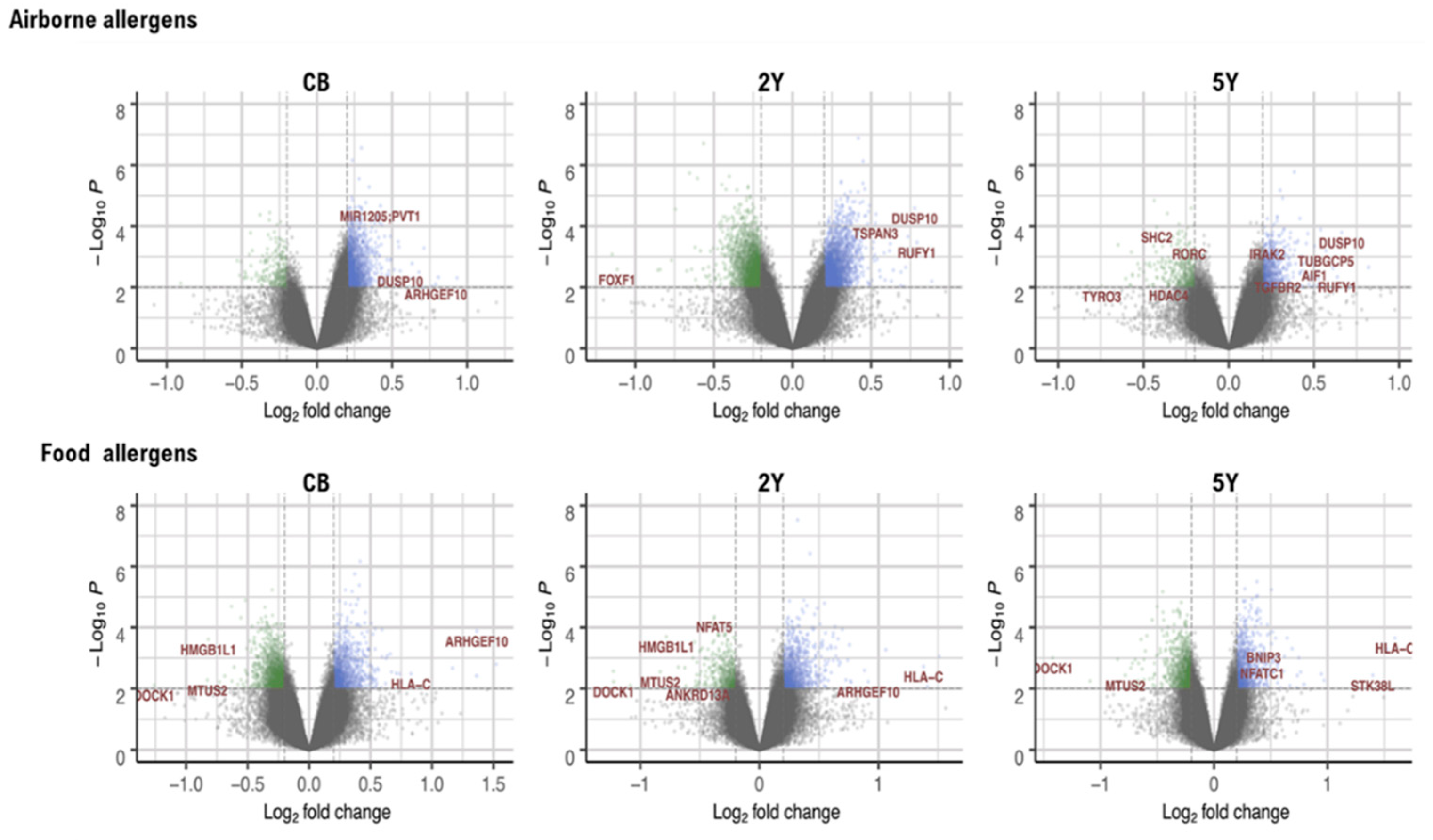
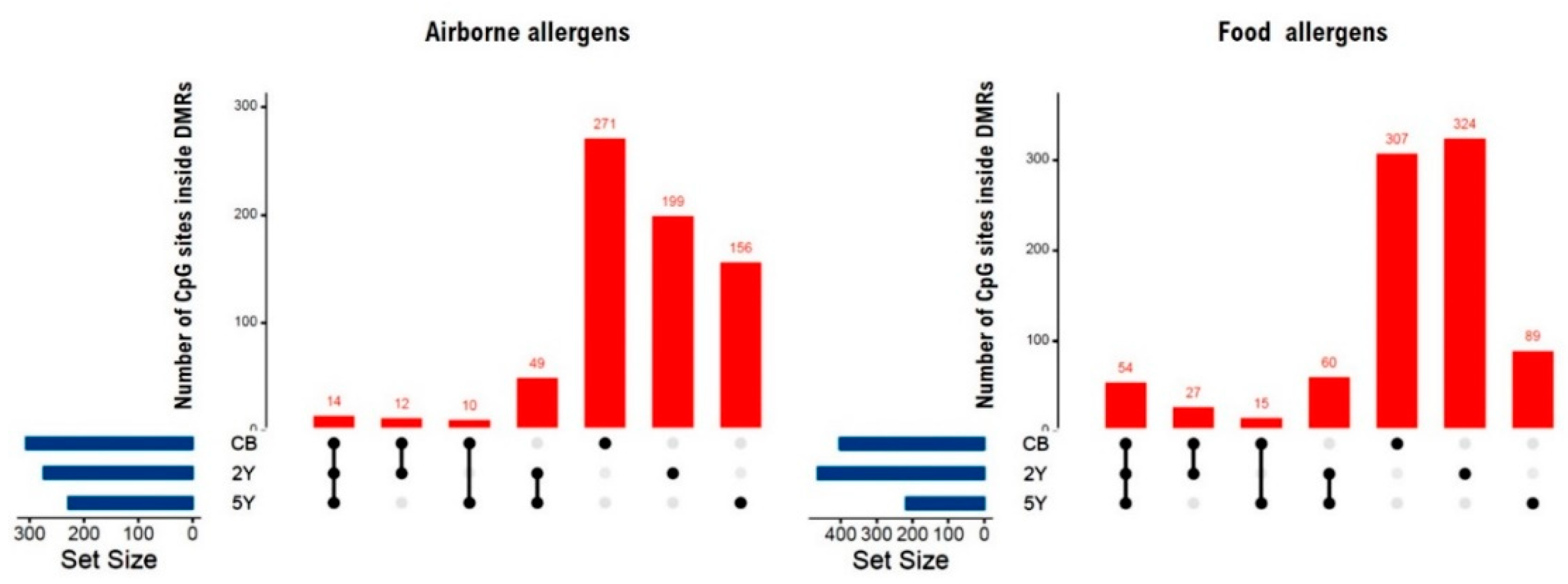
2.1.1. DMRs Associated with IgE Sensitization to Airborne Allergens at 5 Years
2.1.2. DMRs Associated with IgE Sensitization to Food Allergens at 5 Years
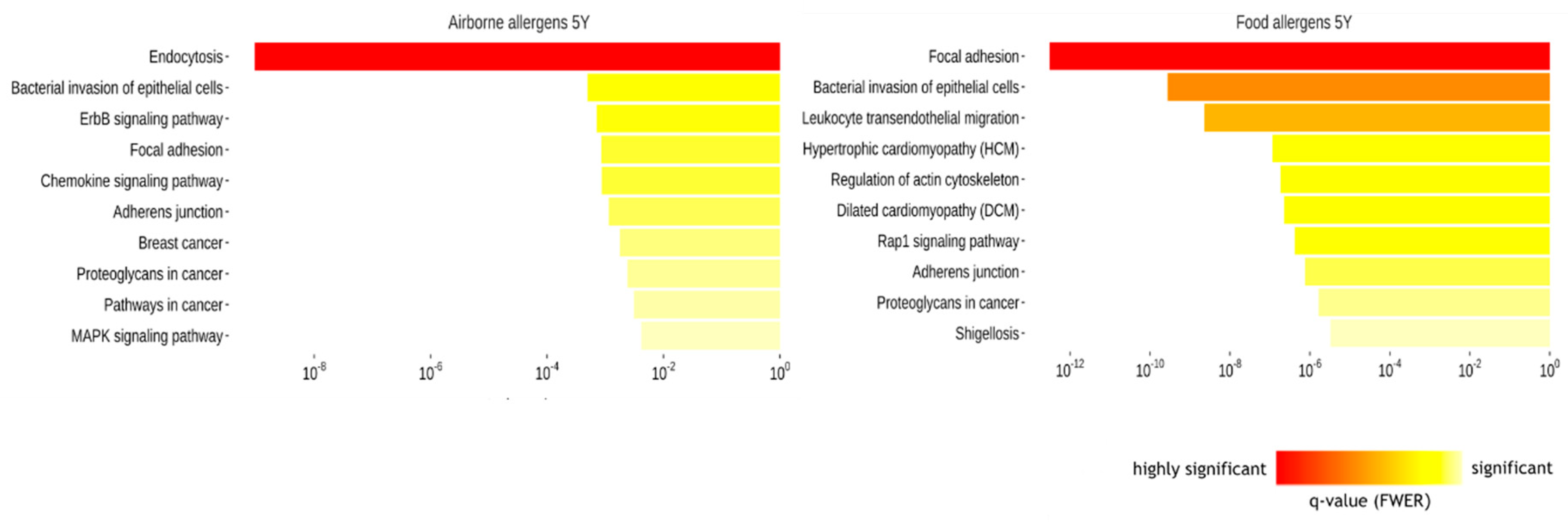
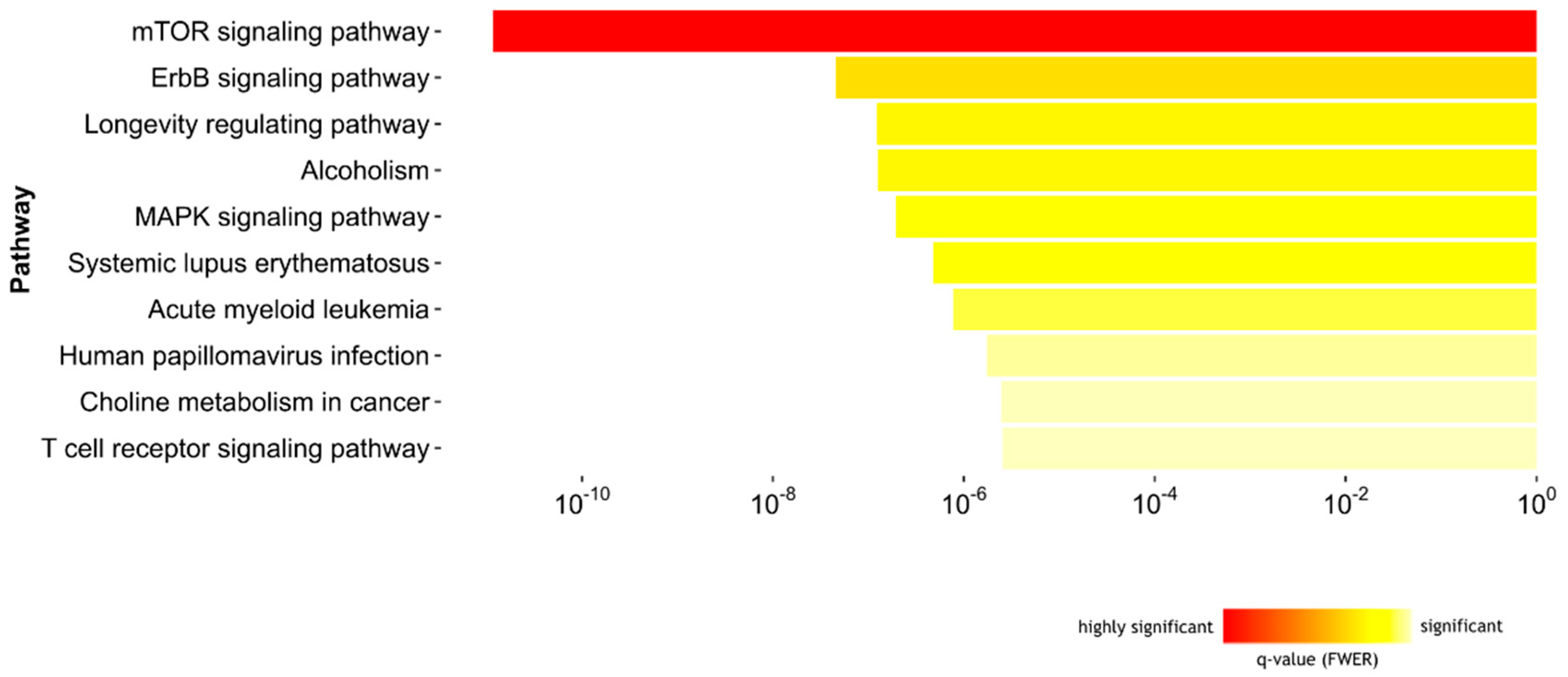
2.1.3. Biological Pathways Related to Differentially Methylated Regions and IgE Sensitization
2.2. Replication in the BAMSE Cohort
3. Discussion
4. Materials and Methods
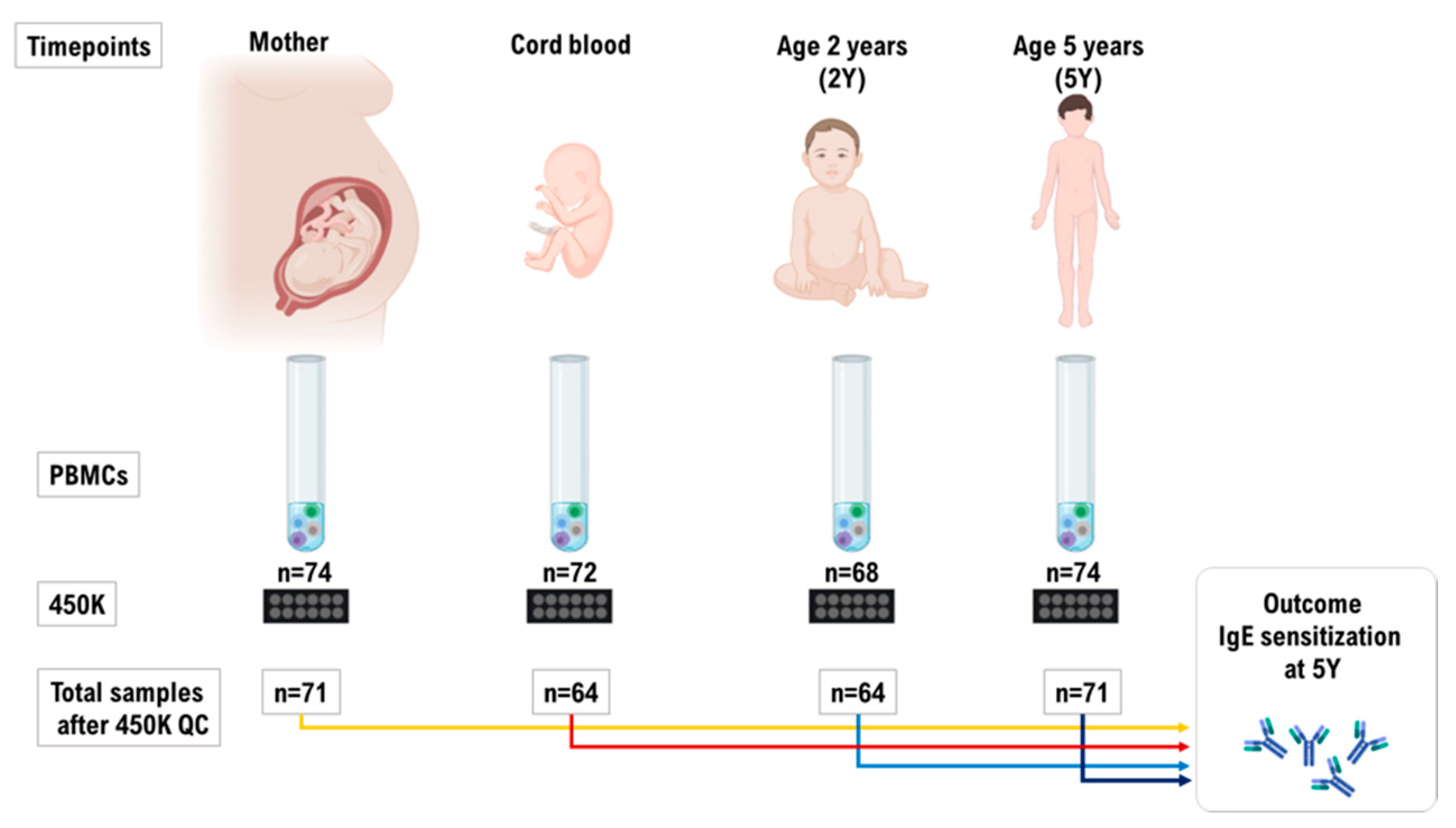
4.1. Study Population and Family Selection
4.2. Preparation of Peripheral Blood Mononuclear Cells (PBMC)
4.3. DNA Extraction and DNA Methylation Profiling
4.4. Flow Cytometry Analysis
4.5. Bioinformatics Analysis of DNA Methylation Data
4.6. Differentially Methylated Regions (DMRs)
4.7. Pathway Enrichment Analysis
4.8. Replication Analysis
4.9. Data Availability
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lambrecht, B.N.; Hammad, H. The immunology of the allergy epidemic and the hygiene hypothesis. Nat. Immunol. 2017, 18, 1076–1083. [Google Scholar] [CrossRef] [PubMed]
- Haahtela, T. A biodiversity hypothesis. Allergy 2019, 74, 1445–1456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alashkar Alhamwe, B.; Alhamdan, F.; Ruhl, A.; Potaczek, D.P.; Renz, H. The role of epigenetics in allergy and asthma development. Curr. Opin. Allergy Clin. Immunol. 2020, 20, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Shaker, M. New insights into the allergic march. Curr. Opin. Pediatr. 2014, 26, 516–520. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.; Van Meel, E.R.; Cardenas, A.; Rifas-Shiman, S.L.; Sonawane, A.R.; Glass, K.R.; Gold, D.R.; Platts-Mills, T.A.; Lin, X.; Oken, E.; et al. Epigenome-wide association study reveals methylation pathways associated with childhood allergic sensitization. Epigenetics 2019, 14, 445–466. [Google Scholar] [CrossRef]
- Cardenas, A.; Sordillo, J.E.; Rifas-Shiman, S.L.; Chung, W.; Liang, L.; Coull, B.A.; Hivert, M.F.; Lai, P.S.; Forno, E.; Celedon, J.C.; et al. The nasal methylome as a biomarker of asthma and airway inflammation in children. Nat. Commun. 2019, 10, 3095. [Google Scholar] [CrossRef] [Green Version]
- Alag, A. Machine learning approach yields epigenetic biomarkers of food allergy: A novel 13-gene signature to diagnose clinical reactivity. PLoS ONE 2019, 14, e0218253. [Google Scholar] [CrossRef] [Green Version]
- Martino, D.; Neeland, M.; Dang, T.; Cobb, J.; Ellis, J.; Barnett, A.; Tang, M.; Vuillermin, P.; Allen, K.; Saffery, R. Epigenetic dysregulation of naive CD4+ T-cell activation genes in childhood food allergy. Nat. Commun. 2018, 9, 3308. [Google Scholar] [CrossRef]
- Hoang, T.T.; Sikdar, S.; Xu, C.J.; Lee, M.K.; Cardwell, J.; Forno, E.; Imboden, M.; Jeong, A.; Madore, A.M.; Qi, C.; et al. Epigenome-Wide Association Study of DNA Methylation and Adult Asthma in the Agricultural Lung Health Study. Eur. Respir. J. 2020, 56. [Google Scholar] [CrossRef]
- Song, Y.; Liu, C.; Hui, Y.; Srivastava, K.; Zhou, Z.; Chen, J.; Miller, R.L.; Finkelman, F.D.; Li, X.M. Maternal allergy increases susceptibility to offspring allergy in association with TH2-biased epigenetic alterations in a mouse model of peanut allergy. J. Allergy Clin. Immunol. 2014, 134, 1339–1345.e7. [Google Scholar] [CrossRef] [Green Version]
- Niedzwiecki, M.; Zhu, H.; Corson, L.; Grunig, G.; Factor, P.H.; Chu, S.; Jiang, H.; Miller, R.L. Prenatal exposure to allergen, DNA methylation, and allergy in grandoffspring mice. Allergy 2012, 67, 904–910. [Google Scholar] [CrossRef] [Green Version]
- Stenius, F.; Swartz, J.; Lilja, G.; Borres, M.; Bottai, M.; Pershagen, G.; Scheynius, A.; Alm, J. Lifestyle factors and sensitization in children-the ALADDIN birth cohort. Allergy 2011, 66, 1330–1338. [Google Scholar] [CrossRef] [PubMed]
- Alm, J.S.; Swartz, J.; Lilja, G.; Scheynius, A.; Pershagen, G. Atopy in children of families with an anthroposophic lifestyle. Lancet 1999, 353, 1485–1488. [Google Scholar] [CrossRef] [Green Version]
- Floistrup, H.; Swartz, J.; Bergstrom, A.; Alm, J.S.; Scheynius, A.; van Hage, M.; Waser, M.; Braun-Fahrlander, C.; Schram-Bijkerk, D.; Huber, M.; et al. Allergic disease and sensitization in Steiner school children. J. Allergy Clin. Immunol. 2006, 117, 59–66. [Google Scholar] [CrossRef]
- Kanehisa, M.; Araki, M.; Goto, S.; Hattori, M.; Hirakawa, M.; Itoh, M.; Katayama, T.; Kawashima, S.; Okuda, S.; Tokimatsu, T.; et al. KEGG for linking genomes to life and the environment. Nucleic Acids Res. 2008, 36, D480–D484. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.; Cardenas, A.; Rifas-Shiman, S.L.; Hivert, M.F.; Gold, D.R.; Platts-Mills, T.A.; Lin, X.; Oken, E.; Baccarelli, A.A.; Litonjua, A.A.; et al. Epigenome-wide association study of total serum immunoglobulin E in children: A life course approach. Clin. Epigenetics 2018, 10, 55. [Google Scholar] [CrossRef] [PubMed]
- Reinius, L.E.; Acevedo, N.; Joerink, M.; Pershagen, G.; Dahlen, S.E.; Greco, D.; Soderhall, C.; Scheynius, A.; Kere, J. Differential DNA methylation in purified human blood cells: Implications for cell lineage and studies on disease susceptibility. PLoS ONE 2012, 7, e41361. [Google Scholar] [CrossRef] [PubMed]
- Gosney, J.A.; Wilkey, D.W.; Merchant, M.L.; Ceresa, B.P. Proteomics reveals novel protein associations with early endosomes in an epidermal growth factor-dependent manner. J. Biol. Chem. 2018, 293, 5895–5908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lang, R.; Raffi, F.A.M. Dual-Specificity Phosphatases in Immunity and Infection: An Update. Int. J. Mol. Sci 2019, 20, 2710. [Google Scholar] [CrossRef] [Green Version]
- Li, J.P.; Yang, C.Y.; Chuang, H.C.; Lan, J.L.; Chen, D.Y.; Chen, Y.M.; Wang, X.; Chen, A.J.; Belmont, J.W.; Tan, T.H. The phosphatase JKAP/DUSP22 inhibits T-cell receptor signalling and autoimmunity by inactivating Lck. Nat. Commun. 2014, 5, 3618. [Google Scholar] [CrossRef] [Green Version]
- Gaudette, B.T.; Jones, D.D.; Bortnick, A.; Argon, Y.; Allman, D. mTORC1 coordinates an immediate unfolded protein response-related transcriptome in activated B cells preceding antibody secretion. Nat. Commun. 2020, 11, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Chen, X.; Zang, A.; Li, T.; Hu, Y.; Ma, S.; Lu, M.; Yin, H.; Wang, H.; Zhang, X.; et al. TSC1/mTOR-controlled metabolic-epigenetic cross talk underpins DC control of CD8+ T-cell homeostasis. PLoS Biol. 2019, 17, e3000420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aladegbami, B.; Barron, L.; Bao, J.; Colasanti, J.; Erwin, C.R.; Warner, B.W.; Guo, J. Epithelial cell specific Raptor is required for initiation of type 2 mucosal immunity in small intestine. Sci. Rep. 2017, 7, 5580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yick, C.Y.; Zwinderman, A.H.; Kunst, P.W.; Grunberg, K.; Mauad, T.; Chowdhury, S.; Bel, E.H.; Baas, F.; Lutter, R.; Sterk, P.J. Gene expression profiling of laser microdissected airway smooth muscle tissue in asthma and atopy. Allergy 2014, 69, 1233–1240. [Google Scholar] [CrossRef] [PubMed]
- Zenarruzabeitia, O.; Vitalle, J.; Eguizabal, C.; Simhadri, V.R.; Borrego, F. The Biology and Disease Relevance of CD300a, an Inhibitory Receptor for Phosphatidylserine and Phosphatidylethanolamine. J. Immunol. 2015, 194, 5053–5060. [Google Scholar] [CrossRef] [Green Version]
- Karra, L.; Singh Gangwar, R.; Shamri, R.; Puzzovio, P.G.; Cohen-Mor, S.; Levy, B.D.; Levi-Schaffer, F. Leukocyte CD300a Contributes to the Resolution of Murine Allergic Inflammation. J. Immunol. 2018, 201, 2998–3005. [Google Scholar] [CrossRef]
- Karra, L.; Gangwar, R.S.; Puzzovio, P.G.; Fyhrquist, N.; Minai-Fleminger, Y.; Landolina, N.; Simon, H.U.; Alenius, H.; Leibovici, V.; Simon, D.; et al. CD300a expression is modulated in atopic dermatitis and could influence the inflammatory response. Allergy 2019, 74, 1377–1380. [Google Scholar] [CrossRef]
- Pathak, M.; Padghan, P.; Halder, N.; Giri, S.; Kulkarni, N.; Sonar, S.A.; Lal, G. CCR9 signaling in dendritic cells drives the differentiation of Foxp3(+) Tregs and suppresses the allergic IgE response in the gut. Eur. J. Immunol. 2020, 50, 404–417. [Google Scholar] [CrossRef]
- Lopez-Pacheco, C.; Soldevila, G.; Du Pont, G.; Hernandez-Pando, R.; Garcia-Zepeda, E.A. CCR9 Is a Key Regulator of Early Phases of Allergic Airway Inflammation. Mediat. Inflamm. 2016, 2016. [Google Scholar] [CrossRef] [Green Version]
- Castan, L.; Cheminant, M.A.; Colas, L.; Brouard, S.; Magnan, A.; Bouchaud, G. Food allergen-sensitized CCR9(+) lymphocytes enhance airways allergic inflammation in mice. Allergy 2018, 73, 1505–1514. [Google Scholar] [CrossRef]
- Martino, D.; Joo, J.E.; Sexton-Oates, A.; Dang, T.; Allen, K.; Saffery, R.; Prescott, S. Epigenome-wide association study reveals longitudinally stable DNA methylation differences in CD4+ T cells from children with IgE-mediated food allergy. Epigenetics 2014, 9, 998–1006. [Google Scholar] [CrossRef] [PubMed]
- Lluis, A.; Depner, M.; Gaugler, B.; Saas, P.; Casaca, V.I.; Raedler, D.; Michel, S.; Tost, J.; Liu, J.; Genuneit, J.; et al. Protection Against Allergy: Study in Rural Environments Study, G., Increased regulatory T-cell numbers are associated with farm milk exposure and lower atopic sensitization and asthma in childhood. J. Allergy Clin. Immunol. 2014, 133, 551–559. [Google Scholar] [CrossRef] [PubMed]
- Syed, A.; Garcia, M.A.; Lyu, S.C.; Bucayu, R.; Kohli, A.; Ishida, S.; Berglund, J.P.; Tsai, M.; Maecker, H.; O’Riordan, G.; et al. Peanut oral immunotherapy results in increased antigen-induced regulatory T-cell function and hypomethylation of forkhead box protein 3 (FOXP3). J. Allergy Clin. Immunol. 2014, 133, 500–510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, C.; Wu, H.; Lu, Q. The Epigenetics of Food Allergy. Adv. Exp. Med. Biol 2020, 1253, 141–152. [Google Scholar] [PubMed]
- Rose, S.; Laan, M.J. Why match? Investigating matched case-control study designs with causal effect estimation. Int. J. Biostat. 2009, 5. [Google Scholar] [CrossRef] [Green Version]
- Maksimovic, J.; Gordon, L.; Oshlack, A. SWAN: Subset-quantile within array normalization for illumina infinium HumanMethylation450 BeadChips. Genome Biol. 2012, 13, R44. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.A.; Lemire, M.; Choufani, S.; Butcher, D.T.; Grafodatskaya, D.; Zanke, B.W.; Gallinger, S.; Hudson, T.J.; Weksberg, R. Discovery of cross-reactive probes and polymorphic CpGs in the Illumina Infinium HumanMethylation450 microarray. Epigenetics 2013, 8, 203–209. [Google Scholar] [CrossRef] [Green Version]
- Leek, J.T.; Johnson, W.E.; Parker, H.S.; Jaffe, A.E.; Storey, J.D. The sva package for removing batch effects and other unwanted variation in high-throughput experiments. Bioinformatics 2012, 28, 882–883. [Google Scholar] [CrossRef]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. Limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef]
- Pedersen, B.S.; Schwartz, D.A.; Yang, I.V.; Kechris, K.J. Comb-p: Software for combining, analyzing, grouping and correcting spatially correlated P-values. Bioinformatics 2012, 28, 2986–2988. [Google Scholar] [CrossRef] [Green Version]
- Ogris, C.; Helleday, T.; Sonnhammer, E.L. PathwAX: A web server for network crosstalk based pathway annotation. Nucleic Acids Res. 2016, 44, W105–W109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogris, C.; Guala, D.; Helleday, T.; Sonnhammer, E.L. A novel method for crosstalk analysis of biological networks: Improving accuracy of pathway annotation. Nucleic Acids Res. 2017, 45, e8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wickman, M.; Ahlstedt, S.; Lilja, G.; van Hage Hamsten, M. Quantification of IgE antibodies simplifies the classification of allergic diseases in 4-year-old children. A report from the prospective birth cohort study--BAMSE. Pediatr. Allergy Immunol. 2003, 14, 441–447. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.-J.; Söderhäll, C.; Bustamante, M.; Baïz, N.; Gruzieva, O.; Gehring, U.; Mason, D.; Chatzi, L.; Basterrechea, M.; Llop, S.; et al. DNA methylation in childhood asthma: An epigenome-wide meta-analysis. Lancet Respir. Med. 2018, 6, 379–388. [Google Scholar] [CrossRef] [Green Version]
- Aryee, M.J.; Jaffe, A.E.; Corrada-Bravo, H.; Ladd-Acosta, C.; Feinberg, A.P.; Hansen, K.D.; Irizarry, R.A. Minfi: A flexible and comprehensive Bioconductor package for the analysis of Infinium DNA methylation microarrays. Bioinformatics 2014, 30, 1363–1369. [Google Scholar] [CrossRef] [Green Version]
- Pidsley, R.; Wong, C.C.Y.; Volta, M.; Lunnon, K.; Mill, J.; Schalkwyk, L.C. A data-driven approach to preprocessing Illumina 450K methylation array data. BMC Genomics 2013, 14, 293. [Google Scholar] [CrossRef] [Green Version]
- Houseman, E.A.; Accomando, W.P.; Koestler, D.C.; Christensen, B.C.; Marsit, C.J.; Nelson, H.H.; Wiencke, J.K.; Kelsey, K.T. DNA methylation arrays as surrogate measures of cell mixture distribution. BMC Bioinform. 2012, 13, 86. [Google Scholar] [CrossRef] [Green Version]
- Willer, C.J.; Li, Y.; Abecasis, G.R. METAL: Fast and efficient meta-analysis of genomewide association scans. Bioinformatics 2010, 26, 2190–2191. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Non-Sensitized to Airborne Allergens at Age 5 Years (n = 36) | IgE Sensitized to Airborne Allergens 1 at Age 5 Years (n = 38) | p Value 3 |
|---|---|---|---|
| Child male gender, n (%) | 20 (55.6) | 20 (52.6) | 0.80 |
| IgE sensitized to food allergens 2 at 5Y, n (%) | 0 (0) | 17 (44.7) | <0.0001 |
| Lifestyle, n (%) | |||
| Anthroposophic | 9 (25) | 9 (23.7) | 0.95 |
| Partly anthroposophic | 13 (36.1) | 15 (39.5) | |
| Non-anthroposophic | 14 (38.9) | 14 (36.8) | |
| Maternal age in years, mean (s.d.) | 31.4 ± 4.2 | 31.1 ± 5.5 | 0.82 |
| Mother sensitized to airborne allergens, n (%) | 9 (25) | 12 (31.6) | 0.53 |
| Father sensitized to airborne allergens, n (%) | 17 (48.6) | 21 (58.3) | 0.41 |
| Mother smoked during pregnancy, n (%) | 0 | 5 (13.5) | 0.054 |
| Father smoked during pregnancy, n (%) | 6 (16.7) | 10 (27) | 0.28 |
| Chr | Start | End | Width bp | Number of CpGs | Gene Name | Location | T | p-Value | Sidak p-Value |
|---|---|---|---|---|---|---|---|---|---|
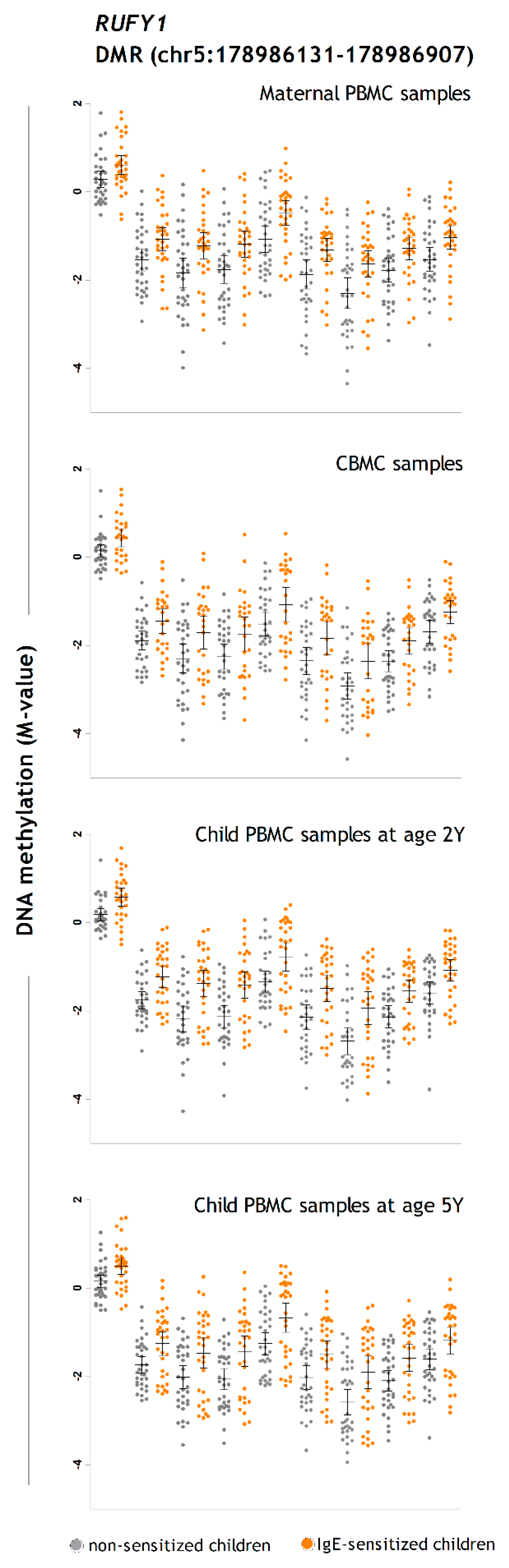
| chr5 | 178,986,131 | 178,986,907 | 777 | 9 | RUFY1 | Promoter | 4.41 | 1.6 × 10−10 | 9.1 × 10−8 |
| chr19 | 15,121,204 | 15,121,597 | 394 | 9 | CCDC105 | Promoter | 1.22 | 6.4 × 10−9 | 0.000007 |
| chr1 | 115,397,374 | 11,5397,617 | 244 | 5 | SYCP1 | Promoter | 0.99 | 7.7 × 10−9 | 0.00001 |
| chr6 | 291,882 | 292,597 | 716 | 7 | DUSP22 | Promoter | −5.31 | 6.3 × 10−8 | 0.00003 |
| chr15 | 22,833,149 | 22,833,803 | 655 | 11 | TUBGCP5 | Promoter | 3.42 | 7.5 × 10−8 | 0.00005 |
| chr17 | 72,462,164 | 72,462,636 | 473 | 6 | CD300A | Promoter | −1.56 | 2.6 × 10−7 | 0.0002 |
| chr13 | 24,144,483 | 24,144,986 | 504 | 6 | TNFRSF19 | Promoter | 1.31 | 3.4 × 10−7 | 0.0002 |
| chr6 | 31,583,458 | 31,584,224 | 767 | 5 | AIF1 | Promoter | 1.97 | 8.0 × 10−7 | 0.0004 |
| chr19 | 49,223,814 | 49,224,166 | 353 | 5 | RASIP1 | 3′ UTR | −1.02 | 5.7 × 10−7 | 0.0007 |
| chr1 | 2,120,985 | 2,121,522 | 538 | 5 | FAAP20 | 3′ UTR | −1.94 | 9.4 × 10−7 | 0.0007 |
| Chr | Start | End | Width bp | Number of CpGs | Gene Name | Location | T | p-Value | Sidak p-Value |
|---|---|---|---|---|---|---|---|---|---|
| chr17 | 6,796,745 | 6,797,772 | 1028 | 9 | ALOX12P2 | Promoter | −2.54 | 1.1 × 10−18 | 4.7 × 10−16 |
| chr14 | 105,944,604 | 105,945,700 | 1097 | 7 | CRIP2 | 3′ UTR | −2.04 | 2.7 × 10−11 | 1.1 × 10−8 |
| chr5 | 178,986,131 | 178,986,907 | 777 | 9 | RUFY1 | Promoter | 4.32 | 1.0 × 10−8 | 0.000005 |
| chr1 | 153,599,479 | 153,600,157 | 679 | 8 | S100A13 | Promoter | −3.21 | 1.0 × 10−8 | 0.000007 |
| chr6 | 28,911,468 | 28,912,167 | 700 | 12 | LINC01556 * | Non-coding Exon | −3.23 | 4.0 × 10−8 | 0.000025 |
| chr19 | 50,249,464 | 50,249,928 | 465 | 6 | TSKS * | intron | −1.52 | 4.3 × 10−8 | 0.000040 |
| chr11 | 43,290,958 | 43,291,211 | 254 | 5 | HNRNPKP3 | Promoter | 1.24 | 2.7 × 10−7 | 0.0004 |
| chr22 | 30,901,249 | 30,901,648 | 400 | 4 | SEC14L4 | Promoter | −0.95 | 0.000003 | 0.0032 |
| chr10 | 77,164,987 | 77,165,751 | 765 | 7 | ZNF503-AS2 | Intron | −1.30 | 0.000007 | 0.0040 |
| chr20 | 36,148,604 | 36,149,272 | 669 | 30 | NNAT | Promoter | −3.71 | 0.000007 | 0.0050 |
| Chr | Start | End | Width bp | Number of CpGs | Gene Name | Location | T | Z p-Value | Z Sidak p-Value |
|---|---|---|---|---|---|---|---|---|---|
| DMRs detected with increased DNA methylation levels in children sensitized to food allergens | |||||||||
| chr10 | 135,278,717 | 135,279,148 | 432 | 5 | SCART1 | Promoter | 1.63 | 1.4 × 10−10 | 1.4 × 10−7 |
| chr19 | 37,825,307 | 37,825,680 | 374 | 7 | HKR1 | Promoter | 2.40 | 1.6 × 10−8 | 0.000019 |
| chr1 | 1,108,820 | 1,109,984 | 1165 | 8 | TTLL10 | Promoter | 2.65 | 7.1 × 10−8 | 0.000027 |
| chr4 | 74,847,646 | 74,847,830 | 185 | 7 | PF4 | Promoter | 2.82 | 1.4 × 10−8 | 0.00003 |
| chr13 | 88,328,009 | 88,328,295 | 287 | 4 | SLITRK5 | Exon | 1.67 | 9.3 × 10−8 | 0.0001 |
| chr13 | 26,586,254 | 26,587,012 | 759 | 7 | ATP8A2 | Exon | 1.91 | 3.6 × 10−7 | 0.0002 |
| chr8 | 70,980,488 | 70,981,069 | 582 | 3 | PRDM14 | Exon | 0.54 | 2.7 × 10−7 | 0.0002 |
| chr16 | 86,766,712 | 86,768,118 | 1407 | 6 | LINC02188 * | Intergenic | 1.59 | 1.1 × 10−6 | 0.0003 |
| chr10 | 99,338,056 | 99,338,241 | 186 | 4 | ANKRD2 | Exon | 1.18 | 2.1 × 10−7 | 0.0005 |
| chr1 | 3,774,827 | 3,775,207 | 381 | 6 | DFFB | Promoter | 1.34 | 7.0 × 10−7 | 0.0008 |
| chr6 | 39,760,607 | 39,761,596 | 990 | 6 | DAAM2 | Promoter | 0.92 | 0.000002 | 0.0010 |
| DMRs detected with reduced DNA methylation levels in children sensitized to food allergens | |||||||||
| chr10 | 128,810,484 | 128,810,905 | 422 | 3 | DOCK1 | Exon | −2.54 | 1.2 × 10−6 | 0.0013 |
| chr17 | 72,462,417 | 72,463,081 | 665 | 6 | CD300A | Promoter | −2.28 | 5.8 × 10−6 | 0.0038 |
| chr17 | 75,315,486 | 75,315,668 | 183 | 6 | SEPT9 | Promoter | −1.70 | 1.2 × 10−6 | 0.0028 |
| chr15 | 45,670,865 | 45,671,196 | 332 | 10 | GATM | Promoter | −1.69 | 1.2 × 10−5 | 0.016 |
| chr11 | 1,463,541 | 1,463,663 | 123 | 4 | BRSK2 | Promoter | −1.58 | 1.0 × 10−5 | 0.038 |
| chr17 | 7,591,564 | 7,591,948 | 385 | 9 | WRAP53 | Promoter | −1.27 | 1.4 × 10−5 | 0.016 |
| chr19 | 49,223,814 | 49,224,166 | 353 | 5 | RASIP1 | 3′ UTR | −1.13 | 4.1 × 10−5 | 0.049 |
| chr3 | 149,469,835 | 149,470,420 | 586 | 7 | COMMD2 | Promoter | −1.12 | 3.6 × 10−5 | 0.026 |
| chr7 | 157,809,235 | 157,809,597 | 363 | 5 | PTPRN2 | Intron | −1.01 | 3.6 × 10−5 | 0.042 |
| chr2 | 179,387,853 | 179,388,065 | 213 | 3 | TTN-AS1 MIR548N | Promoter | −0.7 | 7.3 × 10−7 | 0.0015 |
| Gene Symbol | Gene Name | Locus | Function | DMR for Airborne Sensitization | DMR for Food Sensitization |
|---|---|---|---|---|---|
| RUFY1 | RUN and FYVE Domain Containing 1 | 5q35.3 | Binds phospholipid vesicles containing phosphatidylinositol 3-phosphate and participates in early endosomal trafficking | M, CB, 2Y, 5Y | CB, 2Y |
| DUSP22 | Dual specificity phosphatase 22 | 6p25.3 | Activates the Janus kinase signaling pathway | 5Y | 2Y |
| TUBGCP5 | Tubulin Gamma Complex Associated Protein 5 | 15q11.2 | Microtubule binding | 5Y | . |
| CD300A | CD300a Molecule | 17q25.1 | Negatively regulates TLR signaling via Myd88 | 2Y, 5Y | 5Y |
| AIF1 | Allograft Inflammatory Factor 1 | 6p21.33 | May promote macrophage activation upon being induced by cytokines and interferon | 5Y | . |
| TTLL10 | Tubulin Tyrosine Ligase Like 10 | 1p36.33 | Unknown | CB, 2Y | CB, 2Y, 5Y |
| DOCK1 | Dedicator of cytokinesis 1 | 10q26.2 | Guanine nucleotide exchange factor involved in cytoskeletal rearrangements and focal adhesion | 5Y | 5Y |
| BRSK2 | BR Serine/Threonine Kinase 2 | 11p15.5 | Mediates phosphorylation | 2Y, 5Y | 2Y, 5Y |
| COMMD2 | COMM Domain Containing 2 | 3q25.1 | May down-regulate the activation of NFκB | . | 5Y |
| RPTOR | Regulatory associated protein of MTOR complex 1 | 17q25.3 | Negatively regulates the mTOR kinase | M | . |
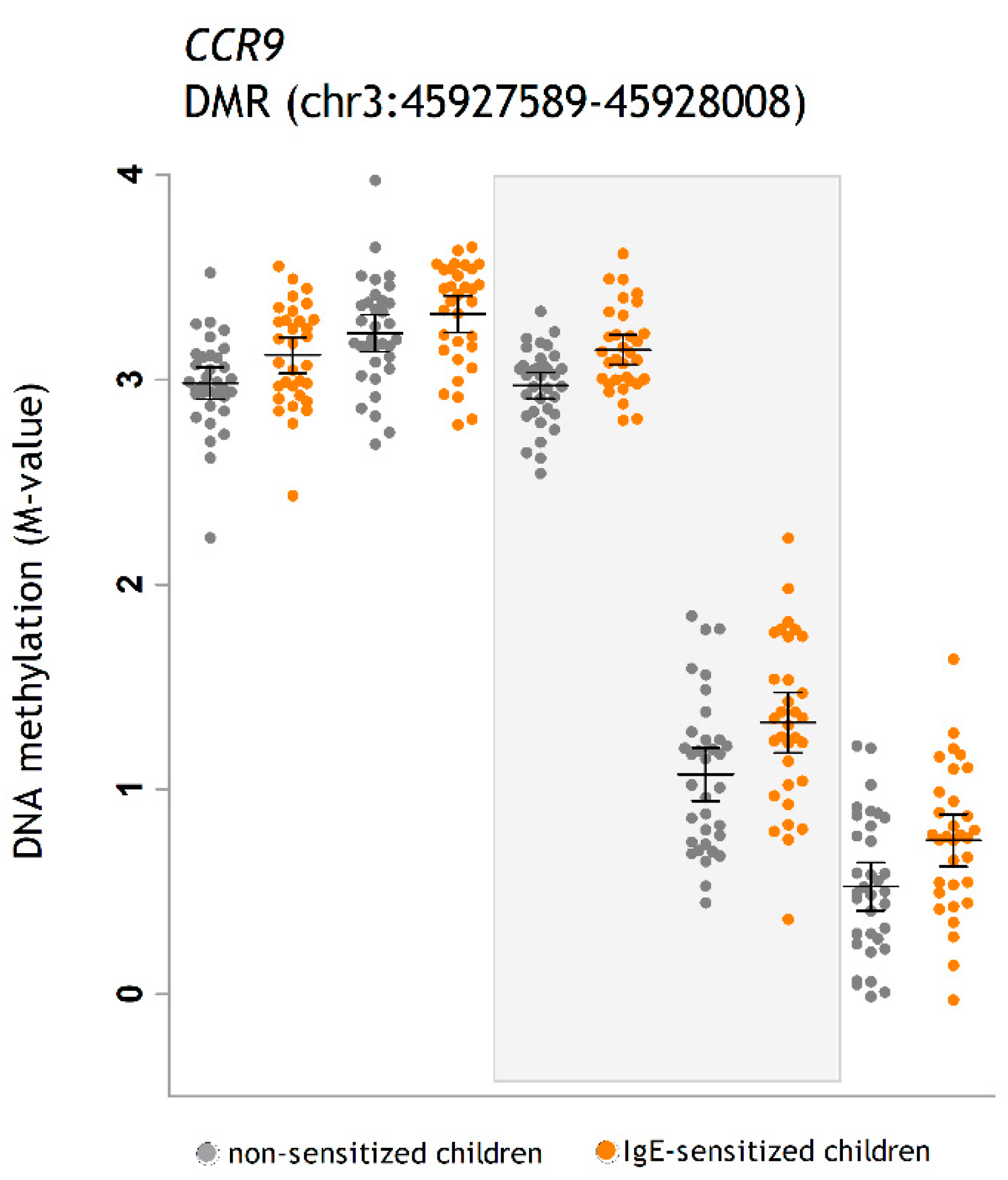
| CCR9 | C-C Motif Chemokine Receptor 9 | 3p21.31 | Chemokine receptor for CCL25. Functional specialization of immune responses in different segment of the gastrointestinal tract. | 5Y | . |
| SEPT9 | Septin 9 | 17q25.3 | Cytokinesis? May play a role in the internalization of intracellular microbial pathogens | . | 5Y |
| IGF1R | Insulin Like Growth Factor 1 Receptor | 15q26.3 | Activation of JAK/STAT signaling | CB | CB, 5Y |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Acevedo, N.; Scala, G.; Merid, S.K.; Frumento, P.; Bruhn, S.; Andersson, A.; Ogris, C.; Bottai, M.; Pershagen, G.; Koppelman, G.H.; et al. DNA Methylation Levels in Mononuclear Leukocytes from the Mother and Her Child Are Associated with IgE Sensitization to Allergens in Early Life. Int. J. Mol. Sci. 2021, 22, 801. https://doi.org/10.3390/ijms22020801
Acevedo N, Scala G, Merid SK, Frumento P, Bruhn S, Andersson A, Ogris C, Bottai M, Pershagen G, Koppelman GH, et al. DNA Methylation Levels in Mononuclear Leukocytes from the Mother and Her Child Are Associated with IgE Sensitization to Allergens in Early Life. International Journal of Molecular Sciences. 2021; 22(2):801. https://doi.org/10.3390/ijms22020801
Chicago/Turabian StyleAcevedo, Nathalie, Giovanni Scala, Simon Kebede Merid, Paolo Frumento, Sören Bruhn, Anna Andersson, Christoph Ogris, Matteo Bottai, Göran Pershagen, Gerard H. Koppelman, and et al. 2021. "DNA Methylation Levels in Mononuclear Leukocytes from the Mother and Her Child Are Associated with IgE Sensitization to Allergens in Early Life" International Journal of Molecular Sciences 22, no. 2: 801. https://doi.org/10.3390/ijms22020801