
Natural Enantiomers: Occurrence, Biogenesis and Biological Properties



1
School of Biological Science and Technology, University of Jinan, Jinan 250022, China
2
Institute for Molecular Bioscience, The University of Queensland, St. Lucia, Brisbane, QLD 4072, Australia
*
Authors to whom correspondence should be addressed.
Molecules 2022, 27(4), 1279; https://doi.org/10.3390/molecules27041279
Submission received: 4 January 2022
/
Revised: 9 February 2022
/
Accepted: 10 February 2022
/
Published: 14 February 2022
(This article belongs to the Section Natural Products Chemistry)
Abstract
:The knowledge that natural products (NPs) are potent and selective modulators of important biomacromolecules (e.g., DNA and proteins) has inspired some of the world’s most successful pharmaceuticals and agrochemicals. Notwithstanding these successes and despite a growing number of reports on naturally occurring pairs of enantiomers, this area of NP science still remains largely unexplored, consistent with the adage “If you don’t seek, you don’t find”. Statistically, a rapidly growing number of enantiomeric NPs have been reported in the last several years. The current review provides a comprehensive overview of recent records on natural enantiomers, with the aim of advancing awareness and providing a better understanding of the chemical diversity and biogenetic context, as well as the biological properties and therapeutic (drug discovery) potential, of enantiomeric NPs.
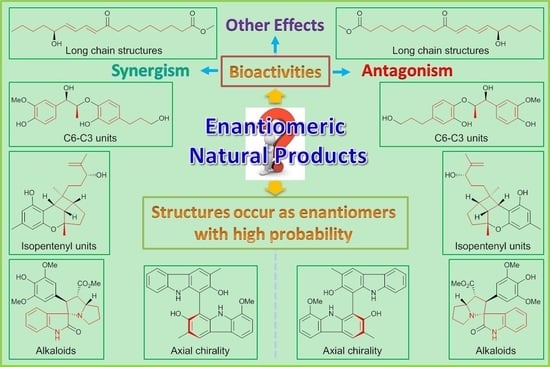
1. Introduction
Natural products (NPs) are usually regarded as small molecule organic compounds which are produced in the metabolic processes of living organisms [1]. Although studies on NPs have informed many areas of science, industry and commerce, including flavorings, perfumes, cosmeceuticals and nutraceuticals, arguably, their most important contribution to society has been as pharmaceuticals and agrochemicals [2]. For example, NPs and NP-inspired chemical entities still account for more than two thirds of all the drugs approved by Food and Drug Administration (FDA) in the USA in roughly the past four decades [2].
The vast majority of reported NPs are chiral molecules that exist in nature as single enantiomers [3]. However, as the adage goes, “Beware of exceptions to the rule”; indeed, there is increasing evidence that both enantiomers of selected NPs exist in nature. Surprisingly, NPs were generally believed to exist as single enantiomers until the 1970s, despite reports of several exceptions, probably owing to the standpoint of the famous French chemist/microbiologist Louis Pasteur, i.e., that life processes were asymmetrical [4]. Benefiting from scientific and technical advances in our understanding of NP biosynthesis, there is increasing acceptance and documentation of the occurrence of natural enantiomers. Finefield et al. reported this trend in a 2012 review, documenting the occurrence and biogenesis (where applicable) of the well-known NP enantiomers reported before 2012 [3].
During our research into bioactive NPs from medicinal plants and other sources, we have regularly encountered NP enantiomers and have documented differences in their bioactivities [5,6,7,8,9]. Surveying the scientific literature revealed the aforementioned report by Finefield et al. as the only systematic record of the occurrence of natural enantiomers [3], supported by a 2018 review by Cass et al. on the techniques for separation and absolute configuration (abs. config.) assignment of enantiomeric NPs [10]. This survey also revealed a dramatic increase in the number of publications on natural enantiomers, especially in the last few years. Against this background, the present review seeks to summarize advances in this fascinating field over the period of January 2012 to December 2019.
2. Enantiomers from Kingdom Plantae
The kingdom Plantae is an important part of nature, providing rich resources and a beautiful environment for human beings. In the field of medicine, various plants have served as the basis of traditional herbal medication to treat a variety of diseases for thousands of years. Phytochemical research on herbs has provided thousands of structural models or leads for modern drug discovery, and some NPs can even be used directly as drugs, such as taxol. NPs derived from plants have been well studied for decades, and a comprehensive system of classification has been devised. On the other hand, new NPs from kingdom Plantae are being identified all the time due to the abundance of resources. Accordingly, enantiomers produced by plants occupy the vast majority of enantiomeric NPs from natural sources.
In this section, natural enantiomers from kingdom Plantae will be classified into fourteen subcategories on the basis of their structural type, i.e., lignans, coumarins, simple phenylpropanoids, alkaloids, flavonoids, terpenoids, phloroglucinols, naphthalene and phenanthrenes, chromanes, acetophenones, diarylheptanoids, triphenylmethanes, fatty acid and miscellaneous. Where appropriate, their biogenesis and structure will also be described.
2.1. Lignans
Lignans are a common class of NPs which is widely distributed in the plant kingdom and which exhibits a broad spectrum of bioactivities including antioxidant, antitumor, anti-inflammatory, antineurodegenerative, antiviral and antimicrobial properties [11,12]. Lignans usually consist of two (sometimes three or even more) C6-C3 units (also known as phenylpropanoids). Their structural diversity arises from the different degrees of oxidation, as well as various substitution and connection patterns. Consistent with IUPAC recommendations [13], lignans are normally divided into classical lignans (only direct 8,8′-connection between the two C6-C3 units), neolignans (non-8,8′ and direct connection between the two C6-C3 units), oxyneolignans (ether oxygen linkage between the two C6-C3 units), and higher lignans (above two C6-C3 units, e.g., sesquineolignans and dineolignans). However, this classification is suggested mainly as a means of clarifying the confusing lignan nomenclature in the past, and is far from sufficient to assort the vast number of natural lignans. In general, NP chemists tend to sort lignans according to their detailed structural types, such as dibenzylbutanes, arylnaphthalenes, benzofurans, etc. [14].
Based on structural features, and for the convenience of discussion, the lignan enantiomers in the period covered by this review are classified into three subcategories: acyclic lignans, cyclic lignans and sesquineolignans. Acyclic lignans refer to those without extra rings except for the existing aromatic rings in the phenylpropanoid units, whereas cyclic lignans possess additional rings. According to the reported compound numbers, acyclic lignans are further divided into 8-4′-oxyneolignans and other acyclic lignans, while cyclic lignans will be presented as furan-incorporating lignans and other cyclic lignans. In the interests of brevity, only the structure of one enantiomer of each pair is provided; this rule applies to all structural classes in the current review.
2.1.1. Acyclic Lignans
8,4′-Oxyneolignans. 8,4′-Oxyneolignans (for structures, see Figure 1; for names, see Table S1 in Supplementary Materials) are formed via 8-O-4′ ether bonds. Also, the C-7 in these lignans is usually oxidized in a nonstereoselective manner. Then, erythro- or threo-isomers are generated, leading to the occurrence of two pairs of enantiomers.
The first reported cases in the period covered by this study were 1a/1b and 2a/2b, isolated from Paeonia lactiflora in 2015 [15]. Compounds 1 and 2 have the same constitutional structure but different relative configurations (rel. configs.). The erythro rel. config. for 1, and the threo rel. config. for 2, were determined via the J7,8 values (3.5 Hz for 1 vs. 6.8 Hz for 2), while their absolute configurations (abs. configs.) were established by the time-dependent density functional theory electronic circular dichroism (TDDFT-ECD) method. In the same year, three dinorneolignan pairs 19a/19b−21a/21b, along with a neolignan pair 25a/25b, were reported and found as scalemic mixtures in Acorus tatarinowii [16]. The determination of their abs. configs. was based on ECD data analysis and the TDDFT-ECD method. A modified Mosher’s method was also used to further confirm the abs. configs. of 19a/19b. In 2016, Gu and coworkers reported the presence of 13a/13b−16a/16b in Euphorbia sikkimensis [17], among which 13a/13b and 14a/14b and their rel. configs. had been reported in 2001 [18]. In addition, compounds 13 and 14 are diastereoisomers, as is the case for 15 and 16. In 2017, compounds 6a/6b and 7a/7b were isolated from Rubus idaeus [19]. In 2019, Song and colleagues discovered 3a/3b−5a/5b, 22a/22b and 23a/23b from Crataegus pinnatifida [20], as well as 8a/8b−12a/12b from Ailanthus altissima [21]. Among them, compounds 3 and 4 possess identical planar structures but different rel. configs., as is the case for 8 and 9.
Due to the structural flexibility of 8,4′-oxyneolignans, it has often been a challenge to correctly assign their configurations at C-7 and C-8. In order to solve this problem, three empirical rules have been developed to determine the rel. configs.: the comparison of J7,8 coupling constants [22,23], and the utilization of 13C (Δδ(C-8−C-7)) [24] and 1H (Δδ(H-9a−H-9b)) [25] NMR chemical shift differences, although each method has its limitations.
The application of the J7,8 value, first reported by Ruveda et al. in 1984 [23], is the simplest and most commonly used method (data see Table 1), but different substituents and their substitution positions could significantly impact the magnitude of J7,8, and sometimes even result in close J7,8 values for erythro and threo configurations. Additionally, the use of different deuterated solvents for NMR measurements will also influence the J7,8 value. Therefore, the configuration assignments based on this empirical rule are sometimes ambiguous or even improper due to misuse. Shi and coworkers have summarized three types of 8,4′-oxyneolignans that are suitable for the application of this rule, i.e., aglycones (J7,8 ≤ 5.0 Hz for erythro and J7,8 ≥ 7.0 Hz for threo), aglycone acetonides (J7,8 > 7.0 Hz for erythro and J7,8 < 2.0 Hz for threo) and glycoside acetates (J7,8 ≤5.3 Hz for erythro and J7,8 ≥ 6.3 Hz for threo); the NMR solvent must be CDCl3 [22]. As reflected by the previously reported data shown in Table 1, some researchers tend to apply this method without being aware of the aforementioned limitations, which could have resulted in incorrect configuration assignments and caused confusion in later studies of other NPs.
The Δδ(C-8−C-7) value was introduced to differentiate between erythro and threo 8,4′-oxyneolignans by Gan et al. [24,26], whereas only a few lignans are applied as reference compounds, and their Δδ(C-8−C-7) values also vary in different deuterated solvents, thus causing this method to lack universality. In 2019, the third method of use of Δδ(H-9a−H-9b) value was developed by Zhang and coworkers [25]. However, as with the rule of Δδ(C-8−C-7) value, lack of enough model compounds has limited its application. In summary, the rel. config. determination for 8,4′-oxyneolignans could be very complicated due to their structural flexibility and diversity, and special cautions are always suggested to avoid erroneous assignments.
Up to now, three methods, i.e., direct ECD analysis by utilizing the Cotton effect at 235 ± 5 nm, TDDFT-ECD method and modified Mosher’s method, have been used to establish the abs. configs. of 8,4′-oxyneolignans. For the first method, it is claimed that the positive Cotton effect at around 235 ± 5 nm is related to 8S-configuration, while a negative one corresponds to 8R-configuration [22]. However, different substituents on the aryl group, C-7, C-8 and C-9 would cause evident impact on the Cotton effects and the corresponding wavelengths. Caution thus should be taken when applying this rule, as improper applications have often been encountered in the literature. The TDDFT-ECD method is to theoretically predict the ECD spectra of the two possible enantiomers and then compare the calculated curves with the experimental ones. It is by far the most commonly used approach to assign abs. configs. of natural enantiomers owing to its easy operability, without the need for chemical derivatization and for constructing theoretical mechanisms to explain the observed properties [27]. Although this method is nowadays readily accessible to nonexperts, experience is still required since unexpected wrong assignments are easily made. As shown in Table 1, the abs. configs. determined by the first two methods are often inconsistent, and those assigned via the TDDFT-ECD method are usually accepted as the final determination in these reports. The third one is modified Mosher’s method that requires chemical derivatization, and its accuracy and feasibility have been proved and accepted by almost all chemists. Nonetheless, a secondary alcohol and enough amount of sample for derivatization are a must for this method, and only pure enantiomers are suitable for investigation. In addition, it is worthwhile to note that the specific optical rotation data have no straight-forward correlation with the abs. configs. of studied structures (Table 1).
Other acyclic lignans. In addition to 8,4′-oxyneolignans, many other acyclic lignan enantiomers with various connection patterns were reported in this period, as shown in Figure 2 (names see Table S2 in Supplementary Materials). Owing to the limited numbers, they have all been put together and are discussed in the current section.
Compounds 26 and 27 from Acorus tatarinowii are rare cases of naturally occurring 7,7′-oxyneolignans [28]. The threo-configurations for C-7/C-8 and C-7′/C-8′ of 26 were first determined by the large J7,8 and J7′,8′ values (both 6.5 Hz), which was further confirmed by single crystal X-ray diffraction analysis, while the abs. configs. for 26a/26b and 27a/27b were established by comparing the calculated and experimental ECD curves. The 9,9′-oxyneolignan 28 with an ester linkage was obtained as a racemic mixture from Bulbophyllum retusiusculum [29]. Compounds 29−32 from the trunk of Torreya yunnanensis are rare examples of 8,9′-neolignans and all feature a 1,3-dioxane motif by acetalization with a 2-methoxy-cinnamaldehyde [30]. Yao and coworkers reported two 8,9′-neolignans (33 and 34) and one 7,8′-neolignan (35) as racemic mixtures from Acorus tatarinowii [28]. The J7,8 values (7.6 Hz for 33 and 6.1 Hz for 34 in CD3OD) were used by the authors to determine the threo and erythro rel. configs for 33 and 34, while the rel. configs for 35 was assigned by single crystal X-ray diffraction analysis. An 8,8′-lignan (36), an 8,3′-neolignan (37) and a 7,2′-neolignan (38) were isolated from Liriodendron hybrid [31], Selaginella moellendorffii [32] and Syringa pinnatifolia [33], respectively. Sasaki et al. acquired a pair of novel 8,8′-lignan enantiomers (39a/39b) with rearranged skeleton (also known as secolignan) from Brachanthemum gobicum, with the abs. configs. being determined by comparing the ECD and specific optical rotation data with those of (−)-lucidenal [34]. The plausible biosynthetic pathway of 39 was also proposed as shown in Scheme 1, with the nonstereospecific free radical coupling being the key factor to generate enantiomers.
2.1.2. Cyclic Lignans
Furan-incorporating lignans. Furan-incorporating lignans are a class of common NPs with one or more or modified furan rings in the structures. These lignan enantiomers reported in this period mainly comprise three subtypes, namely, normal furan-type (40 and 41), benzofuran-type (42–57) and furofuran-type (58–63) (for structures, see Figure 3; for names, see Table S3 in Supplementary Materials).
Compound 40, a normal furan-type lignan enantiomer pair with 8-7′ and 7-O-8′ connections, was reported as a racemic mixture from magnolia salicifolia in 1984 [35], and was synthesized in 1992 [36]; however, its chiral separation and abs. config. determination were only realized by Lu et al. in 2018 [16]. The chiral separation and abs. config. assignment of 41, whose structure with rel. config. was reported in 1996 [37], were accomplished by Zhang and coworkers in their phytochemical investigation of Acorus tatarinowii in 2016 [38].
Benzofuran-type lignan enantiomer pairs 42−44, 45 and 46 were discovered from Jatropha integerrima in 2015 [39], Brachanthemum gobicum in 2018 [34] and Picrasma quassioides in 2018 [40], respectively. The 7,8-trans configurations for 42−46 were determined by the large J7,8 values (7.5 Hz for 42−45 in CDCl3, and 6.7 Hz for 46 in DMSO-d6) and NOE analyses, while the abs. config. assignments for these compounds were based on the reversed helicity rule [39,41]. According to this empirical rule, P-helicity of the nonaromatic ring will lead to a positive Cotton effect within the 1Lb band (around 280 nm) and M-helicity will result in a negative Cotton effect. Phytochemical investigation into the plants Rubus idaeus [42,43] and Phyllanthus glaucus [44] led to the isolation of three enantiomer pairs 47−49, and their abs. configs. were established by TDDFT-ECD method. Compounds 50−52 incorporating an α,β-unsaturated aldehyde unit were obtained from Brachanthemum gobicum [34] and Picrasma quassioides [40], with the abs. configs. also being assigned by the reversed helicity rule, where their 1Lb (α) bands red shifted to around 340 nm due to the conjugation effect from the α,β-unsaturated aldehyde group. In 2018, Huang et al. discovered compound 53 as a racemic mixture from Rubus ideaus and further resolved it into two enantiomers (53a/53b), with the abs. configs. being assigned by application of the TDDFT-ECD method [43]. In fact, compound 53 had been previously reported as an optically pure molecule from Broussonetia papyrifera in 2009, with a much smaller [α]D value [45], suggesting its potential scalemic nature. Compounds 54−57 are a group of dinorneolignans and were isolated from Rubus idaeus [42,43] and Brachanthemum gobicum [34].
In 2019, Song and colleagues obtained the trinorneolignan furolactone 58 as a racemic mixture from Rubus idaeus and resolved it into a pair of enantiomers (58a/58b), the abs. configs. of which were established by analyses of the calculated shielding tensor values and ECD data [46], while the enantiomer 58b had been reported as an optically pure molecule from Lycium chinense in 2013 [47]. The other two pairs of furolactone enantiomers 59a/59b and 60a/60b were isolated from Archidendron clypearia in 2018 [48] and Dendrobium nobile in 2016 [49], respectively. Song and coworkers reported 61a/61b, from Rubus idaeus in 2019 and assigned their abs. configs. by using the TDDFT-ECD method [47]. Compounds 62 and 63 represent another two pairs of furofuran-type lignan enantiomers isolated from Morinda citrifolia [50] and Acorus tatarinowii [16], respectively.
Other cyclic lignans. Except for the aforementioned Furan-incorporating lignan enantiomers, there are also some other cyclic lignan enantiomers with diverse ring systems as listed in Figure 4 (names see Table S4 in Supplementary Materials).
The unusual dinorneolignans 64a/64b incorporating a 1,4-dioxane motif and the arylnaphthalene-type lignans 67a/67b bearing a 2′,9′-epoxy ring were separated from Pithecellobium clypearia in 2018, with their abs. configs. being determined by the TDDFT-ECD method [51]. In 2015, Zhang et al. reported a pair of 7,8′-epoxy-8,7′-oxyneolignans (65a/65b) and a pair of oxidized arylnaphthalene-type lignans (68a/68b) from Acorus tatarinowii, and the abs. configs. of the two pairs were established by employing the TDDFT-ECD method and comparing the Cotton effect at 315 nm with those of known analogues, respectively [38]. The other pair of enantiomeric arylnaphthalenes 66a/66b were also isolated from Acorus tatarinowii, with their abs. configs. being assigned by comparing the Cotton effect at 285 nm with those of known analogues [28]. Compounds 69a/69b featuring a cyclobutane ring via 7,7′ and 8,8′ connections represent a pair of [2 + 2] cycloaddition adducts of two phenylpropanoid units and were obtained from Isatis indigotica in 2019 [52]. Isolated from Tylopilus eximius in 2012 are two pairs of enantiomers 70a/70b and 71a/71b both incorporating a cyclopentenone ring formed by 7,8′ and 9,7′ linkages [53]. The rel. config. of racemic 71 was established by X-ray crystallography, while the abs. configs. of 70a/70b and 71a/71b were confirmed by TDDFT-ECD method. In 2015, two pairs of rare spirodienone neolignans (72a/72b and 73a/73b) were reported from Cinnamomum subavenium, with the absolute structures being elucidated by X-ray crystallographic analysis and TDDFT-ECD method [54].
2.1.3. Sesquineolignans
Sesquineolignans refer to lignans bearing three phenylpropanoid units with various connection patterns. Ten pairs of enantiomeric sesquineolignans were reported in this period, and their structures are shown in Figure 5, with the names being listed in Table S5 in Supplementary Materials.
In 2015, Zhang and colleagues obtained 74−78 from Phyllanthus glaucus, with the 7,8-cis configuration of 74 being determined via the small J7,8 value (2.1 Hz) and the abs. configs. of 74a/74b being assigned by comparing ECD data with those of known analogues [44]. In addition, the 7″,8″-threo configurations of 75 and 76 and 7″,8″-erythro configurations of 77 and 78 were determined by the J7″,8″ values (6.1 Hz in CDCl3 for 75 and 6.3 Hz in Me2CO-d6 for 76; 5.1 Hz in CD3OD for 77 and 4.2 Hz in Me2CO-d6 for 78). In 2015, Yin and coworkers reported 79 and 80 from Brachanthemum gobicum and applied the reversed helicity rule to assign the abs. configs. at C-7′ and C-8′ [34], while the assignments of abs. configs. at C-7″ and C-8″ were established by Rh2(OCOCF3)4-induced ECD analysis. On the basis of the bulkiness rule for secondary alcohols, a positive Cotton effect at around 350 nm (E band) in the Rh2(OCOCF3)4-induced ECD spectrum indicated a S-configuration, while a negative Cotton effect implied a R-configuration. In 2014, Yu’s group discovered a pair of novel enantiomeric tetrahydrofuran spirodienone sesquineolignans (81a/81b) from Xanthium sibiricum and proposed coniferyl alcohol as the biosynthetic precursor (Scheme 2), and the nonstereospecific radical coupling between the two C6-C3 units was the key factor to result in enantiomers [55]. Song and coworkers reported 82 and 83 from Rubus idaeus in 2019 and assigned their abs. configs. by using the TDDFT-ECD method [47].
In summary, a large number of lignans (except lignan glycosides) have been discovered as racemic or scalemic mixtures and chirally separated in recent years, and their structural types, from simple to complex (via rearrangement), cover more than half of the known classes. It is self-evident from the aforementioned examples that enantiomerism widely occurs in the structural categories of lignans especially for 8,4′-oxyneolignans and furan-incorporating lignans. These lignan enantiomers exist as either racemic or scalemic mixtures in plants and can be relatively easily separated by commercially available chiral chromatographic columns. Therefore, it is conceivable that many examples previously reported as optically pure lignans could in fact be scalemic mixtures, and NP researchers should pay extra attention to the enantiomeric purity of lignans in their future work.
2.2. Coumarins
‘Coumarin’ is the general name of ortho-hydroxycinnamate lactones that are derived from the Shikimate biosynthetic pathway1 (for structures, see Figure 6; for names, see Table S6 in Supplementary Materials). As the core backbone of coumarins does not contain chiral factors, the generation of their enantiomersim usually comes from the chiral carbons of substituents (e.g., prenyl substitution) or axial chirality of oligomers. Coumarins are important secondary metabolites in plants and have shown various biological properties such as antitumor, anti-HIV, antimicrobial and anti-inflammatory activities [56,57].
Compounds 84a−87a are a group of angular dihydropyranocoumarins and were obtained as 3′S,4′S-configured pure enantiomers from Peucedanum japonicum in 2017 [58], while their corresponding 3′R,4′R-enantiomers (84b−87b) had been previously reported from Angelica morii in 1974 [59], Peucedanum praeruptorum in 2012 [60], Seseli gummiferum in 1971 [61] and Angelica furcijuga in 2000 [62], respectively. Another eight pairs of analogues 88−95 were found to be present as scalemic mixtures in Peucedani Radix [63] and were successfully separated into pure enantiomers for the first time. Except 88a, 89b, 92a and 92b, the others have been formerly reported as optically pure compounds [63], but the small [α]D values compared with those for the purified enantiomers suggested their scalemic natures. Tang and coworkers isolated two pairs of coumarin enantiomers (96a/96b and 97a/97b) from Toddalia asiatica and assigned the rel. and abs. configs. of 96a/96b via X-ray diffraction experiment and TDDFT-ECD method, respectively [64]. From Sapium baccatum, three coumarin enantiomer pairs incorporating one additional α-pyrone ring (98−100) were assigned the abs. configs. by comparing their specific optical rotations with those of known analogues [65]. Compounds 102 and 103 are two pairs of hybrid dimer enantiomers from Cnidium monnieri and they were also total synthesized for further biological test [66]. The most complex coumarin enantiomers so far are the oligomeric coumarin hybrids 104 and 105 bearing a spirodienone-sesquiterpene skeleton, and they were isolated from Toddalia asiatica in 2016 [67]. The only coumarin enantiomers generated by axial chirality are the prenylated coumarin dimers 101a/101b, with the abs. configs. being determined by TDDFT-ECD method [68].
2.3. Simple Phenylpropanoids
Simple phenylpropanoids are naturally occurring phenolic substances containing only one C6-C3 biosynthetic block. They often exist as racemic or scalemic mixtures in nature (for structures, see Figure 7; for names, see Table S7 in Supplementary Materials). They are also derived from the Shikimate biosynthetic pathway [1]. Many phenylpropanoids play vital roles in plant growth regulation and pathogen defense by acting as essential components of cell wall, as protectants against high light and UV radiation, and as phytoalexins against herbivores and pathogens [69]. It is generally difficult to acquire high quality crystals for X-ray diffraction analysis due to the rotary nature of the sidechains in most phenylpropanoids, so normally their rel. configs. are assigned by J values and the abs. configs. are determined on the basis of Snatzke’s rule, modified Mosher’s method or TDDFT-ECD calculation.
Qiu and coworkers reported two pairs of phenylpropanoid enantiomers, 106a/106b and 107a/107b, from the leaves of Eucommia ulmoides and assigned their rel. and abs. configs. by analysis of J7,8 values and Snatzke’s method, respectively [70]. The planar structure of 108 had already been reported in 2001 [71], but its enantiomeric nature was not revealed by Liu et al. until 2017, with the abs. configs. being determined by Snatzke’s rule [72]. Two pairs of rare chlorine-containing enantiomers (109a/109b and 110a/110b) were isolated from Acorus tatarinowii in 2017, and their rel. and abs. configs. were established by analyzing J7,8 values and employing modified Mosher’s method, respectively [73]. The enantiomer pairs 111a/111b−115a/115b were obtained from Acorus tatarinowi in 2017 by Gao’s group, among which 111b and 113b had been reported previously [74]. Compounds 115a/115b are the first cases in nature of asarone-derived phenylpropanoids with an isopropyl fragment tethered to the benzene core, and their abs. configs. were assigned by TDDFT-ECD method [74]. Song and colleagues isolated 116a/116b and 117a/117b from the fruit of Crataegus pinnatifida in 2018 and applied the TDDFT-ECD method to establish their abs. configs [74]. Compounds 118−126 with an extra phenyl group on the sidechain are also considered to be 1,2-diphenylpropane derivatives. The enantiomer pairs 118−120 were separated from Rubus idaeus in 2018 with their abs. configs. being determined via the TDDFT-ECD method [75]. Compounds 121 and 122 were first reported as racemic mixtures from Casearia grewiifolia in 2012 [76] and were resolved into two pairs of enantiomers by Qiu et al. in 2018, with the rel. and abs. configs. being determined by analyzing J7,8 values and applying TDDFT-ECD method, respectively [77]. Compounds 123a/123b−126a/126b featuring a 1,3-dioxane ring derived from condensation of diol with different aldehydes, were obtained from Crataegus pinnatifida with their abs. configs. being determined by TDDFT-ECD method [78].
2.4. Alkaloids
The term “alkaloids” traditionally describes nitrogen-containing small molecule organic compounds with basicity, although there is no unified definition. In this section, we include all nitrogen-bearing NPs in this category. Based on the structural types, natural alkaloid enantiomers from plants in the period of 2012–2019 are classified into indole alkaloids, quinoline and isoquinoline alkaloids, β-carboline and carbazole alkaloids, piperidine alkaloids, thiohydantoin alkaloids, indolizidine and quinolizidine alkaloids, and other alkaloids.
2.4.1. Indole Alkaloids
Indoles are biogenetically derived from tryptophan or tryptamine and make up one of the largest groups of alkaloid metabolites. They have attracted tremendous attention because of their therapeutic values such as anti-inflammatory, antinociceptive, antitumor, antioxidant and antimicrobial effects [79,80]. The structures of indole alkaloid enantiomers reported in this period are depicted in Figure 8 and the names summarized in Table S8 in Supplementary Materials.
Song and coworkers reported two pairs of oxindole enantiomers 127a/127b and 128a/128b from Isatis tinctoria in 2019 [81], while 128b had been previously reported in optically pure form from Isatis indigotica in 2012 [82]. In 2017, Zhang and colleagues discovered two pairs of enantiomers bearing two prenyl groups (129a/129b and 130a/130b) from Clausena lansium and assigned the rel. config. of 129b by converting it into an acetonide derivative [83]. Two pairs of novel enantiomers including the indole 3,4-dihydronaphthalen-1(2H)-one hybrids (131a/131b) and the indolizino [7,8-b]indol alkaloids (139a/139b) were found to exist as scalemic mixtures in Juglans regia [84]. In 2018, Liu et al. reported 132−135 featuring a spiropyrrolizidine oxindole skeleton from Isatis indigotica [85]. Reported from Isatis indigotica in 2019, the enantiomers 136a/136b incorporate an interesting spiro-oxindole skeleton [86]. Concurrently isolated with 136a/136b is another pairs of enantiomers 146a/146b featuring a pyrrolo[2,3-b]indolo[5,5a,6-b,a]quinazoline skeleton that had also been reported from the same species in 2012 [82]. The abs. configs. of 146a/146b were determined by using the bulkiness rule for the Rh2(OCOCF3)4-induced ECD data, wherein the E band (around 350 nm) was demonstrated to be useful for determining the abs. configs. of chiral secondary and tertiary alcohols [82]. Characterized by the presence of a dihydrothiopyran ring and a 1,2,4-thiadiazole ring in the structure, the oxindole alkaloid enantiomers 137a/137b and 138a/138b were reported from Isatis indigotica by Shi’s research group in 2018 and 2012, respectively [87,88]. The iboga-type indole alkaloid 140a was obtained as an optically pure molecule from Tabernaemontana corymbosa in 2016 [89] with the rel. config. being determined by X-ray diffraction analysis, while its enantiomer 140b was reported from Ervatamia hainanensis in 2015 [90]. Two pairs of rare indole-styrene hybrid derivatives 141a/141b and 142a/142b were isolated from Isatis indigotica [91]. The rearranged rutaecarpine-type indole alkaloid enantiomers 143a/143b from Evodia rutaecarpa incorporate an unprecedented 6/5/5/7/6 skeleton [92]. The dimeric isoechinulin-type indole alkaloid enantiomers 144a/144b and 145a/145b from Uncaria rhynchophylla feature an intriguing and complex skeleton with a symmetrical cyclobutane ring, and their rel. configs. were assigned by X-ray crystallography [93]. Except for 146a/146b, the abs. configs. of other indole enantiomers were assigned by using the TDDFT-ECD method, with the abs. config. of 138a being further confirmed by modified Mosher’s method.
It is interesting to note that suitable crystals for X-ray diffraction analysis of the enantiomeric mixtures seem relatively easy to be obtained in these reports, but the acquisition of high quality crystals of pure single enantiomers appears difficult. As above described, from simple indoles (e.g., 127) to monoterpenoid indole hybrids (e.g., 140), from single indoles (e.g., 129) to dimeric indoles (e.g., 134), from one-chiral-center examples (e.g., 131) to complex multiple-chiral-center indole dimers (e.g., 144), natural indole alkaloid enantiomers spread in a wide range of structural subtypes. Therefore, checking enantiomeric purity for this important class of NPs appears to be key in the future work.
2.4.2. Quinoline and Isoquinoline Alkaloids
Most quinoline and isoquinoline alkaloids biosynthetically originate from anthranilic acid or from indoles via rearrangement [94]. Quinoline & isoquinoline alkaloids have attracted great interest from researchers worldwide because of their wide-range biological activities, including antitumor, antiparasitic and insecticidal, antibacterial and antifungal, cardioprotective, antiviral, antiinflammatory, hepatoprotective, antioxidant, anti-asthma, antitussive, and other activities [95,96]. The structures and names of these alkaloid enantiomers in the covered stage are summarized in Figure 9 and Table S9 in Supplementary Materials, respectively.
Compounds 147a/147b, featuring a furoquinoline core hybridized with a phenylpropanoid unit via a 1,4-dioxane ring, were separated and characterized from Zanthoxylum nitidum in 2018 [97]. Three 2-quinolinone enantiomer pairs 148−150 were reported from Isatis tinctoria by Song and coworkers in 2019 [81], and in the same year, Zhang et al. discovered the same type of alkaloid enantiomers 151a/151b from the roots of Isatis indigotica [86]. The last example of quinolinone enantiomer pair is compound 152 also from I. indigotica, and it incorporates an additional anthranilic acid residue [98].
Compounds 153−158 are a series of isoquinoline enantiomers, among which 154a/154b were acquired from Corydalis hendersonii in 2016 [99] and the others were obtained from Corydalis mucronifera in 2018 [100]. Compounds 154a/154b were proposed to be derived from the condensation of a benzylisoquinoline and a succinic acid [99]. In 2016, Hua and colleagues reported from Macleaya cordata five dihydrobenzophenanthridine enantiomer pairs 159−163 and a racemate 164, among which 162 and 163 had been previously isolated in racemic form from Macleaya cordata [101] and here is the first record of their chiral separation [102]. Three same type of enantiomer pairs 165, 166 and 173 were isolated and characterized from Corydalis ambigua var. amurensis by Han and coworkers, and three racemic mixtures 167−169 were also acquired and analyzed by chiral chromatography but without further separation due to their limited amount [103]. As for structure elucidation, single-crystal X-ray diffraction analysis was applied to determine the abs. config. of 165a, followed by the abs. config. assignments for 165b and 166a/166b via comparing the ECD curves with that of 165a. In addition, Ye’s group reported a pair of berberine-type alkaloid enantiomers 170a/170b from Coptis chinensis in 2014 [104]. Sai et al. discovered from Corydalis ambigua two pairs of alkaloid dimers 171a/171b and 172a/172b, featuring a novel dimerization pattern from two different types of monomers via a C–C single bond [105]. The plausible biosynthetic pathways for 171 and 172 were also proposed as shown in Scheme 3 by the authors, and the nonstereospecific nucleophilic addition was assumed to be the key factor to generate enantiomers [105]. As with the aforementioned indole alkaloids, the assignments of abs. configs. for most quinoline and isoquinoline enantiomers have been based on the TDDFT-ECD method.
2.4.3. β-Carboline and Carbazole Alkaloids
β-Carbolines and carbazoles are among the most intriguing alkaloid groups; they derive from various sources. They have gained increasing attention due to their broad spectrum of biological activities [106,107]. Seven β-carboline (174, 178−183), three β-carboline-carbazole hybrid (175−177) and nine carbazole (184−192) enantiomer pairs have been reported in this period (for structures, see Figure 10; for names, see Table S10 in Supplementary Materials). The abs. configs. for all separated enantiomers in this section were determined by the TDDFT-ECD method unless otherwise specified.
Song and coworkers phytochemically studied the stems of Picrasma quassioides to detect four enantiomer pairs 174a/174b−177a/177b. While 174a/174b possess a β-carboline-phenylpropanoid hybrid skeleton [108], the latter three pairs represent alkaloid heterodimers of a β-carboline and a carbazole units which are linked via a C4 fragment. Alkaloids 178a/178b−180a/180b are dimeric β-carbolines obtained as trifluoroacetates from Picrasma quassioides in different years [109,110]. Compounds 181a/181b, as β-carboline-quinazoline hybrid dimers from Peganum harmala, were biogenetically produced through Mannich/Pictet–Spengler-type and intermolecular Michael addition reactions [111]. Compounds 182 and 183 from Pausinystalia yohimbe were characterized in racemic forms in 2018 without further chiral separation, and their racemic nature was further proved by X-ray diffraction analysis [112]. Interestingly, the enantiomerism of 182 results from the N-4 chiral center which is very rare in nature [112].
The enantiomerism of carbazole alkaloids comes from the axial chirality of dimers or from the chiral centers in the additional structural fragments. Four pairs of biscarbazole atropisomers (184a/184b−187a/187b) and a pair of dihydropyranocarbazole enantiomers (188a/188b) were discovered by Jiang and colleagues from Clausena dunniana, where the planar structure of 185 had been previously described from Clausena wallichii in 2011 [113]. The same authors from Jiang’s group further reported 189a/189b−192a/192b from Murraya microphylla [114,115], with the rel. config. of 189 being confirmed by X-ray crystallographic data [115].
2.4.4. Piperidine Alkaloids
Piperidine alkaloids that have one or more piperidine rings in the structures are generally believed to be biogenetically derived from lysine [1]. During the period covered by this review, fifteen pairs of piperidine enantiomers (for structures, see Figure 11; for names, see Table S11 in Supplementary Materials) have been reported.
The enantiomer pairs 193−197 were isolated from Anacyclus pyrethrum in 2018 [116]. Among them, compounds 193 and 194 possess novel dimeric piperidine backbones with 6/5/6/6 and 6/5/6 ring systems, respectively, while 195a/195b incorporate a rare cyclopentane-piperidine framework. In 2017, compounds 198−205 were obtained from Viola tianschanica. All of them bear more than one nitrogen atom and incorporate fascinating heterocyclic architectures such as the 6/5/6/5 and 6/5/5/6/5 ring systems in 198 and 201–202, respectively [117]. The abs. configs. of these alkaloid enantiomers were established by the TDDFT-ECD method. Compounds 206 and 207 were found to occur as enantiomeric pairs in Clausena lansium with only 206 being successfully resolved into pure enantiomers. The rel. and abs. configs. of 206a/206b were established by X-ray crystal data and comparing ECD and specific optical rotation data with calculated ones [118].
The biogenetic origins of these piperidines, especially of those with highly complex skeletons and multiple chiral centers like 198 and 201–202, have not been examined, and this intriguing puzzle definitely deserves further investigations.
2.4.5. Thiohydantoin Alkaloids
Naturally occurring thiohydantoin alkaloids are a rare class of NPs. Compounds 208a/208b−218a/218b (for structures, see Figure 12, names see Table S12 in Supplementary Materials), a panel of thiohydantoin derivatives of two structural groups, were initially obtained as racemic mixtures from Lepidium meyenii and further resolved into eleven pairs of enantiomers [119]. Among them, an unidentified enantiomer of 208 had been reported as a synthetic product in 2007 [120]. Although the biogenesis of these alkaloids has never been studied, they very likely belong to the imidazole class originating from histidine on the basis of their core structures [1].
2.4.6. Indolizidine and Quinolizidine Alkaloids
Both indolizidine and quinolizidine alkaloids are biogenetically originated from lysine [1], and an equal number of four pairs of indolizidine (219−222) and quinolizidine (223−226) enantiomers (for structures, see Figure 13; for names, see Table S13 in Supplementary Materials) have been reported in the period covered by this review.
Alkaloid 219 from Ficus fistulosa var. tengerensis was identified as a scalemic mixture by [α]D, ECD and X-ray crystallographic data [121], while 220 as a long-known NP re-isolated from Tylophora indica [122] was demonstrated to be a nearly racemic mixture with only a slight excess of the R-enantiomer [123]. Compounds 221a/221b, a pair of enantiomeric indolizidine alkaloid dimers from Dendrobium crepidatum, were assigned the abs. configs. by single-crystal X-ray diffraction analysis [124]. Enantiomers 222a/222b, whose structures were also confirmed by X-ray diffraction analysis to be indolizidine dimers linked via a cyclobutane ring, were obtained from the same species as 219 [121]. Zhang et al. discovered four pairs of neosecurinane-type alkaloid enantiomers 223a/223b−226a/226b of the quinolizidine class from Flueggea virosa in 2017, and it is the first time to report the enantiomerism of this interesting type of alkaloids [5]. The rel. and abs. configs. of 223a/223b−226a/226b were characterized by a variety of techniques including X-ray crystallography and ECD experiments.
2.4.7. Other Alkaloids
In addition to the aforementioned alkaloid enantiomers occurring naturally in plants, there are also many other types of alkaloid enantiomers reported in this period, as summarized in Figure 14 (names see Table S14 in Supplementary Materials).
Compounds 227a/227b−230a/230b are a panel of quinazoline enantiomer pairs obtained from Peganum harmala in 2018 [125], Isatis indigotica in 2019 [86], I. indigotica in 2016 [98] and P. harmala in 2016 [126], respectively. Biogenetically, quinazoline alkaloids have been demonstrated to be derived from anthranilic acid [1].
Qin and coworkers discovered two pairs of adenine alkaloids 231a/231b and 232a/232b from Juglans regia in 2016 [84]. Compounds 233a/233b, along with its scalemic analogue 224, were reported from Geijera parviflora, and they feature a novel heterotrimer structure incorporating a norsesquiterpenoid unit between a coumarin moiety and a proline residue [127]. Compounds 235a/235b from Juglans regia possess a benzo[b]azepine-2-carboxamide skeleton [84], while 236a/236b from Peganum harmala are amphoteric alkaloids with a four-membered N-heterocyclic ring [126].
Compounds 237−252 are amide alkaloid enantiomers with miscellaneous backbones. The simplest cases are 237a/237b bearing a thiazolidin-2-one ring and they were isolated from Isatis indigotica [87]. Zhang and coworkers discovered 238a/238b from Clausena lansium and assigned their abs. configs. by using modified Mother’s method [83], and two pairs of germacrane-type sesquiterpenoid lactams 239 and 240 were obtained from Curcuma phaeocaulis by Qiu and colleagues [128]. Compounds 241a/241b and 242a/242b are flavonoid alkaloid enantiomers reported from Scutellaria moniliorrhiza in 2018 [129], while 243a/243b represent a pair of 9,10-dihydrophenanthrene alkaloid enantiomers from Bletilla striata [130]. The enantiomer pairs 244a/244b and 245a/245b featuring a spiro[benzofuranone-benzazepine] skeleton from Juglans mandshurica [131], as well as 246a/246b incorporating a benzo[f][1,3,5]triazocine backbone from Isatis tinctoria [81], were all reported by the research team of Song and Huang. Compounds 247a/247b are a pair of enantiomers formed by an oxyneolignan and a phenethylamine units from Lycium chinense [132], while 248a/248b and 249a/249b are rearranged nor-lignan amide enantiomers featuring a unique benzo-angular triquinane skeleton from Cannabis sativa [133]. Alkaloids 250−252 were obtained as racemic mixtures from Endiandra kingiana without further chiral separation, and their racemic nature was claimed on the basis of their zero [α]D values [134].
Except for the specified ones, the abs. configs. of the alkaloid enantiomers in this section were all established by applying the TDDFT-ECD method.
2.5. Flavonoids
Flavonoids are a large family of secondary metabolites that exist widely in the plant kingdom. They exhibit a variety of bioactivities such as anti-inflammatory, antioxidant, antibacterial, antiviral, anticancer and neuroprotective effects [135]. Traditionally, flavonoids mainly refer to compounds incorporating a 2-phenylchromone core, and nowadays, this term has extended to all structures with two phenyl units linked via a C3 fragment [14]. In addition, some NPs such as xanthones and furanochromones are also included in this structural family as atypical flavonoids. Flavonoid enantiomers reported in this period are classified into three subgroups: flavones and isoflavones, chalcones and xanthones
2.5.1. Flavones and Isoflavones
In this section, the definition ‘flavones’ refers to all those incorporating the basic 2-phenylchromone backbone, including classical flavones, flavanones, flavanes, etc. The abs. configs. of these enantiomers are mostly determined by the TDDFT-ECD method unless otherwise specified. Their structures and names are shown in Figure 15 and Table S15 in Supplementary Materials, respectively.
The biflavonoid enantiomers 253a/253b were isolated from Selaginella trichoclad by Tan and coworkers in 2019 [136], with the abs. configs. being assigned by an empirical rule developed by Gaffield [137]. This rule was described as that 2S-configured flavanones and 2R,3R-configured 3-hydroxyflavanones have a positive Cotton effect at ~330 nm caused by the n→π* transition and a negative Cotton effect due to the π→π* transition at around 280−290 nm. In 2017, a pair of enantiomers hybridized from a flavonol and a coumarin via a prenyl unit (254a/254b) were isolated from Cnidium monnieri, and their constitutional structure was further confirmed by semi-synthesis through condensation of the monomeric precursors [66]. Compounds 255a/255b and 256a/256b are flavanol-phenylpropanoid adducts and were discovered from Uncaria rhynchophylla in 2017 [138,139]. Muhammad and colleagues reported a pair of 6-formylated flavanone enantiomers 257a/257b from Eugenia rigida and also semi-synthesized them for further biological studies [140]. Two pairs of flavanones coupled with a propionate residue (258a/258b and 259a/259b) were separated from the aerial parts of Abrus precatorius in 2019 by Li et al. [141]. Wang and coworkers reported three pairs of flavanone-stilbene hybrid enantiomers 260a/260b−262a/262b from Cajanus cajan, and 261a/261b feature a cyclopenta[1,2,3-de]isobenzopyran-1-one tricyclic unit with cajanolactone A being proposed as the biosynthetic precursor [142]. Two prenylated flavones 263 and 264 were isolated from Morus nigra and successfully resolved into two pairs of enantiomers in 2019 [143], while 264 had been previously reported as a racemic mixture in 2018 [144]. Compounds 263a/263b incorporate an interesting framework with a novel 7/6/6 heterocyclic ring system. Flavanes 265 and 266 were characterized as two racemic mixtures by X-ray diffraction analysis but without further separation into pure enantiomers [145]. Compounds 267a/267b−270a/270b are four pairs of diprenylated flavane enantiomers from Daphne giraldii, with the abs. configs. being determined by Rh2(OCOCF3)4 induced ECD method [146]. Compounds 271a/271b, as heterodimers derived from a flavane and a diphenylpropanoid, were isolated from Dracaena cochinchinensis in 2016 [147].
Compounds 272a/272b−278a/278b, seven pairs of enantiomeric diprenylated isoflavones with diverse ring systems, were reported from Maclura tricuspidata by Lee’s research group in 2018 [148]. The enantiomer pairs 279a/279b and 280a/280b were obtained from the stems of Pisonia umbellifera and characterized as hybrids from an isoflavone and a phenylpropanoid [149].
As can been from the above-mentioned structures, the enantiomerism of these flavones mainly comes from either the chirality of flavanone/flavane core or that of additional structural units especially prenyl group(s), or both. Meanwhile, the enantiomerism of the described isoflavones arises exclusively from the chirality of extra structural units, i.e., prenyl group(s) and phenylpropanoid fragment for the current cases.
2.5.2. Chalcones
Chalcones are open-chain flavonoids which have attracted increasing attention from researchers due to the wide range of their bioactivities, including antimicrobial, antimalarial, anticancer, anti-inflammatory, antiprotozoal, anti-HIV, antioxidant properties, etc. [150]. The chalcone enantiomers covered by this review, including monomers and dimers, are summarized in Figure 16 (Names see Table S16 in Supplementary Materials). Similarly, the determination of abs. configs. by TDDFT-ECD method will not be specified.
Compounds 281a/281b are a pair of dihydrochalcone enantiomers from Pteris ensiformis [151] with the abs. configs. being determined by Rh2(O2CCF3)4-induced ECD method. Zhang and coworkers reported three chalcone dimers formed by [2 + 2] (282a/282b and 283a/283b) and [2 + 4] (284a/284a) cycloaddition reactions from Oxytropis chiliophylla in 2018 [152]. The hydroxycinnamoylated chalcones, including four pairs of separated enantiomers 285a/285b−288a/288b and one racemate 289, were obtained from Populus balsamifera, with the abs. configs. for 285b being established by single-crystal X-ray diffraction analysis [153]. Li and coworkers reported two pairs of enantiomeric dimers formed by a dihydrochalcone and a deoxohydrochalcone (290a/290b) and by a deoxohydrochalcone and a homoisoflavane (291a/291b) from Dracaena cochinchinensis in 2016 [147]. From Horsfieldia tetratepala, compounds 292−296 were obtained as scalemic deoxohydrochalcone dimers without chiral separation [154].
2.5.3. Xanthones
Xanthones are polyphenolic compounds incorporating a common 9H-xanthen-9-one scaffold with various substituents, making them ‘privileged structures’ which are likely to bind to a variety of biological targets. They have been shown to display significant bioactivities including antimicrobial, antioxidant, cytotoxic activities, and so on [155]. Most xanthone enantiomers reported in this period are prenylated; their structures are listed in Figure 17 (names see Table S17 in Supplementary Materials).
Hua and coworkers reported a pair of diprenylated xanthone enantiomers 297a/297b with only one chiral center from Cratoxylum cochinchinense in 2019 [156]. The deoxoxanthone enantiomers 298a/298b and 299a/299b incorporating a phenylpropanoid unit were isolated from Uvaria valderramensis in 2014 [157]. Also in 2014, three pairs of prenylxanthone enantiomers (300a/300b−302a/302b) were isolated from Cratoxylum formosum, with the abs. configs. being established by X-ray crystallographic experiment [158]. As shown in Scheme 4, the generation of enantiomeric 300a/300b−302a/302b is plausibly derived from diallylxanthone through a key process of nonstereospecific Claisen rearrangement [158]. In addition to 300−302, eleven pairs of caged prenylxanthone enantiomers 303a/303b−307a/307b and 308a/308b−313a/313b were reported from Garcinia bracteata in 2018 [159] and from Garcinia propinqua in 2017 [160], respectively, with the abs. config. of 313a being determined by single-crystal X-ray diffraction analysis. The biogenetic origins of those xanthones with multiple chiral centers are indeed interesting topics that deserves further investigations.
2.6. Terpenoids
Terpenoids are probably the biggest family of NPs with diverse structures and various biological activities [3]. All terpenoids are initially assembled from the head-to-tail condensation of repeated isoprene units (C5), and according to the number of isoprene residues, terpenoids are normally classified into monoterpenoids (C10), sesquiterpenoids (C15), diterpenoids (C20), sesterterpenoids (C25) and triterpenoids (C30). Additionally, meroterpenoids are also an interesting class of terpenoid products with mixed biogenesis [3]. To the best of our knowledge, enantiomeric cases have been reported for all terpenoid subclasses except triterpenoids.
2.6.1. Sesquiterpenoids
Sesquiterpenoids are constructed from three isoprenyl fragments. Among all the terpenoid classes, they have the most diverse carbon skeletons and are probably the largest group of terpenoid NPs. Corresponding to their various structural types, natural sesquiterpenoids have also exhibited a myriad of biological properties [161], and this has been well reflected by the success of artemisinin (for malaria), the most famous sesquiterpenoid whose discovery was rewarded the Nobel prize in Physiology or Medicine in 2015. Enantiomeric sesquiterpenoids reported in this period include 16 pairs (314−329) with different backbones (for structures, see Figure 18, names see Table S18 in Supplementary Materials).
Enantiomers 314a/314b represent the first examples of 1,2-seco bisabolane-type sesquiterpenoid lactones from Artabotrys hexapetalus, with the abs. configs. being determined by employing the helicity rule to analyze the Cotton effect at around 220 nm [162]. Qiu and coworkers reported four pairs of megastigmane-type norsesquiterpenoid enantiomers 315a/315b−317a/317b and 325a/325b from Eucommia ulmoides in 2017, while the racemic mixtures of 315 and 325, along with pure enantiomers 316b and 317b, had been previously reported [163]. Compounds 318a/318b and 319a/319b are two pairs of enantiomeric germacrane type sesquiterpenes from Curcuma phaeocaulis reported in 2017 [128]. Chai and colleagues discovered 320a/320b−324a/324b with a humulane framework and 329a/329b incorporating a rare 2,2,5,9-tetramethylbicyclo[6.3.0]-undecane skeleton from Syringa pinnatifolia [164]. The abs. configs. of these compounds were established by single-crystal X-ray diffraction analysis, modified Mosher’s method and TDDFT-ECD calculation [164]. Compounds 326a/326b and 327a/327b were isolated from Commiphora myrrha [165] and Daphne genkwa [166], respectively. The guaianolide sesquiterpenoid 328 was reported as a racemic mixture as indicated by X-ray crystallography from Kadsura interior in 2013 [167].
2.6.2. Diterpenoids
Diterpenoids are biosynthesized from the head-to-tail condensation of four isoprene (C20) units, and have the second largest number of carbon backbones in the terpenoid family. Like sesquiterpenoids, they are also well-known in the NP community for their diverse bioactivities particularly antitumor effects with therapeutic values [168]. As is well known, the most notable diterpenoid is taxol, which has been used as a cancer treatment for over three decades. Nine pairs of enantiomeric diterpenoids 330a/330b−338a/338b (for structures, see Figure 19, names see Table S19 in Supplementary Materials) with different structural skeletons were recorded in the study period.
Yue and coworkers phytochemically investigated Croton mangelong to afford two pairs of macrocyclic diterpenoid enantiomers 330a/330b and 331a/331b featuring a bicyclo[9.3.1]pentadecane core and a rare bridgehead double bond, with the abs. config. for 330b being determined by single-crystal X-ray diffraction analysis [169]. Compounds 332a/332b are bis-seco-abietane diterpenoids from Cryptomeria fortune and were asymmetrically synthesized through a readily made intermediate orthoquinone from sugiol [170]. Compounds 333a/333b are a pair of norditerpenoid enantiomers from the roots of Salvia miltiorrhiza [171]. Compounds 334a/334b, rearranged abietane-type diterpeniods featuring a 5/6/6 tricyclic architecture with the five-membered ring formed via C-2–C-11 single bond, were isolated and characterized from Salvia prionitis in 2015 [171]. Compounds 335a/335b are a pair of diterpenoid enantiomers with a highly oxygenated novel backbone obtained from Swertia leducii in 2014, with the rel. config. being determined by X-ray diffraction analysis and the abs. configs. by TDDFT-ECD method [172]. In the same year, compounds 336a/336b were isolated from Paeonia veitchii [173], and they incorporate an aromatized norditerpenoid skeleton, with the rel. config. being confirmed by X-ray crystallography. Compounds 337a/337b and 338a/338b, two pairs of norditerpenoid enantiomers with unusual 5,5-spiroketal core, were obtained from Hypericum japonicum in 2016, with the abs. configs. being assigned by a combination of TDDFT-ECD calculation, modified Mosher’s method and quantum chemical predictions (QCP) of 13C NMR data [174].
2.6.3. Meroterpenoids
The term meroterpenoid was first proposed by Cornforth in 1968 to describe NPs of mixed biosynthetic origin which are partially derived from terpenoids [175]. Enantiomeric meroterpenoids reported in the covered period are exclusively formed by the condensation of phenolic compounds with a monoterpenoid or a sesquiterpenoid via at least one ether bond, and the chirality generating enantiomerism all exists in the terpenoid part except for 340. There are 27 pairs of enantiomeric meroterpenoids reported in this period (for structures, see Figure 20; for names, see Table S20).
Compounds 339a/339b−346a/346b are eight pairs of enantiomeric meromonoterpenoids with diverse heterocyclic frameworks from Rhododendron nyingchiense [176]. Among them, 339a/339b possess a rare 6/7/5/5 heterocyclic ring system, while 340a/340b incorporate a 6/6/5 tricyclic backbone with an extra oxirane ring coupled to the quinone motif. The assignments of abs. configs. for 339a and 341a were based on X-ray crystallographic experiment, while those for the others were via TDDFT-ECD method. Four enantiomeric meromonoterpenoid pairs 347a/347b−350a/350b, along with three merosesquiterpenoid pairs 351a/351b−352a/352b, were isolated and identified from Rhododendron capitatum by Hou and coworkers [177,178]. Of these interesting molecules, compounds 348 and 349 bear unprecedented 6/6/6/5 and 6/6/5/5 ring systems, respectively, while 350 and 351−333 possess unique 6/6/6/4 and 6/6/5/4 heterocyclic architectures, respectively. In addition, the abs. configs. of 350−353 pairs were confirmed by X-ray diffraction and ECD analyses. Compounds 354a/354b−360a/360b, seven pairs of bibenzyl-based meroterpenoid enantiomers, were obtained from the Chinese liverwort Radula sumatrana in 2017 by Lou’s group [179]. Compounds 361a/361b−365a/365b are five pairs of magnolol-derived lignan-monoterpenoid hybrid enantiomers that have been isolated from Magnolia officinalis in 2019 [180].
2.7. Phloroglucinols
Phloroglucinol derivatives represent a unique class of NPs featuring one or more intact or modified phloroglucinol units, with alkylation and acylation as the common structural modifications [181]. The chirality of them arises usually from their prenyl/terpenyl substituents and/or from the dearomatization of phloroglucinol core. In most cases, they can also be classified into the ‘meroterpenoid’ group, but here we describe them separately owing to the considerable number of reports and their popularity among NP workers in recent years. Phloroglucinal enantiomers reported in this period all incorporate one or more acyl groups including acetyl, isobutyryl, benzoyl, cinnamoyl and dihydrocinnamoyl, and their structures are shown in Figure 21 (names see Table S21 in Supplementary Materials). Wherever the abs. configs. are determined by the TDDFT-ECD method, it will be not specifically mentioned in this section.
Ye and coworkers reported a pair of enantiomeric isobutyrylated phloroglucinol dimers 366a/366b from Myrtus communis in 2019 and also completed their total synthesis in the same year [182]. Laphookhieo and colleagues discovered the benzoylated phloroglucinol enantiomers 367a/367b from Cratoxylum sumatranum ssp. Neriifolium and assigned their abs. configs. by comparing the specific optical rotations with those of known analogues [183]. Compounds 368a/368b−371a/371b, as phloroglucinol-monoterpenoid hybrids, were reported from Hypericum japonicum in 2016, with the abs. configs. of 370b and 371b being determined by single-crystal X-ray diffraction analysis [184]. Compounds 368a/368b and 369a/369b incorporate interesting pyrano[3,2-b]pyran and 2-oxabicyclo[3.3.1]nonane skeletons, respectively, while 370a/370b possess a benzo[b]cyclopenta[e]oxepine ring system. Laphookhieo and coworkers also obtained acetylated (372a/372b) and cinnamoylated (373a/373b) phloroglucinol enantiomers from Mallotus philippensis in 2019 and established their abs. configs. on the basis of X-ray crystallographic studies [185]. Hans et al. re-acquired myrtucommulone A (374) from Myrtus communis in 2015 and proved that, by converting it into separable derivatives, 374 consisted of the racemate and the meso form in a ca. 1:1 ratio [186]. Compounds 375a/375b−377a/377b, dihydrocinnamoylated and rearranged phloroglucinol dimers, were isolated from Xanthostemon chrysanthus in 2019 [187]. Compounds 375a/375b feature an bis-phenylpropanoyl-benzo[b]cyclopent[e]oxepine tricyclic backbone and 376a/376b and 377a/377b represent the first examples of 1-(cyclopentylmethyl)-3-(3-phenylpropanoyl)benzene scaffold [187]. Compounds 378a/378b and 379a/379b, cinnamoylated phloroglucinol dimers from Cleistocalyx operculatus, possess a polycyclic skeleton with a highly functionalized dihydropyrano[3,2-d]xanthene tetracyclic core, and the enantiotropy could be derived from the nonstereoselective hetero-Diels-Alder [4 + 2] cycloaddition as shown in the proposed plausible biosynthetic pathway (Scheme 5) [188]. The benzoylated phloroglucinol 380a was isolated from Triadenum japonicum in 2015 [189] and identified as the enantiomer of (+)-nemorosonol (380b) previously reported from Clusia nemorosa [190]. Compounds 381a/381b are a pair of digeranylated phloroglucinol enantiomers from Garcinia multiflora and feature a caged tetracyclo[5.4.1.11,5.09,13]tridecane skeleton, with the generation of enantiomerism being likely from the intramolecular Diels-Alder [4 + 2] cycloaddition on different sides as shown in Scheme 6 [191]. Compounds 382a/382b−385a/385b, four similar type of enantiomer pairs as 381, were obtained from Garcinia multiflora, and 382a/382b are characterized by the coupling of two novel caged fragments, i.e., 2,11-dioxatricyclo[4.4.1.03,9]undecane and tricyclo[4.3.1.03,7]decane, with the rel. config. being determined by X-ray diffraction analysis [192].
2.8. Naphthalenes and Phenanthrenes
The enantiomerism of naphthalene and phenanthrene derivatives is generally attributable to chiral centers and, in many cases, axial chirality. The structures of the naphthalene and phenanthrene enantiomers reported in this period are displayed in Figure 22 (names see Table S22 in Supplementary Materials). The abs. configs. of most of these enantiomers were determined by the TDDFT-ECD method, unless otherwise specified.
2.8.1. Naphthalenes
Compounds 386−397 are 12 pairs of naphthalene enantiomers reported in this period. Among them, 386−391 were isolated from the roots of Morinda officinalis var. officinalis and identified as prenylated methyl 2-naphthoates by the authors’ team, with the rel. configs. of 386 and 388 being confirmed by X-ray diffraction analysis and 13C NMR calculation, respectively [8]. Compounds 392−394, three pairs of 3,4-dihydro-4-naphthyl-naphthalen-1(2H)-one enantiomers, were obtained from Juglans regia in 2019 [193]. Compounds 395a/395b from Rubia oncotricha were characterized by Tan and coworkers as novel naphthoquinone dimers with an unprecedented spiro[4.5] carbon core [194]. Another pair of dimeric naphthoquinone enantiomers 396a/396b were also reported by the same research team from Rubia alata in 2014, with the rel. config. being corroborated by X-ray crystallographic experiment [195]. Compounds 397a/397b are simple tetrahydronaphthoquinone enantiomers reported from Eremurus altaicus in 2015 [196].
2.8.2. Phenanthrenes
Phenanthrene enantiomers (398a/398b−408a/408b) in this period have been solely reported from Bletilla striata by Li’s and Hou’s research teams in 2019 [130], and they can be divided into three groups, namely, phenanthrene monomer (405), phenanthrene dimers (404 and 406−408) and phenanthrene-phenylpropanoid hybrids (398−403). The enantiomerism of these compounds has been generated from the axial chirality of phenanthrene moiety and/or from the chiral centers of phenylpropanoid unit. Notably, compounds 404−408 with only axial chirality were able to be separated into five pairs of enantiomers. Axial chirality, although well known to organic chemists, has often been overlooked by NP researchers, owing to its rare presence in natural molecules. Therefore, the enantiomeric purity of NPs with axial chirality is strongly recommended to be checked no matter they are new or known.
2.9. Chromanes
Chromane derivatives are a class of NPs having a chromane core or a modified one (e.g., chromone, chromanone) in their structures. Enantiomeric chromane derivatives reported in this period are listed in Figure 23 (names see Table S23 in Supplementary Materials). Compounds 409a/409b−415a/415b had been studied previously in many occasions as pure enantiomers, scalemic mixtures or racemates, and as natural molecules, biotransformation products or synthetic intermediates, but none of these reports had paid attention to the enantiomerism of this group of structures. They were separated from the flower buds of Tussilago farfara in the authors’ lab in 2018, with the abs. configs. being determined by chemical method as well as TDDFT-ECD calculation and ECD comparison [7]. Compounds 416a/416b were proposed to be a pair of norbisabolane sesquiterpenoid enantiomers yet incorporating a chromone core and were obtained from Curcuma longa in 2019 [197]. Compounds 417a/417b−421a/421b are five pairs of prenylated chromone enantiomers isolated from Harrisonia perforate in 2014, whereas only the abs. configs. of 417a/417b were assigned by Mosher’s method [198]. From the same plant, 422a/422b were reported as a pair of enantiomeric molecules by Yuan et al. in 2017 [199].
2.10. Acetophenones
Acetophenones are a rare class of NPs bearing normal or rearranged acetophenone units in their structures. To date, ten pairs of acetophenone enantiomers have been reported, and their structures are depicted in Figure 24 and names listed in Table S24 in Supplementary Materials. Kong and coworkers reported four pairs of diprenylated and rearranged acetophenone enantiomers 423a/423b−426a/426b from the leaves of Melicope ptelefolia in 2019 and assigned their abs. configs. by a combination of modified Mosher’s and TDDFT-ECD methods [200], while 423a/423b and 424 in racemic form had been previously isolated from Evodia lepta by Tang et al. in 2018 [201]. Also identified as rearranged acetophenones, compounds 427a/427b coupled with a phenylpropanoid fragment were isolated from Xanthostemon chrysanthus in 2019 [187]. Compounds 428a/428b are prenylated hydroacetophenone enantiomers obtained from Melicope viticina in 2019 [202], while 429a/429b and 430a/430b with intact acetophenone unit were discovered from Eupatorium chinense in 2013 [203]. Compounds 431a/431b and 432a/432b, rearranged acetophenones with a novel 9-oxatricyclo[3.2.1.13,8]nonane core from Melicope ptelefolia, were assigned the abs. configs. by single-crystal X-ray diffraction analysis [204].
2.11. Diarylheptanoids
Diarylheptanoids, a class of NPs characteristic of a 1,7-diphenylheptane core, have been increasingly recognized as potential therapeutic agents for their diverse biological properties including antiinflammatory, antitumor, antioxidant, antiestrogen, hepatoprotective, antileishmania and neuroprotective activities [205]. Nine pairs of diarylheptanoid enantiomers (433−441, for structures, see Figure 25; for names, see Table S25 in Supplementary Materials) have been documented in the covered period. The occurrence of enantiomerism in these compounds results from the chiral centers generated by oxidation or Diels-Alder cycloaddition with other molecules.
The enantiomeric pairs of diarylheptanoid-monnoterpene adduct (433 & 434) and diarylheptanoid-sesquiterpene hybrid (435–437) from Alpinia officinarum, were hypothesized to be produced via a crucial Diels-Alder cycloaddition between the diarylheptanoids and corresponding terpenyl units. The rel. configs. for the chiral centers in the cyclohexene ring were assigned by comparing the experimental and calculated 13C NMR data, followed by the establishment of the abs. configs. via the TDDFT-ECD method [206]. Compounds 438−441 are four pairs of diarylheptanoid enantiomers acquired from Dioscorea villosa in 2012, with the abs. configs. being determined by the modified Mosher’s method [207]. Compounds 438 and 439 incorporate an extra tetrahydropyran ring formed via C-1 and C-5, while 440 and 441 are normal linear examples.
2.12. Triphenylmethanes
Triphenylmethanes are a unique class of NPs with one central carbon being linked by three aryl groups. They have been discovered to have a wide range of biological activities including antioxidant, antitumor, anti-HPK (histidine protein kinases) activities, etc. [208]. Six pairs of triphenylmethane enantiomers (442−447, Figure 26, Table S26 in Supplementary Materials) were reported in this period; their enantiomerism is attributable to chiral centers (442 and 443) or axial chirality (444−447).
Compounds 442a/442b and 443a/443b are two pairs of triarylmethane enantiomers reported from Securidaca inappendiculata in 2018, with the abs. configs. being determined by X-ray crystallography [209]. In addition, bio-inspired total syntheses for these compounds were also completed [209]. Compounds 444−447 occurred as racemates generated by axial chirality in the plant Selaginella pulvinate [210], and subsequent chiral fractionation divided 444 and 447 into 444a/444b and 447a/447b, respectively, with the abs. configs. being assigned by TDDFT-ECD method. However, 445 and 446 had not been enantiomerically separated.
2.13. Fatty Acids
Five pairs of enantiomeric fatty acid esters (448a/448b−452a/452b) were recorded in this covered stage and their structures are listed in Figure 27, with names being shown in Table S27 in Supplementary Materials. Usually, the generation of chirality in these compounds derives from the nonstereoselective oxidations on the aliphatic chain (448−451) or substitution on the glycerol moiety (452). Compounds 448a/448b−452a/452b from Plantago depressa were characterized as four pairs of 9-oxo octadecanoid derivatives by the authors’ group, with 451 bearing a rare chlorine atom [6]. We have also isolated 452a/452b as octadecanoid monoglycerides from the seeds of Ipomoea nil in 2019 and established their abs. configs. via an in situ dimolybdenum ECD method [9].
2.14. Miscellaneous
Other enantiomeric NPs from plants reported in this period are displayed in Figure 28 (names see Table S28). The abs. configs. of all these enantiomers were assigned by the TDDFT-ECD method unless otherwise specified.
Compounds 453−456 are four pairs of enantiomeric phthalide derivatives, all of which were isolated and characterized from Angelica sinensis in 2018 [211]. Phthalides are a rare class of NPs referring to lactones of 2-hydroxymethyl benzoic acids. They exist in nature as monomers or oligomers, and the latter are generally produced via [2 + 2] or [4 + 2] cycloaddition to form a number of complex polycyclic skeletons with multiple chiral centers [211]. Among them, 453 and 454 are dimers, while 455 and 456 are trimers.
Compounds 457a/457b and 458a/458b are enantiomeric stilbenoids that have 1,2-diphenylethylene (stilbene) as their basic scaffold and exist as monomers or oligomers in nature. They normally act as phytoalexins to assist plants in their resistance to pathogens or stress factors [212]. Compounds 457a/457b, prenylated stilbenoid dimers isolated from Cajanus cajan in 2014, possess an interesting dimerization pattern generated from nonstereoselective radical addition as shown in Scheme 7, and their structures including the abs. configs. were determined by a combination of X-ray diffraction analysis and TDDFT-ECD calculation [213]. Compounds 458a/458b are enantiomeric stilbenoid trimers obtained from Cyperus rhizomes in 2012, and their abs. configs. were established by comparing the [α]D and ECD data with those of known analogues [214,215].
Compounds 459a/459b are butenolide derivatives isolated from Dendrobium nobile in 2016 [49], while 460a/460b, with an unprecedented skeleton incorporating both butyrolactone and butenolide moieties, were obtained from Melicope viticina in 2019 [202]. Compounds 461a/461b featuring an oxabicyclo[3.2.1]octane ring were discovered from Ligusticum chuanxiong in 2019 [216], and 462a/462b are a pair of enantiomeric cyclohexylethanoid dimers acquired from Incarvillea younghusbandii in 2012 [217]. Compounds 463a/463b from Dendrobium nobile were identified as a pair of spirodiketone enantiomers in 2016 [218]. Styrylpyrone monomer (464a/464b) and dimer (465) enantiomers were reported from Sanrafaelia ruffonammari and Ophrypetalum odoratum, respectively, but the dimer 465 was only obtained as a racemate without further chiral separation [219]. Compounds 466a/466b are a pair of enantiomeric 2,3-dihydro-1H-indene derivatives discovered from Streblus indicus in 2016 [220].
3. Enantiomers from Kingdom Fungi
Enantiomers originating from fungi, i.e., from phyla Ascomycota and Basidiomycota, will be presented in this section. The structural classification of NPs from fungi is not as regular and clear as those from plants; a myriad of fungal NPs belong to the super family of polyketides that derive biogenetically from the acetate pathway [1]. Also, considering the limited number of molecules described in this section, the enantiomers described here are simply divided into nonalkaloids and alkaloids. Where applicable, their biogenesis and structure will also be described.
3.1. Enantiomers from Phylum Ascomycota
3.1.1. Nonalkaloids
Nonalkaloid enantiomers from phylum Ascomycota show great structural diversity and biological importance. The structures of those documented in the covered period are summarized in Figure 29a,b, and their names are presented in Table S29 in Supplementary Materials. The abs. configs. of those established by TDDFT-ECD method are not specifically mentioned in this section.
Zhang and co-authors discovered a racemic polyketide 467, together with four pairs of analogue enantiomers 468a/468b–471a/471b, from the starfish-symbiotic fungus Penicillium sp. GGF16-1-2 in 2019 [221]. The enantiomeric cyclopentenones 472a/472b and spiro-butenolides 473a/473b were isolated from Aspergillus Sclerotiorum in 2019 [222]. Gao and coworkers investigated three endolichenic fungal strains Nigrospora sphaerica, Alternaria alternata and Phialophora sp. in 2016 and obtained the same polyketide enantiomers 474a/474b, whose abs. configs. were determined by modified Mosher’s method [223]. Compounds 475a/475b incorporating a benzannulated 6,6-spiroketal skeleton were isolated from the mangrove-derived fungus Penicillium dipodomyicola HN4-3A. Compounds 476a/476b, a pair of ketal enantiomers from Paraconiothyrium sporulosum, were assigned the abs. configs. by application of Snatzke’s chirality rule for cyclopentenones [224]. The isocoumarins 477 and 478 were reported as racemic mixtures from Penicillium coffeae MA-314 in 2019 [225]. Compounds 479a/479b and 480a/480b are spiro-orthoester enantiomers bearing a novel 1,4,6-trioxaspiro[4,5]decane-7-one unit from Penicillium minioluteum, and their rel. configs. were assigned by single-crystal X-ray diffraction analysis [226]. Compounds 481a/481b were characterized as a pair of cyclopentaisochromenone enantiomers from Alternaria sp. TNXY-P-1 in 2018 [227]. Puno and coworkers discovered 482a/482b as dibenzo-α-pyrones bearing a diepoxy-cage-like moiety from an Endophytic Alternaria sp. in 2019 and confirmed their rel. configs. by X-ray crystallography [228]. Also elucidated as dibenzo-α-pyrones, 483a/483b and 484a/484b were reported from the endophytic fungus Alternaria alternate in 2014 [229]. Compounds 485a/485b, a pair of enantiomeric chromone derivatives from the marine-derived fungus Taeniolella sp. BCC31839, were established the abs. configs. by the modified Mosher’s method in 2019 [230]. Compound 486 was obtained as a racemate from Periconia sp. in 2015, without further chiral fractionation [231], while 487a/487b and 488a/488b were isolated from the endophytic fungus Aspergillus Fumigatus in 2018 [232]. Compound 489 was elucidated as a racemic mixture from the cordyceps-colonizing fungus Fimetariella sp. in 2012, and it incorporates a novel spiro[chroman-3,7′-isochromene]-4,6′(8′H)-dione skeleton [233]. Compounds 490a/490b were characterized as a pair of enantiomeric isochromanes from an endophytic fungus Aspergillus fumigatus in 2019 [234]. Enantiomers 491a/491b and 492a/492b were identified as p-terphenyl derivatives from the endolichenic fungus Floricola striata [235], with the abs. configs. being determined by using the helicity rule for α,β-unsaturated ketone. Compounds 493a/493b and 494a/494b, as xanthene enantiomers with an unprecedented hexacyclic heterocylic backbone, were isolated from Xylaria feejeensis GM06 in 2018 [236]. The abs. config. assignment for 493a/493b was based on the X-ray crystallographic experiment. Compounds 495a/495b were characterized as a pair of dimeric polyketide enantiomers from a mangrove endophytic fungus Ascomycota sp. SK2YWS-L [237], with the absolute structures being determined by X-ray diffraction analysis and TDDFT-ECD calculation. Compounds 496a/496b are a pair of 2,3-diaryl indone atropisomers isolated from Ascomycota sp. SK2YWS-L in 2018 [238].
Compounds 497a/497b were identified as simple δ-lactone enantiomers from the fungus Aspergillus terreus in 2018 [239], while 498 is a α-pyrone derivative obtained as a nearly racemic mixture from the endolichenic fungus Tolypocladium sp. in 2017 [240]. Pei and coworkers discovered two pairs of dimeric α-pyrone enantiomers (499a/499b and 500a/500b), which was formed via intermolecular nonstereoselective [2 + 2] cycloaddition reaction (Scheme 8), from the endophytic fungus Phoma sp. YN02-P-3 in 2017, and 499a/499b possess a novel 6/4/5/6 tetracyclic ring system. Moreover, the rel. config. assignment for 500a/500b was confirmed by single-crystal X-ray diffraction analysis [241]. Compounds 501 and 502 were elucidated as C-ring open flavonoids from Pochonia chlamydosporia var. spinulospora FKI-7537 in 2018, and 502 was successfully resolved into two enantiomers but without assigning the abs. configs., while 501 was not subjected to chiral separation due to limited amount [242]. Compounds 503a/503b and 505a/505b are two pairs of polyketides isolated from Penicillium chrysogenum MT-12 in 2017, where the racemic mixture of 505 had been previously reported from an endophytic fungus Aspergillus sp [243]. Compounds 504a/504b, a pair of funicone enantiomers, were obtained from the mangrove sediment-derived fungus Penicillium pinophilum SCAU037 [244]. The polyketide dimers 506a/506b and 507a/507b bearing a rare pentacyclic dihydrobenzo[1,4]dioxine core were isolated from Penicillium canescens in 2019 [245]. Enantiomers 508a/508b, a pair of caged norsesquiterpenoids with a novel tricyclo[4.4.01,6.02,8]decane carbon skeleton, were obtained from the endophytic fungus Preussia isomera in 2019, with the rel. config. being confirmed by X-ray diffraction data [246]. Kong and coworkers discovered 509a/509b, featuring a prenylated chlorobenzophenone backbone, from the plant endophytic fungus Pestalotiopsis sp. in 2017 [247]. Compounds 510a/510b are 2-benzofuran-1(3H)-one derivatives isolated from a mangrove-derived fungus Eurotium rubrum MA-150 in 2016 [248]. Compounds 511a/511b−513a/513b, three prenylated dibenzo[b,e]oxepinone enantiomer pairs, were reported from a wetland soil-derived fungus Talaromyces flavus in 2016 [249], and the same type of enantiomers 514a/514b were obtained from an endophytic fungus Xylaria sp. in 2015 [250]. The benzophenone-hemiterpene adducts 515a/515b were separated from the endophytic fungus Cytospora rhizophorae in 2019 [251]. Compounds 516a/516b are a pair of enantiomeric polyketides incorporating a 6/6/6/6/5/6/6 heptacyclic backbone and were isolated from fungus Alternaria sp. MG1 in 2019 [252]. Compounds 517a/517b were identified as dimeric polyketide enantiomers from a marine-derived fungus Eurotium sp. SCSIO F452 in 2019 [253].
3.1.2. Alkaloids
Alkaloid enantiomers found in phylum Ascomycota during the reported period are displayed in Figure 30, with their names being shown in Table S30 in Supplementary Materials. The abs. configs. of those established by TDDFT-ECD method are not specifically mentioned in this section.
Compounds 518−537 are a series of indole alkaloid derivatives reported from different fungal species. Except for the bisindole enantiomers 518a/518b (from Fusarium sp. XBB-9), whose abs. configs. were assigned by X-ray diffraction analysis [254], all other alkaloids possess a cyclodipeptide scaffold (also known as diketopiperazine) formed from tryptophan and a second amino acid. Four pairs of enantiomers (519a/519b−522a/522b) biosynthesized from tryptophan and proline were isolated from the marine-derived species Aspergillus versicolor OUCMDZ-2738 in 2019 [255], while the tryptophan-alanine dipeptide enantiomers 523a/523b−525a/525b bearing rare thiomethyl and N-methoxy groups were obtained from an alga-derived endophytic fungus Acrostalagmus luteoalbus TK-43 in 2019 [256]. Wang and coworkers discovered the spirocyclic alkaloids 526a/526b−529a/529b from Eurotium sp. SCSIO F452 in 2019 and proposed their biosynthetic pathways as shown in Scheme 9 [257], while 529a/529b had also been reported from Aspergillus effuses H1-1 in 2012 [258]. Compounds 530a/530b, whose abs. configs. were determined by X-ray crystallography, incorporate a novel 6/5/4/5/6 pentacyclic motif and were acquired from the mangrove endophytic fungus Aspergillus sp. SK-28 in 2019. As shown in Scheme 10, the nonenzymatic catalyzed [2 + 2] cycloaddition could be the plausible key biosynthetic step to generate both enantiomers of 530 [259]. Alkaloids 531a/531b with a 3′,3a′,5′,6′-tetrahydrospiro[piperazine-2,2′-pyrano[2,3,4-de]chromene] ring system were isolated from a mangrove rhizosphere soil derived fungus Aspergillus effuses H1-1 in 2012 [258], and compounds 532a/532b−534a/534b as three pairs of variecolortide enantiomers were reported from the fungus Eurotium sp. [260]. Wang and coworkers disclosed three spirocyclic diketopiperazine enantiomer pairs (535a/535b−537a/537b) from the marine-derived fungus Eurotium sp. SCSIO F452 in 2019 and proposed their plausible biosynthetic pathways as shown in Scheme 11 [261]. Compounds 535a/535b possess a highly functionalized seco-anthronopyranoid structural unit with a 2-oxa-7-azabicyclo[3.2.1]octane core, while 536a/536b and 537a/537b represent rare examples of diketopiperazines with a 6/6/6/6 tetracyclic cyclohexene-anthrone fragment.
The decalin-containing 4-hydroxy-2-pyridones (538a/538b) and their four pairs of rearranged analogues (539a/539b−542a/542b) were isolated from the solid culture of fungus Coniochaeta cephalothecoides in 2017 [262]. In 2019, Liu and colleagues investigated the metabolites of fungus Xylaria longipes to detect two highly conjugated alkaloids, 543a/543b and 548a/548b. The former possesses a 5/6/6/5/5 fused ring system with a unique 2-azaspiro[4.4]nonane substructure [263]. From the same fungal species, the same authors reported 544a/544b and their dimers 545a/545b [264] as thiopyranodipyridine enantiomers. Compounds 546a/546b and 547a/547b were identified as N,N′-ketal quinazolinone alkaloid enantiomers from an ascidian-derived fungus Penicillium sp. 4829 in 2019 [265] and from an algicolous Talaromyces sp. in 2016 [266], respectively. Compounds 549a/549b are a pair of enantiomeric 4-oxabicyclo[4.3.0]lactam derivatives from the marine-derived fungus Penicillium griseofulvum reported in 2017 [267]. The aromatic polyketide enantiomers 550a/550b with a 5/6/6/6/5 heterocyclic architecture were separated from Penicillium canescens in 2019 [245], while the enantiomeric phthalimidine derivatives 551a/551b were acquired from the sponge-derived fungus Stachylidium sp. in 2012 [268]. Compounds 552a/552b were characterized as a pair of N-furanone amide enantiomers from the solid culture of Trichoderma atroviride S361 in 2018 [269], and 553a/553b, a pair of enantiomeric hydantoin (imidazolidin-2,4-dione) derivatives, were obtained from the fungus Fusarium sp. in 2015 [270]. Compounds 554−557 were isolated as bisabolane sesquiterpenoid amide racemates from the plant endophytic fungus Paraconiothyrium brasiliense in 2015, but only 554 was chirally separated into pure enantiomers [271]. Compounds 558a/558b are a pair of enantiomeric alkaloid dimers with a symmetrical spiro[oxazinane-piperazinedione] skeleton from Pestalotiopsis sp. in 2015 [272].
3.2. Enantiomers from Phylum Basidiomycota
It is interesting to note that all natural enantiomers from phylum Basidiomycota collected in this period, with only one exception (Granulobasidium vellereum), were reported from species of the well-known medicinal macrofungus genus Ganoderma. More interestingly, all the enantiomers from Ganoderma fungi, with one exception., are hydroquinone derivatives (602). In addition, the majority of these enantiomers belong to the meroterpenoid class (hydroquinone-terpenoid hybrid), and the terpenyl units here are usually monoterpenoid or sesquiterpenoid. Their structures and names are summarized in Figure 31a,b and Table S31, respectively. The abs. configs. of these enantiomers in this section have all been determined by TDDFT-ECD calculation unless otherwise specified.
Compounds 559–584 represent monomeric hydroquinone-terpenoid enantiomers. Cheng and coworkers discovered a pair of hydroquinone-trinorsesquiterpenoid enantiomers (559a/559b) possessing a fused 6/5/6/6/5 polycyclic skeleton from G. lucidum in 2019 [273]. Compounds 560a/560b−563a/563b, identified as a series of hydroquinone-mononorsesquiterpenoid hybrids from G. cochlear in 2014, possess a spiro[4,5]decane ring system (560−562) and an eight-membered ring (563), with the abs. configs. being assigned by single-crystal X-ray diffraction analysis [274]. Compounds 564a/564b from G. lucidum are a pair of rotary door-shaped hydroquinone-normonoterpenoid enantiomers with an unusual 5/5/6/6 ring system, and their abs. configs. were established by interpretation of X-ray crystallographic data [275]. Compounds 565a/565b, a pair of macrocyclic meroterpenoid enantiomers derived from a hydroquinone and an intact sesquiterpenoid, were isolated from G. resinaceum by Chen et al. in 2017 [276]. The hydroquinone-monoterpenoid enantiomers 566a/566b with an unusual dioxacyclopenta[c,d]inden motif were reported from G. applanatum in 2016 [277]. Nine pairs of enantiomers 567a/567b−575a/575b incorporating either monoterpenoid or sesquiterpenoid fragments were obtained from G. applanatum in 2015 [278], and 570a/570b was also reported from G. lucidum in the same year with the abs. configs. being not assigned [279]. Compounds 576a/576b featuring an interesting polycyclic meroterpenoid skeleton with a glycerol unit were isolated from G. applanatum in 2017 [280]. Five pairs of hydroquinone-sesquiterpenoid enantiomers 577a/577b−581a/581b and a racemate 582, all bearing a butenolide fragment, were isolated from G. sinense in 2016 [281]. Compounds 583a/583b and 584a/584b, two pairs of farnesylated hydroquinone enantiomers incorporating a p-hydroxycinnamoyl residue, were discovered from G. sinense in 2016 [281].
Compounds 585–600 represent dimeric hydroquinone-terpenoid enantiomers. Enantiomers 585a/585b were elucidated as hydroquinone dimers hybridized with a highly oxygenated monoterpenoid moiety from G. applanatum in 2016 [282]. They feature an unprecedented dioxaspirocyclic skeleton constructed from a 6/6/6/6 tetracyclic system and an unusual tricyclo[4.3.3.03′,7′]dodecane unit, and their abs. configs. were determined by single-crystal X-ray diffraction analysis [282]. Also from G. applanatum, Cheng and coworkers separated three pairs of dimeric hydroquinone-monoterpenoid enantiomers 586a/586b−588a/588b [283]. Two types of meroterpenoid heterodimer enantiomers (589a/589b & 590a/590b; 591a/591b–593a/593b) from G. cochlear were reported by the same authors from Cheng’s group, and their abs. configs. were assigned by comparing the ECD curves with those of reported analogues [284,285]. Five pairs of enantiomers (594a/594b−598a/598b) of the same type as 591–593, along with the novel hybrid dimers (599a/599b & 600a/600b) formed by a hydroquinone-pyridine and a hydroquinone-monoterpenoid, were also isolated and characterized from G. cochlear in 2015 [286].
Compounds 601a/601b were identified as a pair of hydroquinone-pyridine alkaloid enantiomers from G. luteomarginatum in 2019 [287], while butyrolactone 602 from G. lucidum was chirally separated without assigning the abs. configs. of the enantiomers [279]. In 2015, sesquiterpenoids 603a and 604a [288] from the fungus Granulobasidium vellereum were identified as the enantiomers of illidin M (603b) [289] and dihydroilludin (604b) [290], respectively.
4. Enantiomers from Kingdom Prokaryota
Few enantiomers have been reported from the kingdom Prokaryota, i.e., only five pairs (605–609) to date, all of which were discovered from actinomycetes. Their structures and names are provided in Figure 32 and Table S32 in Supplementary Materials, respectively.
Compounds 605a/605b are a pair of angucyclinone enantiomers featuring a unique epoxybenzo[f]naphtho[1,8-bc]oxocine heterocyclic scaffold, and were isolated from a Streptomyces sp. in 2019, with the abs. configs. being determined by X-ray diffraction analysis [291]. Compound 606, a simple prenylated indole alkaloid bearing a rare cyano group, was isolated as a racemate without further chiral separation from Streptomyces sp. ZZ820 in 2019 [292]. Compounds 607a/607b−609a/609b are three pairs of enantiomeric indole alkaloids with a spiro indolinone-naphthofuran skeleton reported from a Streptomyces sp. in 2017 [293].
5. Enantiomers from Kingdom Animalia
Compared with those from plants and microorganisms, compounds from animals only account for a small proportion of the large NP family, and have been mainly reported from lower animals such as sponges and corals. Therefore, the number of enantiomers from kingdom Animalia is also limited. According to their biological source, animal-derived enantiomers will be divided into the following three subcategories.
5.1. Enantiomers from Phylum Porifera
Animals from phylum Porifera (also termed Spongia) are generally known as sponges. They also represent a very important source of bioactive NPs. Natural enantiomers from sponges mainly include terpenoids and alkaloids; see Figure 33 and Table S34.
5.1.1. Terpenoids
Compounds 610a/610b were identified as a pair of valerenane-type sesquiterpene enantiomers from a Spongia sp. in 2019 [294], while the trinorsesquiterpenoid enantiomers 611a/611b incorporating furan and butenolide rings were isolated from the Beihai sponge Spongia officinalis in 2018, with the abs. configs. being determined by biomimetic total synthesis and modified Mosher’s method [295]. The three C17 norditerpenoid pairs 612−614 with a γ-lactone unit, together with two pairs of sesterterpenoids 615 and 616 with a butenolide unit, were obtained from a Cacospongia sp. in 2019. Among them, all enantiomeric pairs except 614 were successfully separated. Compounds 617a/617b are a pair of sesterterpenoid enantiomers featuring a bicyclo[4.2.0]octene core and were isolated from Hippospongia lachne in 2017 [296]. Compounds 618−621 are four pairs of furanosesterterpene tetronic acids from a Psammocinia sp., and 618 and 619 were found to be geometrical isomers of two pairs of enantiomers as revealed by chiral HPLC analysis. Similar to the case of 618 and 619, compounds 620 and 621 were also proved to be two enantiomeric pairs, but only 620 was finally separated into pure enantiomers [297].
5.1.2. Alkaloids
Interestingly, alkaloid enantiomers from sponges were discovered from species collected in South China Sea, with most of them belonging to the pyrrole alkaloid family, incorporating a pyrrole-2-carboxylic acid residue. Compounds 622a/623b−628a/628b were characterized as a panel of bromopyrrole enantiomers from an Agelas sp. in 2016 by Zhu et al. [298]. Except 622a/622b and 627a/627b, the abs. configs. of the others were assigned by one of the three following methods including TDDFT-ECD calculation, ECD exciton chirality method and ECD comparison with known analogues [298]. Pyrrole alkaloids 629a/629b−631a/631b are three pairs of enantiomers obtained from Agelas aff. Nemoechinata in 2017, and 631a/631b possess an interesting cyclopentane-fused imidazole ring system [299]. Compounds 632a/632b−634a/634b are also pyrrole alkaloid enantiomer pairs obtained from Agelas nakamurai in 2017 [300]. Alkaloids 635a/635b featuring an unusual spiro bisheterocyclic quinoline-imidazole backbone were reported from Fascaplysinopsis reticulate in 2015 [301], while 636a/636b represent a pair of trinorsesquiterpenoid amide enantiomers isolated from the Beihai sponge Spongia officinalis in 2018 [295].
5.1.3. Lipids
Compounds 637a/637b, a pair of interesting C20 bisacetylenic lipid enantiomers, were discovered from the marine sponge Callyspongia sp. in 2013, with the abs. configs. being determined by modified Mother’s method [302]. The lipid zwitterions 638a and 639a were separated from Spirastrella abata in 2012 [303], and their respective enantiomers (638b and 639b) had been previously reported from the same species in 2002 [304].
5.2. Enantiomers from Phylum Arthropoda
Compounds 640a/640b−643a/643b (Figure 34, names see Table S34 in Supplementary Materials) bearing a 2,3-dihydrobenzo[b][1,4]dioxin fragment were separated from the insect Blaps japanensis in 2015 [305], with the abs. config. of 640a being determined by X-ray crystallographic analysis. Compounds 644a/644b and 645 were characterized as N-acetyldopamine dimer and trimer, respectively, from the insect Aspongopus chinensis in 2014, and 645 possesses a novel tetrahydrobenzo[a]dibenzo[b,e][1,4]dioxine moiety and occurs as a racemate [306]. Compounds 646a/646b are a pair of dimeric N-acetyldopamine enantiomers obtained from the insect Polyphaga plancyi in 2016 [307].
5.3. Enantiomers from Phylum Chordata
There have been only two pairs of enantiomers reported from the animals of phylum Chordata (see Figure 34 and Table S34 in Supplementary Materials). Compounds 647a/647b, a highly nitrogenated enantiomer pair with a novel heterocyclic scaffold incorporating two extra phenol units, were isolated from a marine ascidian Eudistoma sp. in 2016 [308]. A pair of oxygenated myristic acid enantiomers bearing a tetrahydrofuran moiety (648a/648b) was obtained from a larval sea lamprey Petromyzon marinus in 2015, with the abs. configs. being determined by modified Mosher’s method [309].
6. Biological Properties
As is well known, NPs, on one hand, play a decisive role in maintaining their source organisms’ health, helping defend against internal or external adverse stresses and enticing favorable stimuli. On the other hand, NPs in the form of herbal medicines have long been used by humans as therapeutic agents against various diseases, thus guaranteeing the continuation of human civilization. With the advances of science and technology, NPs and their derivatives still shine in the research field of modern drug discovery and development [2].
It is widely accepted that chirality, as an important feature of most NPs, is closely related with their bioactivities. Normally, life systems tend to produce/utilize only one molecule of an enantiomeric pair. For example, humans only take in d-glucose and l-amino acids as nutrients. The fact that a pair of enantiomers can exert utterly different bioactivities was recognized as far back as the 1960s, when the ‘Phocomelia infants event’ caused by the (S)-enantiomer of the synthetic drug thalidomide taught the pharmaceutical industry an important lesson. For many years, however, NP workers failed to recognize the widespread occurrence of enantiomerism in nature, and failed to explore the differences in bioactivity between pairs of enantiomers. Fortunately, as data on natural enantiomers increase in scope, more and more biological properties of different classes of enantiomeric pairs have also been reported, and this has provided more examples with which to investigate the differences in bioactivity among enantiomers.
As bioassay protocols vary in different research labs and even in different batches from the same lab, it should be clarified that we do not intend to invite direct comparisons regarding the activity potency by tabulating the assay data from different reports. Instead, bioactivity comparisons between different labs will be completely avoided in the current review and the use of potency descriptors will also be kept to a minimum. Meanwhile, we will not list the biological data of all reported enantiomers, and only selective cases with obvious activity differences at the enantiomeric level are discussed, under the following subcategories: cytotoxic, antiviral, antibacterial, antifungal, anti-inflammatory, antioxidative, cell protective, enzyme inhibitory, β-amyloid (Aβ) aggregation inhibitory and miscellaneous activities.
6.1. Cytotoxicity
Cytotoxic evaluations of chemical entities were likely the most important primary strategy in the past in the search for potential chemotherapies for cancers, and remain among the most popular bioassays for NPs. The cytotoxic activities of selective enantiomeric pairs against a series of human tumor cell lines are summarized in Table 2.
The antiproliferative activities of the alkaloid racemates (±)-171 and (±)-172, along with their respective enantiomers, against human HL-60 tumor cells were assessed in Hua’s lab. While the levoisomers (171b and 172b) showed slightly better inhibitory activity than their respective dextroisomers (171a and 172a), the racemic mixtures exhibited more potency than both enantiomers, indicating a likely synergistic effect [105]. The flavane enantiomers 268a/268b were reported to show cytotoxicity against human Hep3B cells, and the dextroisomer 268b was obviously more active than the levoisomer 268a [146]. The xanthones 297a/297b were able to inhibit the proliferation of human HL-60 and MDA-MB-231 cancer cells, and the (−)-enantiomer 297b showed much stronger inhibitory activity than its (+)-enantiomer 297a against MDA-MB-231 cells [156]. The levorotatory enantiomer 312b was found to inhibit the proliferation of colorectal HCT-116 cell line with an IC50 of 14.23 μM, but its antipodal enantiomer 312a was considered to be inactive [160].
The bisabolene-derived sesquiterpenoids 314a/314b were tested in vitro for their cytotoxicities against five human tumor cell lines (HCT-116, HepG2, BGC-823, NIC-H1650 and A2780), and the (−)-enantiomer 314a exerted significant inhibition against all tested cell lines with IC50 values in the range of 1.38−8.19 μM, while the (+)-enantiomer 314b was considered inactive (IC50 > 10 μM) [162]. In contrast, the (+)-enantiomer 381a, an acylphloroglucinol derivative, showed cytotoxic activities against the tested tumor cell lines (HL-60, SMMC-7721, A549, MCF-7 and SW480) with IC50 values in the range of 3.42−7.22 μM, while its (−)-enantiomer 381b was taken inactive (IC50 > 20 μM) [191]. The same research group that reported 381a/381b also screened the cytotoxicities of both racemates and pure enantiomers of acylphloroglucinols 382a/382b−385a/385b against the aforementioned tumor cell lines [192], and as a result, the levorotary series exhibited higher potency than both the dextrorotary series and the racemates for most cells [192]. Of particularly note, the racemate (±)-383 showed apparently decreased activity compared with its both enantiomers against all cell lines especially toward HL-60 cells (5.7- and 7.7-fold decrements), indicative of an antagonistic action between the two enantiomers, and similar effects were also observed for 384a/384b and 385a/385b on selective cell lines [192].
The methyl 2-naphthoate enantiomers 390a/390b were found to show inhibition against the proliferation of three types of cancer cells (A549, MCF-7 and MDA-MB-231), with the levoisomer 390a being ca. three times more active than the dextroisomer 390b [8]. In addition, only the dextrorotary enantiomers of the spirocyclic diketopiperazines 536 and 537 showed growth inhibition against SF-268 and HepG2 tumor cell lines [261], but their corresponding levoisomers were inactive (>100 μM).
6.2. Antiviral
The antiviral activities of the enantiomers described in this review are shown in Table 3. The coumarins 254a/254b did not show significant inhibition differences against either the herpes simplex virus 1 (HSV-1) or the host cell between enantiomers, but their racemate (±)-254 exhibited obviously increased activity which was suggestive of a strong synergistic action [66]. Similar synergistic effects of enantiomers were also observed for another two pairs of coumarins, 104a/104b and 105a/105b, with the racemates (±)-104 and (±)-105 displaying 3.2- to 6.1-fold antiviral activity against the influenza virus A (H3N2) compared with the pure enantiomers [67]. Four pairs of phloroglucinol enantiomers 368a/368b−371a/371b were subjected to antiviral assay against Kaposi’s sarcoma-associated herpes virus (KSHV); they all showed certain degrees of bioactivity differences at the enantiomeric level [184]. The fungus-derived alkaloid enantiomers 558a/558b and the racemate (±)-558 all exhibited antiviral activity against EV71 virus, with the dextrorotary enantiomer being nearly five times as active as its antipodal isomer [272].
6.3. Antibacterial
It appears that most of the antibacterial enantiomeric pairs collected in this review showed remarkably differentiable activities between enantiomers. Nonetheless, a few exceptions were still found and are listed in Table 4. The furoquinoline alkaloid enantiomers 147a/147b were reported to have antibacterial activity against Enterococcus faecalis, and the (–)-enantiomer showed about two-fold activity as the (+)-enantiomer [97]. The p-hydroxycinnamoylated dihydrochalcone enantiomers 285a/285b−288a/288b exhibited in vitro antibacterial activity against Staphylococcus aureus with IC50 values ranging from 0.61 to 6.0 μM [153], and it appeared that all the dextrorotary enantiomers were more effective than their respective levorotary isomers, with 288a/288b showing the greatest activity difference, i.e., 3.7 fold [153].
6.4. Antifungal
Few reports have been published on the antifungal activities of the enantiomers mentioned in this review, although a handful of examples have shown about two-fold bioactivity differences between enantiomers (Table 5). The δ-lactone enantiomer (−)-464a was reported to display inhibitory activity against Candida albicans with an MIC of 26.4 μM, while its antipodal enantiomer (+)-464b was considered inactive [219]. In addition, both levorotary enantiomers of compounds (±)-483 and (±)-484 exhibited better antifungal activity again C. albicans than their respective dextrorotary isomers [229], and similar effect against Fusarium solani was also recorded for the indole-piperidine enantiomer pair (±)-524 [256].
6.5. Anti-Inflammation
The anti-inflammatory activities of NPs have often been evaluated by testing their inhibitory capability against NO release in LPS-induced BV-2 microglial cells or RAW 264.7 macrophages (Table 6). The benzofuran-type lignan enantiomers 43a/43b and 44a/44b were tested for their NO production inhibitory effect in LPS-induced BV-2 microglial cells, with (−)-43b and (+)-44a exhibiting pronounced activity with IC50 values of 8.9 and 5.9 μM, being nearly twice as active as their respective antipodal enantiomers [39]. The levorotary spirodienone lignan enantiomers (−)-82b and (−)-83b showed significant inhibition against NO production in LPS-induced RAW 264.7 macrophages, with both being >3 fold as active as their respective dextrorotary enantiomers [54]. In the same bioassay model, the indolizidine dextroisomer 221a displayed much stronger inhibitory activity (6.3 fold) than the levoisomer 221b [124,128,164]. In contrast, the levorotary enantiomer 407b was much more active (ca. 5 fold) than its antipodal enantiomer 407a in the LPS-induced NO release assay in BV-2 cells [130,132,226,265].
6.6. Antioxidation
The DPPH and ABTS radical scavenging assay models have been widely used to evaluate the antioxidative capacity of NPs, although not many of the listed enantiomers in this review have been tested with these bioassays. Owing to their radical mechanism, most tested enantiomers displayed equal potency in both assays as expected, whereas the tryptophan-alanine dipeptide enantiomers 527a/527b–529a/529b showed some activity differences at the enantiomeric level, particularly for dextroisomers 527a and 528a, that showed obviously enhanced radical scavenging activity (4.1 and 2.5 fold, respectively) compared with their levoisomers in the DPPH assay model (Table 7) [257].
6.7. Cell Protection
Cell protection assays are usually performed in neuronal cells to explore new chemicals that could be developed for the treatment of neurodegenerative disorders, but which could likely also be used in the search for molecules with which to treat other diseases (Table 8). Generally speaking, a >10% cell viability difference can be considered significant. The protective activity of neolignan enantiomers 24a/24b against H2O2-induced cell injury in human neuroblastoma SHSY5Y cells was tested and the (+)-enantiomer showed obviously better activity than the (−)-enantiomer [20]. In a same assay model by Zhou et al., the analogous enantiomeric pair 72a/72b also exhibited a similar trend of bioactivity difference, with the dextroisomer displaying better protective effect than the reference drug and the levoisomer being found to be inactive [46]. Further investigations revealed that (+)-72a could significantly decrease the percentages of both early and late apoptotic cells. The phenylpropanoid dextrorotary enantiomer 119a presented much better neuroprotective activity than its levorotary enantiomer in the H2O2-treated SH-SY5Y cell injury assay, with nearly 20% cell viability increment [75]. Further studies demonstrated that (+)-119a could selectively inhibit the apoptosis induction and reactive oxygen species (ROS) accumulation by enhancing the activity of catalase (CAT). Compared with their respective antipodal enantiomers, indole alkaloids (−)-129b and (+)-130a also increased the cell viability by about 20% in an OKA-induced PC12 cell damage assay [83]. The isoquinoline alkaloids (−)-165a, (−)-166a and (+)-173b exhibited slightly better protective effects (51%−55% cell viability) than the positive control on hypoxic H9C2 cells, while (+)-166b were less active (45% cell viability) and (+)-165b and (−)-173a were considered inactive [103]. Two pairs of acetophenone enantiomers 423a/423b and 426a/426b exerted excellent protection on human vein endothelial cells (HUVEC) against extreme glucose-induced oxidative stress at 1 μM [200], with both dextrorotary enantiomers being much more active than their levorotary counterparts and showing complete cell protection. The (+)-enantiomer of diarylheptanoids (±)-433 significantly increased the cell viability of cortical neurons compared with the control group (MPP+ treatment alone), while its (−)-enantiomer was inactive [206].
6.8. Enzyme Inhibition
A number of diseases are caused by the dysfunction of enzymes, so the discovery of enzyme inhibitors is one the most important tasks of the study of NPs. The enantiomers in this review have been shown to exert inhibitory activities against many enzymes including phosphodiesterase-9A (PDE9A), acetylcholinesterase (AChE), butyrylcholinesterase (BChE), α-glucosidase, tyrosinase, protein tyrosine phosphatase 1B (PTP1B), serine protease HLE, isocitrate lyase deubiquitinating enzyme USP7, isocitrate lyase, Na+/K+-ATPase and cyclooxygenase 2 (COX-2). Selective enantiomeric pairs with activity differences between enantiomers are listed in Table 9.
Isoquinoline enantiomers 157a/157b were evaluated for their anti-AChE activity, with the (−)-enantiomer being >3.5 fold more active than the (+)-enantiomer [100]. In the same assay from another lab, the dextrorotary indole-diketopiperazine enantiomer 523a was reported to be six times as active as its antipodal enantiomer, and their racemate showed a compromised activity [256]. The fungus-originated xanthones 495a/495b and their racemate (±)-495 were identified as potent α-glucosidase inhibitors with the levoisomer showing stronger activity [237]. The meroterpenoid enantiomers (+)-339b, (+)-340a, (−)-341a and (−)-342a displayed inhibitory effects against PTP1B with IC50 values ranging from 38.1 to 61.0 μM [176], while their respective antipodal enantiomers were considered inactive [178]. Two pairs of N-acetyldopamine enantiomers (641 and 644) derived from insect exhibited inhibitory activity against COX-2 with IC50 values in the range of 2.52–17.8 μM [305], and (+)-641a and (−)-644b were around two times as active as their respective antipodal enantiomers.
6.9. Aβ Aggregation Inhibition
Eleven pairs of plant-originated enantiomers including lignans (1a/1b, 2a/2b, 6a/6b, 7a/7b, 73a/73b and 76a/76b) and alkaloids (132a/132b, 133a/1333b, 241a/241b and 242a/242b) were evaluated for their inhibitory effects on β-amyloid (Aβ) aggregation, which had been considered as a central event in the pathogenesis of Alzheimer’s disease according to the “amyloid hypothesis”. Still, most enantiomeric pairs did not show much difference in Aβ aggregation inhibitory activity. Nevertheless, four pairs (6a/6b, 73a/73b, 133a/1333b and 242a/242b) did display obvious activity variations at the enantiomeric level (Table 10). Notably, the 8,4′-oxyneolignan pair 6a/6b presented a significant gap in their inhibition against Aβ aggregation, with the (−)-enantiomer showing a 141% activity increment compared with the (+)-enantiomer [17].
6.10. Miscellaneous Activities
In addition to the above-described biological properties, the enantiomers covered by the current review also showed positive responses in a variety of other bioassays. Those with obvious activity differences between enantiomers are listed in Table 11. The levorotary indole-diketopiperazine enantiomer (−)-145a exerted impact on MT1 and MT2 receptors with agonistic rates of 11.26% and 52.44% (at 0.25 mM), respectively, while its enantiomer (+)-145b was evaluated as inactive [93]. The (−)-enantiomer of diterpenoid 330 exhibited NF-κB inhibition with an IC50 value of 7.27 μM, while its (+)-enantiomer was considered inactive [169]. The fungus-derived indole-diketopiperazine enantiomer (+)-530b displayed antifouling activity against the barnacle Balanus reticulatus with an adhesive rate of 48.4% at 10 μg/cm2, while the (−)-enantiomer 530a was inactive [259]. The nitrogen-rich alkaloid enantiomer (−)-647b was identified as a moderate protein-protein interaction inhibitor of HIF-1α and p300, while its antipodal enantiomer (+)-647a was inactive [308]. Lastly, the fish-produced dextrorotary lipid enantiomer (+)-648a elicited a strong olfactory response on the sea lamprey, and its levorotary enantiomer (−)-648b only showed weak activity [309].
7. Conclusions
As can be seen from Figure 35, the number of identified natural enantiomers steadily increased during the period covered by this study, albeit with slight drops in 2013 and 2018. Notably, more than 100 enantiomers have been reported in the last three years (2017–2019), indicating rapid development in this field. It is also worth noting that plant-derived enantiomers made up 72% of all cases (Figure 36) in the study period, which suggests the continuing vitality of phytochemical studies, despite severe funding cutbacks for traditional NP research in recent years [3]. Another set of statistics (Figure 37) revealed that alkaloid enantiomers represent the biggest group of molecules from plants, followed by lignans and flavonoids.
7.1. Natural Distribution of Enantiomers
As demonstrated by the examples in this review, natural enantiomers have been widely reported from species of all kingdoms except Protoctista, which could be attributed to the fact that few NP researchers have been focusing on Protoctista organisms since they are not well-known sources of interesting molecules. Therefore, the discovery of enantiomers from Protoctista species in the near future is to be expected if NP workers continue to focus on them. From another perspective, the enantiomers collected in the period covered in this review have a broader distribution at the originated species level, from microbial fungi (e.g., mold) to macrofungi (e.g., mushrooms), from lower plants (e.g., moss) to higher plants (e.g., herbs), and from lower animals (e.g., sponges) to higher animals (e.g., fishes). Another noteworthy point is the distribution of enantiomers in different structural families, which can be clearly revealed by the examples in the current and previous reviews [3] that were discovered in all major structural classes such as terpenoids, alkaloids, flavonoids and polyketides (mainly from a biogenetic view). At the lower level of classification, it seems that there have been no enantiomeric cases reported for triterpenoids and steroids. The above-mentioned two points clearly demonstrate the universal occurrence of enantiomerism in nature.
7.2. Natural Formation of Enantiomers
It is interesting to note that unlike the previous report [3], in which many enantiomeric examples were obtained from different species, the majority of the cases collected in the current study were isolated from the same species as scalemic or racemic mixtures. Although Williams and colleagues predicted in 2012 [3] that the biogenetic studies of natural enantiomers would be “a fertile area for future inquiry and discovery”, there has been no significant progress in this research field since then. Nonetheless, some common reasons or rules regarding enantiomeric production can still be rationalized on the basis of currently available knowledge: (1) For cases in which the two antipodal enantiomers are produced by two different species (from the same or different genus or even different families), such as (+) and (–)-limonenes [3], two distinct enzymes and mechanisms are involved in their biosynthesis; (2) When an enantiomeric pair (racemic or scalemic mixture) is discovered from the same species, the lack (partially or completely) of stereo-specificity of the catalytic enzyme could be responsible for the enantiodivergent formation; (3) The absence of enzyme substrate or a completely chemical process would also lead to the production of two enantiomers, which is especially true for many NPs with only one chiral center. The following two explanations, though not as reasonable as the above-mentioned three, could also not be excluded. (4) In some biochemical processes which involve radicals, though normally stereo-controlled by enzymes, the generation of enantiomers is possible due to the extremely high reactivity of radicals. (5) The extraction and isolation procedures of NPs could also lead to the formation of new chiral centers, and thus, the production of enantiomers [310,311]. At this point, these enantiomeric molecules should be classified as NP derivatives or artifacts.
7.3. Structures Tend to Exist as Enantiomers in Nature
With the discovery of more and more enantiomeric NPs containing diverse structures, it can be concluded that enantiomerism may occur for each structural type, although no enantiomers have been reported for triterpenoids and steroids. Compared with enantiomers from plants, those from microorganisms are able to incorporate more complicated structures, e.g., with high molecular weights. It is possible that the enzyme systems in microorganisms are not fully developed and stereoselectivity is lacking. With this investigation into the structures of enantiomers reported from 2012–2019 in hand, we can easily conclude which structures or which groups in the structures tend to exist in nature as enantiomers. (1) NPs contain C6-C3 units in their structures, such as lignans, flavones, coumarins, simple phenylpropanoids, and hybrids between C6-C3 units and other structures. The enantiomerism for those structures presumably derives from the nonstereoselective oxidation of the C3 unit or nonstereoselective coupling of the C6-C3 units, through enzymatic or nonenzymatic reactions. (2) NPs formed by combination of 2~4 isopentenyl units, such as monoterpenoids, sesquiterpenoids, diterpenoids, and meroterpenoids, or having isopentenyl units as side chains, can exist in the form of enantiomers, and need to be further researched. (3) Alkaloid NPs have a variety of structural types, each of which may exist in the form of enantiomers. (4) When NPs with long chain, e.g., fatty acids and diarylheptanoids, have chiral centers, testing whether they are enantiomers or not is necessary. (5) NPs with axial chirality tend to exist as enantiomers in nature.
7.4. Identification of the Presence of Enantiomers
The criteria of enantiomeric presence vary, and a confirmative conclusion should be made based on comprehensive considerations. Ideally, the enantiomeric purity of every NP should be checked, but apparently this is neither economical nor technically feasible. Nevertheless, some general guidelines can still be summarized. Firstly, if a NP belongs to a structural group with strong enantiomeric tendency as listed in this review, such as 8,4′-oxyneolignans, caution is required. Secondly, for a previously undescribed NP, when its [α]D value is very small (e.g., <5) or close to zero, the presence of an enantiomeric mixture should be considered. However, this method is not always fully indicative, as some chiral compounds naturally have low [α]D. For a known NP, regardless of whether the magnitude of [α]D value is big or small, if it obviously deviates from the reported datum, the occurrence of enantiomerism is possible, and the purity of the tested NP should first be guaranteed. Thirdly, ECD measurement can also be used to check the enantiomeric purity of a NP (in case it shows a response in the experiment). A good-quality ECD curve usually looks smooth with clear Cotton effect(s) in the normal wavelength range (mostly 190–400 nm); if not, there is a high probability of enantiomeric presence. The aforementioned empirical knowledge is only based on general cases, and in fact, determination of the presence of enantiomers can be complicated. Notably, when the natural e.e. value of a pair of enantiomers is very high, as in the case of neosecurinane alkaloids [5], the researchers’ level of experience and sensitivity to chirality will make the difference.
7.5. Separation and Differentiation of Enantiomers
The separation (use of different chiral stationary materials) and differentiation (abs. config. assignment of an enantiomeric pair) of natural enantiomers were well documented in the review by Cass and Batista Jr. [10] and will not be included here. However, we do wish to emphasize that no omnipotent separation material and single technique can be applied for the purification and abs. config. determination, respectively, for all types of enantiomers, and any doubt regarding enantiomeric purity deserves further investigation.
7.6. Stereochemistry–Bioactivity Relationship of Enantiomers
As for the stereochemistry–bioactivity relationship (SBR) of natural enantiomers, analyses of the biological data gathered in this review do not provide many meaningful clues, and the relevance between the bioactivity and the chirality (dextroisomer or levoisomer) of a pair of enantiomers seems random and irregular in both enzymatic and cellular level bioassays. Although factors regarding the ‘chirality’ of life systems are well-known (e.g., d-glucose and l-amino acids as primary metabolites), there is still a long way to go before we are able to reveal the secrets of the exact SBR of enantiomers. Nevertheless, some general conclusions can still be reached according to the presently accessible information, similar to what Prof. Mori described for insect pheromones [4]. For a specific bioassay model: (1) One enantiomer is active, while the opposite enantiomer is less or not active, and the mixture of them does not result in any extra effect; (2) Both enantiomers are equally active, and their mixture does not result in any extra effect; (3) Both enantiomers are inactive or active, but their mixture is active or more active, suggestive of a synergistic action; (4) One enantiomer is active, whereas the antipodal enantiomer exhibits antagonistic activity, and thus, their mixture will exert an offset effect. Please note that the aforementioned general rules vary for different bioassays and are thus to be taken on a case-by-case basis, because all NPs are produced by the source organisms for their own use, and not for use by humans; we simply take advantage of their biological properties.
All in all, notwithstanding the rapidly growing number of reports and improving awareness of natural enantiomers in recent years, there are still a number of questions which remain to be answered. Our understanding of this fascinating natural phenomenon is only in its infancy
Here, we would like to say to the NP community that enantiomerism in nature is ubiquitous and vital. We hope that this review will prompt future researchers to routinely ask “Is my natural product enantiomerically pure, and if so, which enantiomer have I obtained?”, and in so doing, to perhaps even alter the methods applied by scientists in the future.
Supplementary Materials
Author Contributions
Conceptualization, H.Z. and R.J.C.; formal analysis, J.-H.Y.; data curation, J.-H.Y. and H.Z.; writing—original draft preparation, J.-H.Y. and Z.-P.Y.; writing—review and editing, H.Z. and R.J.C.; supervision, H.Z.; funding acquisition, H.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by National Natural Science Foundation of China (82073729 and 21807040), Natural Science Foundation of Shandong Province (JQ201721), Innovation Team Project of Jinan Science & Technology Bureau (No. 2018GXRC003).
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Dewick, P.M. Medicinal Natural Products: A Biosynthetic Approach, 3rd ed.; John Wiley & Sons, Ltd.: Chichester, UK, 2009; p. 148. [Google Scholar]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs over the Nearly Four Decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef] [PubMed]
- Finefield, J.M.; Sherman, D.H.; Kreitman, M.; Williams, R.M. Enantiomeric natural products: Occurrence and biogenesis. Angew. Chem. 2012, 51, 4802–4836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mori, K. Bioactive natural products and chirality. Chirality 2011, 23, 449–462. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhu, K.K.; Gao, X.H.; Yue, J.M. Natural occurrence of all eight stereoisomers of a neosecurinane structure from Flueggea virosa. Tetrahedron 2017, 73, 4692–4697. [Google Scholar] [CrossRef]
- Song, X.Q.; Zhu, K.; Yu, J.H.; Zhang, Q.; Zhang, Y.; He, F.; Cheng, Z.Q.; Jiang, C.S.; Bao, J.; Zhang, H. New octadecanoid enantiomers from the whole plants of Plantago depressa. Molecules 2018, 23, 1723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, J.; Yu, J.H.; Zhang, J.S.; Song, X.Q.; Bao, J.; Zhang, H. Chromane enantiomers from the flower buds of Tussilago farfara L. and assignments of their absolute configurations. Chem. Biodivers. 2019, 16, e1800581. [Google Scholar] [CrossRef]
- Yu, J.H.; Zhai, H.J.; Yu, Z.P.; Zhang, Q.Q.; Ge, Y.X.; Zhang, Y.Y.; Jiang, C.S.; Zhang, H. Methyl 2-naphthoates from a traditional Chinese herb Morinda officinalis var. officinalis. Tetrahedron 2019, 75, 3793–3801. [Google Scholar] [CrossRef]
- Song, X.Q.; Yu, S.J.; Zhang, J.S.; Yu, J.H.; Zhang, H. New octadecanoid derivatives from the seeds of Ipomoea nil. Chin. J. Nat. Med. 2019, 17, 303–307. [Google Scholar] [CrossRef]
- Batista, A.N.L.; dos Santos, F.M., Jr.; Batista, J.M., Jr.; Cass, Q.B. Enantiomeric mixtures in natural product chemistry: Separation and absolute configuration assignment. Molecules 2018, 23, 492. [Google Scholar] [CrossRef] [Green Version]
- Runeberg, P.A.; Brusentsev, Y.; Rendon, S.M.K.; Eklund, P.C. Oxidative Transformations of Lignans. Molecules 2019, 24, 300. [Google Scholar] [CrossRef] [Green Version]
- Zalesak, F.; Bon, D.J.Y.D.; Pospisil, J. Lignans and Neolignans: Plant secondary metabolites as a reservoir of biologically active substances. Pharmacol. Res. 2019, 146, 104284. [Google Scholar] [CrossRef] [PubMed]
- Moss, G.P. Nomenclature of lignans and neolignans: (IUPAC recommendations 2000). Pure Appl. Chem. 2000, 72, 1493–1523. [Google Scholar] [CrossRef]
- Xu, R.S. Natural product Chemistry, 2nd ed.; Science Press: Beijing, China, 2004. [Google Scholar]
- Liu, X.; Wang, X.B.; Xie, S.S.; Li, Z.R.; Yang, M.H.; Kong, L.Y.; Kim, D.H.; Park, J.S. Lignans from the root of Paeonia lactiflora and their anti-β-amyloid aggregation activities. Fitoterapia 2015, 103, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Xue, Y.; Liu, J.; Yao, G.; Li, D.; Sun, B.; Zhang, J.; Liu, Y.; Qi, C.; Xiang, M.; et al. (±)-Acortatarinowins A-F, Norlignan, Neolignan, and Lignan Enantiomers from Acorus tatarinowii. J. Nat. Prod. 2015, 78, 2205–2214. [Google Scholar] [CrossRef]
- Jiang, C.; Luo, P.; Zhao, Y.; Hong, J.; Morris-Natschke, S.L.; Xu, J.; Chen, C.H.; Lee, K.H.; Gu, Q. Carolignans from the Aerial Parts of Euphorbia sikkimensis and Their Anti-HIV Activity. J. Nat. Prod. 2016, 79, 578–583. [Google Scholar] [CrossRef] [Green Version]
- Seca, A.M.L.; Silva, A.M.S.; Silvestre, A.J.D.; Cavaleiro, J.A.S.; Domingues, F.M.J.; Pascoal-Neto, C. Phenolic constituents from the core of Kenaf (Hibiscus cannabinus). Phytochemistry 2001, 56, 759–767. [Google Scholar] [CrossRef]
- Zhou, L.; Lou, L.L.; Wang, W.; Lin, B.; Chen, J.N.; Wang, X.B.; Huang, X.X.; Song, S.J. Enantiomeric 8-O-4’ type neolignans from red raspberry as potential inhibitors of β-amyloid aggregation. J. Funct. Foods 2017, 37, 322–329. [Google Scholar] [CrossRef]
- Zhao, P.; Zhang, H.; Han, F.Y.; Guo, R.; Huang, S.W.; Lin, B.; Huang, X.X.; Song, S.J. Chiral resolution and neuroprotective activities of enantiomeric 8-O-4’ neolignans from the fruits of Crataegus pinnatifida Bge. Fitoterapia 2019, 136, 104164. [Google Scholar] [CrossRef]
- Du, Y.Q.; Yan, Z.Y.; Hou, Z.L.; Guo, R.; Bai, M.; Zhou, L.; Lin, B.; Huang, X.X.; Song, S.J. Enantiomeric 8,4’-type oxyneolignans from the root barks of Ailanthus altissima (Mill.) Swingle and their neuroprotective effects against H2O2-induced SH-SY5Y cells injury. Fitoterapia 2019, 139, 104403. [Google Scholar] [CrossRef]
- Xiong, L.; Zhu, C.; Li, Y.; Tian, Y.; Lin, S.; Yuan, S.; Hu, J.; Hou, Q.; Chen, N.; Yang, Y.; et al. Lignans and neolignans from Sinocalamus affinis and their absolute configurations. J. Nat. Prod. 2011, 74, 1188–1200. [Google Scholar] [CrossRef]
- Braga, A.C.H.; Zacchino, S.; Badano, H.; Gonzalez Sierra, M.; Ruveda, E.A. Carbon-13 NMR spectral and conformational analysis of 8-O-4’ neolignans. Phytochemistry 1984, 23, 2025–2028. [Google Scholar] [CrossRef]
- Gan, M.; Zhang, Y.; Lin, S.; Liu, M.; Song, W.; Zi, J.; Yang, Y.; Fan, X.; Shi, J.; Hu, J.; et al. Glycosides from the root of Iodes cirrhosa. J. Nat. Prod. 2008, 71, 647–654. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.N.; Han, B.; Yang, P.F.; Feng, Z.M.; Jiang, J.S.; Zhang, P.C. A concise approach for determining the relative configuration of H-7 and H-8 in 8,4′-oxyneolignans by 1H NMR spectroscopy. Org. Chem. Front. 2019, 6, 886–891. [Google Scholar] [CrossRef]
- Lin, S.; Wang, S.; Liu, M.; Gan, M.; Li, S.; Yang, Y.; Wang, Y.; He, W.; Shi, J. Glycosides from the Stem Bark of Fraxinus sieboldiana. J. Nat. Prod. 2007, 70, 817–823. [Google Scholar] [CrossRef] [PubMed]
- Pescitelli, G.; Bruhn, T. Good Computational Practice in the Assignment of Absolute Configurations by TDDFT Calculations of ECD Spectra. Chirality 2016, 28, 466–474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, D.-P.; Feng, X.L.; Zhang, W.Y.; Gao, H.; Cheng, X.R.; Zhou, W.X.; Yu, Y.; Yao, X.S. Anti-neuroinflammatory asarone derivatives from the rhizomes of Acorus tatarinowii. RSC Adv. 2017, 7, 8512–8520. [Google Scholar] [CrossRef] [Green Version]
- Fang, Y.S.; Cai, L.; Wang, J.P.; Yin, T.P.; Yu, J.; Ding, Z.T.; Fang, Y.S.; Yang, M.H. New phenylpropanoids from Bulbophyllum retusiusculum. Arch. Pharm. Res. 2018, 41, 1074–1081. [Google Scholar] [CrossRef]
- Cheng, Z.B.; Lu, X.; Bao, J.M.; Han, Q.H.; Dong, Z.; Tang, G.H.; Gan, L.S.; Luo, H.B.; Yin, S. (±)-Torreyunlignans A-D, Rare 8-9’ Linked Neolignan Enantiomers as Phosphodiesterase-9A Inhibitors from Torreya yunnanensis. J. Nat. Prod. 2014, 77, 2651–2657. [Google Scholar] [CrossRef]
- Yang, D.T.; Lin, S.S.; Chen, J.H.; Yuan, S.T.; Shi, J.S.; Wang, J.S.; Jia, A.Q. (+)- and (−)-liriodenol, a pair of novel enantiomeric lignans from Liriodendron hybrid. Bioorg. Med. Chem. Lett. 2015, 25, 1976–1978. [Google Scholar] [CrossRef]
- Zhu, Y.; Huang, R.Z.; Wang, C.G.; Ouyang, X.L.; Jing, X.T.; Liang, D.; Wang, H.S. New inhibitors of matrix metalloproteinases 9 (MMP-9): Lignans from Selaginella moellendorffii. Fitoterapia 2018, 130, 281–289. [Google Scholar] [CrossRef]
- Jiao, S.; Su, G.; Zhou, X.; Wuken, S.; Li, J.; Tu, P.; Chai, X. Alashinols I and J, two novel phenols from stem barks of Syringa pinnatifolia. Phytochem. Lett. 2019, 33, 61–63. [Google Scholar] [CrossRef]
- Odonbayar, B.; Murata, T.; Buyankhishig, B.; Sasaki, K.; Suganuma, K.; Ishikawa, Y.; Batkhuu, J. Acylated Lignans Isolated from Brachanthemum gobicum and Their Trypanocidal Activity. J. Nat. Prod. 2019, 82, 774–784. [Google Scholar] [CrossRef] [PubMed]
- Mori, K.; Komatsu, M.; Kido, M.; Nakagawa, K. Synthesis of lignans. II. A simple biogenetic-type synthesis of magnosalicin, a new neolignan with antiallergy activity isolated from Magnolia salicifolia. Tetrahedron 1986, 42, 523–528. [Google Scholar] [CrossRef]
- Muraoka, O.; Sawada, T.; Morimoto, E.; Tanabe, G. Chalcones as synthetic intermediates. A facile route to (±)-magnosalicin, an antiallergy neolignan. Chem. Pharm. Bull. 1993, 41, 772–774. [Google Scholar] [CrossRef] [Green Version]
- Lopes, N.P.; Blumenthal, E.E.d.A.; Cavalheiro, A.J.; Kato, M.J.; Yoshida, M. Lignans, γ-lactones and propiophenones of Virola surinamensis. Phytochemistry 1996, 43, 1089–1092. [Google Scholar] [CrossRef]
- Lu, Y.; Xue, Y.; Chen, S.; Zhu, H.; Wang, J.; Liu, J.; Qi, C.; Zhang, Y.; Lu, Y.; Zhang, J.; et al. Antioxidant Lignans and Neolignans from Acorus tatarinowii. Sci. Rep. 2016, 6, 22909. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.Y.; Cheng, B.; Zheng, Y.J.; Dong, Z.; Lin, S.L.; Tang, G.H.; Gu, Q.; Yin, S. Enantiomeric neolignans and sesquineolignans from Jatropha integerrima and their absolute configurations. RSC Adv. 2015, 5, 12202–12208. [Google Scholar] [CrossRef]
- Lou, L.L.; Yao, G.D.; Wang, J.; Zhao, W.Y.; Wang, X.B.; Huang, X.X.; Song, S.J. Enantiomeric neolignans from Picrasma quassioides exhibit distinctive cytotoxicity on hepatic carcinoma cells through ROS generation and apoptosis induction. Bioorg. Med. Chem. Lett. 2018, 28, 1263–1268. [Google Scholar] [CrossRef]
- Antus, S.; Kurtan, T.; Juhasz, L.; Kiss, L.; Hollosi, M.; Majer, Z. Chiroptical properties of 2,3-dihydrobenzo[b]furan and chromane chromophores in naturally occurring O-heterocycles. Chirality 2001, 13, 493–506. [Google Scholar] [CrossRef]
- Zhou, L.; Yao, G.D.; Lu, L.W.; Song, X.Y.; Lin, B.; Song, S.J.; Wang, X.B.; Huang, X.X. Neolignans from Red Raspberry (Rubus idaeus L.) Exhibit Enantioselective Neuroprotective Effects against H2O2-Induced Oxidative Injury in SH-SY5Y Cells. J. Agric. Food Chem. 2018, 66, 11390–11397. [Google Scholar] [CrossRef]
- Zhou, L.; Xi, Y.F.; Wang, W.; Lin, B.; Wang, X.B.; Huang, X.X.; Song, S.J. Chiral resolution and bioactivity of enantiomeric benzofuran neolignans from the fruit of Rubus ideaus L. Fitoterapia 2018, 127, 56–61. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Zhou, L.; Wu, Y.; Zhu, H.; Hu, Z.; Wang, J.; Luo, Z.; Xue, Y.; Zhang, Y.; Lai, Y.; et al. Enantiomeric Lignans and Neolignans from Phyllanthus glaucus: Enantioseparation and Their Absolute Configurations. Sci. Rep. 2016, 6, 24809. [Google Scholar] [CrossRef] [PubMed]
- Mei, R.Q.; Wang, Y.H.; Du, G.H.; Liu, G.M.; Zhang, L.; Cheng, Y.X. Antioxidant lignans from the fruits of Broussonetia papyrifera. J. Nat. Prod. 2009, 72, 621–625. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Han, F.Y.; Lu, L.W.; Yao, G.D.; Zhang, Y.Y.; Wang, X.B.; Lin, B.; Huang, X.X.; Song, S.J. Isolation of enantiomeric furolactones and furofurans from Rubus idaeus L. with neuroprotective activities. Phytochemistry 2019, 164, 122–129. [Google Scholar] [CrossRef]
- Li, X.N.; Chu, C.; Tong, S.Q.; Cheng, D.P.; Yan, J.Z. A new furolactone-type lignan from Lycium chinense. Nat. Prod. Res. 2013, 27, 750–752. [Google Scholar] [CrossRef]
- Wang, Y.X.; Lin, B.; Zhou, L.; Yan, Z.Y.; Zhang, H.; Huang, X.X.; Song, S.J. Anti-β-amyloid aggregation activity of enantiomeric furolactone-type lignans from Archidendron clypearia (Jack) I.C.N. Nat. Prod. Res. 2020, 34, 456–463. [Google Scholar] [CrossRef]
- Zhou, X.M.; Zheng, C.J.; Wu, J.T.; Chen, G.Y.; Chen, J.; Sun, C.G. Five new lactone derivatives from the stems of Dendrobium nobile. Fitoterapia 2016, 115, 96–100. [Google Scholar] [CrossRef]
- Liu, W.J.; Chen, Y.J.; Chen, D.N.; Wu, Y.P.; Gao, Y.J.; Li, J.; Zhong, W.J.; Jiang, L. A new pair of enantiomeric lignans from the fruits of Morinda citrifolia and their absolute configuration. Nat. Prod. Res. 2018, 32, 933–938. [Google Scholar] [CrossRef]
- Wang, Y.X.; Zhou, L.; Wang, J.; Lin, B.; Wang, X.B.; Huang, X.X.; Song, S.J. Enantiomeric lignans with anti-β-amyloid aggregation activity from the twigs and leaves of Pithecellobium clypearia Benth. Bioorg. Chem. 2018, 77, 579–585. [Google Scholar] [CrossRef]
- Xi, Y.F.; Liu, S.F.; Hong, W.; Song, X.Y.; Lou, L.L.; Zhou, L.; Yao, G.D.; Lin, B.; Wang, X.B.; Huang, X.X.; et al. Discovery of cycloneolignan enantiomers from Isatis indigotica Fortune with neuroprotective effects against MPP+-induced SH-SY5Y cell injury. Bioorg. Chem. 2019, 88, 102926. [Google Scholar] [CrossRef]
- Fukuda, T.; Nagai, K.; Tomoda, H. (±)-Tylopilusins, Diphenolic Metabolites from the Fruiting Bodies of Tylopilus eximius. J. Nat. Prod. 2012, 75, 2228–2231. [Google Scholar] [CrossRef]
- Lai, Y.; Liu, T.; Sa, R.; Wei, X.; Xue, Y.; Wu, Z.; Luo, Z.; Xiang, M.; Zhang, Y.; Yao, G. Neolignans with a Rare 2-Oxaspiro[4.5]deca-6,9-dien-8-one Motif from the Stem Bark of Cinnamomum subavenium. J. Nat. Prod. 2015, 78, 1740–1744. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Liu, Y.; Li, Y.; Li, L.; Qu, J.; Ma, S.; Yu, S. Chiral resolution and absolute configuration of a pair of rare racemic spirodienone sesquineolignans from Xanthium sibiricum. Org. Lett. 2014, 16, 5406–5409. [Google Scholar] [CrossRef] [PubMed]
- Supuran, C.T. Coumarin carbonic anhydrase inhibitors from natural sources. J. Enzyme Inhib. Med. Chem. 2020, 35, 1462–1470. [Google Scholar] [CrossRef] [PubMed]
- Akkol, E.K.; Genc, Y.; Karpuz, B.; Sobarzo-Sanchez, E.; Capasso, R. Coumarins and Coumarin-Related Compounds in Pharmacotherapy of Cancer. Cancers 2020, 12, 1959. [Google Scholar] [CrossRef] [PubMed]
- Hong, M.J.; Kim, J. Determination of the Absolute Configuration of Khellactone Esters from Peucedanum japonicum Roots. J. Nat. Prod. 2017, 80, 1354–1360. [Google Scholar] [CrossRef] [PubMed]
- Hata, K.; Kozawa, M.; Baba, K.; Yen, K.Y.; Yang, L.L. Coumarins from the roots of Angelica morii. Chem. Pharm. Bull. 1974, 22, 957–961. [Google Scholar] [CrossRef] [Green Version]
- Xiong, Y.Y.; Wu, F.H.; Wang, J.S.; Li, J.; Kong, L.Y. Attenuation of airway hyperreactivity and T helper cell type 2 responses by coumarins from Peucedanum praeruptorum Dunn in a murine model of allergic airway inflammation. J. Ethnopharmacol. 2012, 141, 314–321. [Google Scholar] [CrossRef]
- Nielsen, B.E.; Larsen, P.K.; Lemmich, J. Constituents of umbelliferous plants. XVII. Coumarins from Seseli gummiferum. Structure of two new coumarins. Acta Chem. Scand. 1971, 25, 529–533. [Google Scholar] [CrossRef] [Green Version]
- Matsuda, H.; Murakami, T.; Nishida, N.; Kageura, T.; Yoshikawa, M. Medicinal foodstuffs. XX. Vasorelaxant active constituents from the roots of Angelica furcijuga Kitagawa: Structures of hyuganins A, B, C, and D. Chem. Pharm. Bull. 2000, 48, 1429–1435. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.L.; Jing, W.H.; Tu, P.F.; Wang, Y.T. Enantiomeric separation of angular-type pyranocoumarins from Peucedani Radix using AD-RH chiral column. Nat. Prod. Res. 2014, 28, 545–550. [Google Scholar] [CrossRef] [PubMed]
- Górski, B.; Talko, A.; Basak, T.; Barbasiewicz, M. Olefination with Sulfonyl Halides and Esters: Scope, Limitations, and Mechanistic Studies of the Hawkins Reaction. Org. Lett. 2017, 19, 1756–1759. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Wang, S.; Lou, H.; Fan, P. New coumarins and monoterpene galloylglycoside from the stem bark of Sapium baccatum. Fitoterapia 2019, 134, 435–442. [Google Scholar] [CrossRef] [PubMed]
- Su, F.; Zhao, Z.; Ma, S.; Wang, R.; Li, Y.; Liu, Y.; Li, L.; Qu, J.; Yu, S.; Li, Y. Cnidimonins A-C, Three Types of Hybrid Dimer from Cnidium monnieri: Structural Elucidation and Semisynthesis. Org. Lett. 2017, 19, 4920–4923. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.H.; Liu, Y.B.; Ma, S.G.; Li, L.; Li, Y.; Jiang, J.D.; Qu, J.; Yu, S.S. Antiviral Spirotriscoumarins A and B: Two Pairs of Oligomeric Coumarin Enantiomers with a Spirodienone-Sesquiterpene Skeleton from Toddalia asiatica. Org. Lett. 2016, 18, 5146–5149. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Tang, G.H.; Guo, F.L.; Li, W.; Zhang, J.S.; Liu, B.; Yin, S. (P)/(M)-corinepalensin A, a pair of axially chiral prenylated bicoumarin enantiomers with a rare C-5-C-5’ linkage from the twigs of Coriaria nepalensis. Phytochemistry 2018, 149, 140–145. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Lu, S. Biosynthesis and Regulation of Phenylpropanoids in Plants. Crit. Rev. Plant Sci. 2017, 36, 257–290. [Google Scholar] [CrossRef]
- Shi, X.L.; Yan, J.K.; Li, W.K.; Donkor, P.O.; Gao, X.M.; Ding, L.Q.; Qiu, F. Two pairs of phenylpropanoid enantiomers from the leaves of Eucommia ulmoides. J. Asian Nat. Prod. Res. 2018, 20, 1045–1054. [Google Scholar] [CrossRef]
- Baderschneider, B.; Winterhalter, P. Isolation and characterization of novel benzoates, cinnamates, flavonoids, and lignans from Riesling wine and screening for antioxidant activity. J. Agric. Food Chem. 2001, 49, 2788–2798. [Google Scholar] [CrossRef]
- Bai, L.; Wang, Y.; Ge, Y.; Liu, J.; Luan, X. Diastereoselective Synthesis of Dibenzo[b,d]azepines by Pd(II)-Catalyzed [5 + 2] Annulation of o-Arylanilines with Dienes. Org. Lett. 2017, 19, 1734–1737. [Google Scholar] [CrossRef]
- Lu, Y.Y.; Gong, X.P.; Xue, Y.B.; Zhu, H.C.; Li, X.N.; Hu, L.Z.; Guan, J.K.; Zhang, J.W.; Du, G.; Zhang, Y.H. Two pairs of chlorine-containing phenylpropanoid enantiomers from Acorus tatarinowii. Chin. Chem. Lett. 2017, 28, 1460–1464. [Google Scholar] [CrossRef]
- Gao, E.; Zhou, Z.Q.; Zou, J.; Yu, Y.; Feng, X.L.; Chen, G.D.; He, R.R.; Yao, X.S.; Gao, H. Bioactive Asarone-Derived Phenylpropanoids from the Rhizome of Acorus tatarinowii Schott. J. Nat. Prod. 2017, 80, 2923–2929. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Yao, G.D.; Song, X.Y.; Wang, J.; Lin, B.; Wang, X.B.; Huang, X.X.; Song, S.J. Neuroprotective Effects of 1,2-Diarylpropane Type Phenylpropanoid Enantiomers from Red Raspberry against H2O2-Induced Oxidative Stress in Human Neuroblastoma SH-SY5Y Cells. J. Agric. Food Chem. 2018, 66, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Rayanil, K.O.; Nimnoun, C.; Tuntiwachwuttikul, P. New phenolics from the wood of Casearia grewiifolia. Phytochem. Lett. 2012, 5, 59–62. [Google Scholar] [CrossRef]
- Yan, J.K.; Shi, X.L.; Donkor, P.O.; Gao, X.M.; Ding, L.Q.; Qiu, F.; Yan, J.K.; Shi, X.L.; Donkor, P.O.; Gao, X.M.; et al. Two pairs of phenolic enantiomers from the leaves of Eucommia ulmoides Oliver. Nat. Prod. Res. 2019, 33, 1162–1168. [Google Scholar] [CrossRef]
- Guo, R.; Shang, X.Y.; Lv, T.M.; Yao, G.D.; Lin, B.; Wang, X.B.; Huang, X.X.; Song, S.J. Phenylpropanoid derivatives from the fruit of Crataegus pinnatifida Bunge and their distinctive effects on human hepatoma cells. Phytochemistry 2019, 164, 252–261. [Google Scholar] [CrossRef]
- Rosales, P.F.; Bordin, G.S.; Gower, A.E.; Moura, S. Indole alkaloids: 2012 until now, highlighting the new chemical structures and biological activities. Fitoterapia 2020, 143, 104558. [Google Scholar] [CrossRef]
- Xu, D.; Xu, Z. Indole Alkaloids with Potential Anticancer Activity. Curr. Top. Med. Chem. 2020, 20, 1938–1949. [Google Scholar] [CrossRef]
- Xi, Y.F.; Lou, L.L.; Xu, Z.Y.; Hou, Z.L.; Wang, X.B.; Huang, X.X.; Song, S.J. Alkaloid Enantiomers from Isatis tinctoria with Neuroprotective Effects against H2O2-Induced SH-SY5Y Cell Injury. Planta Med. 2019, 85, 1374–1382. [Google Scholar] [CrossRef]
- Chen, M.; Gan, L.; Lin, S.; Wang, X.; Li, L.; Li, Y.; Zhu, C.; Wang, Y.; Jiang, B.; Jiang, J.; et al. Alkaloids from the root of Isatis indigotica. J. Nat. Prod. 2012, 75, 1167–1176. [Google Scholar] [CrossRef]
- Liu, J.; Du, Y.Q.; Li, C.J.; Li, L.; Chen, F.Y.; Yang, J.Z.; Chen, N.H.; Zhang, D.M. Alkaloids from the stems of Clausena lansium and their neuroprotective activity. RSC Adv. 2017, 7, 35417–35425. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Deng, A.J.; Li, L.; Wu, L.Q.; Ji, M.; Zhang, H.J.; Li, Z.H.; Ma, L.; Zhang, Z.H.; Chen, X.G.; et al. Azacyclo-indoles and Phenolics from the Flowers of Juglans regia. J. Nat. Prod. 2017, 80, 2189–2198. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.F.; Lin, B.; Xi, Y.F.; Zhou, L.; Lou, L.L.; Huang, X.X.; Wang, X.B.; Song, S.J. Bioactive spiropyrrolizidine oxindole alkaloid enantiomers from Isatis indigotica Fortune. Org. Biomol. Chem. 2018, 16, 9430–9439. [Google Scholar] [CrossRef]
- Zhang, D.; Shi, Y.; Xu, R.; Du, K.; Guo, F.; Chen, K.; Li, Y.; Wang, R. Alkaloid enantiomers from the roots of Isatis indigotica. Molecules 2019, 24, 3140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Q.; Xu, C.; Chen, M.; Lin, S.; Zhu, C.; Jiang, J.; Yang, Y.; Shi, J.; Chen, M.; Li, Y.; et al. Sulfur-enriched alkaloids from the root of Isatis indigotica. Acta Pharm. Sin. B 2018, 8, 933–943. [Google Scholar] [CrossRef]
- Chen, M.; Lin, S.; Li, L.; Zhu, C.; Wang, X.; Wang, Y.; Jiang, B.; Wang, S.; Li, Y.; Jiang, J.; et al. Enantiomers of an Indole Alkaloid Containing Unusual Dihydrothiopyran and 1,2,4-Thiadiazole Rings from the Root of Isatis indigotica. Org. Lett. 2012, 14, 5668–5671. [Google Scholar] [CrossRef]
- Nge, C.E.; Chong, K.W.; Thomas, N.F.; Lim, S.H.; Low, Y.Y.; Kam, T.S. Ibogan, aspidosperman, vincamine, and bisindole alkaloids from a Malayan Tabernaemontana corymbosa: Iboga alkaloids with C-20α substitution. J. Nat. Prod. 2016, 79, 1388–1399. [Google Scholar] [CrossRef]
- Zhang, D.B.; Yu, D.G.; Sun, M.; Zhu, X.X.; Yao, X.J.; Zhou, S.Y.; Chen, J.J.; Gao, K. Ervatamines A-I, Anti-inflammatory Monoterpenoid Indole Alkaloids with Diverse Skeletons from Ervatamia hainanensis. J. Nat. Prod. 2015, 78, 1253–1261. [Google Scholar] [CrossRef]
- Li, D.W.; Guo, Q.L.; Meng, X.H.; Zhu, C.G.; Xu, C.B.; Shi, J.G. Two pairs of unusual scalemic enantiomers from Isatis indigotica leaves. Chin. Chem. Lett. 2016, 27, 1745–1750. [Google Scholar] [CrossRef]
- Li, Y.H.; Zhang, Y.; Peng, L.Y.; Li, X.N.; Zhao, Q.S.; Li, R.T.; Wu, X.D. (±)-Evodiakine, A Pair of Rearranged Rutaecarpine-Type Alkaloids From Evodia rutaecarpa. Nat. Prod. Bioprospect. 2016, 6, 291–296. [Google Scholar] [CrossRef] [Green Version]
- Geng, C.-A.; Huang, X.Y.; Ma, Y.B.; Hou, B.; Li, T.Z.; Zhang, X.M.; Chen, J.J. (±)-Uncarilins A and B, dimeric isoechinulin-type alkaloids from Uncaria rhynchophylla. J. Nat. Prod. 2017, 80, 959–964. [Google Scholar] [CrossRef] [PubMed]
- Dewick, P.M. Medicinal Natural Products: A Biosynthetic Approach, 3rd ed.; John Wiley & Sons, Ltd.: Chichester, UK, 2009; pp. 380–395. [Google Scholar]
- Shang, X.F.; Morris-Natschke, S.L.; Yang, G.Z.; Liu, Y.Q.; Guo, X.; Xu, X.S.; Goto, M.; Li, J.C.; Zhang, J.Y.; Lee, K.H. Biologically active quinoline and quinazoline alkaloids part II. Med. Res. Rev. 2018, 38, 1614–1660. [Google Scholar] [CrossRef] [PubMed]
- Shang, X.F.; Yang, C.J.; MorrisNatschke, S.L.; Li, J.C.; Yin, X.D.; Liu, Y.Q.; Guo, X.; Peng, J.W.; Goto, M.; Zhang, J.Y.; et al. Biologically active isoquinoline alkaloids covering 2014-2018. Med. Res. Rev. 2020, 40, 2212–2289. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.N.; Guo, X.X.; Liu, S.; Feng, L.; Bi, Q.R.; Wang, Z.; Tan, N.H. (±)-Zanthonitidine A, a Pair of Enantiomeric Furoquinoline Alkaloids from Zanthoxylum nitidum with Antibacterial Activity. Nat. Prod. Bioprospect. 2018, 8, 361–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Wang, X.; Lin, S.; Li, L.; Shi, J.; Chen, M. Three pairs of alkaloid enantiomers from the root of Isatis indigotica. Acta Pharm. Sin. B 2016, 6, 141–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, X.; Bai, R.; Guo, Q.; Su, G.; Wang, J.; Yang, X.; Li, L.; Tu, P.; Chai, X. Hendersine A, a novel isoquinoline alkaloid from Corydalis hendersonii. Tetrahedron Lett. 2016, 57, 4858–4862. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, Q.Y.; Tu, P.F.; Liang, H.; Xu, F.C. Mucroniferanines A-G, Isoquinoline Alkaloids from Corydalis mucronifera. J. Nat. Prod. 2018, 81, 364–370. [Google Scholar] [CrossRef]
- Feng, F.; Ye, F.Z.; Li, C.L.; Liu, W.Y.; Xie, N. Two new benzo phenanthridine isoquinoline alkaloids from Macleaya cordata. Chin. J. Nat. Med. 2012, 10, 378–382. [Google Scholar] [CrossRef]
- Sai, C.M.; Li, D.H.; Li, S.G.; Han, T.; Guo, Y.Z.; Pei, Y.H.; Bai, J.; Jing, Y.K.; Li, Z.L.; Hua, H.M. Racemic alkaloids from Macleaya cordata: Structural elucidation, chiral resolution, and cytotoxic, antibacterial activities. RSC Adv. 2016, 6, 41173–41180. [Google Scholar] [CrossRef]
- Yang, Z.; Liu, Z.; Han, N.; Jiang, B.; Guo, D.; Teng, F.; Wang, Y.; Yin, J. Ambidalmines A-E and ambidimerine F: Bioactive dihydrobenzophenanthridine alkaloids from Corydalis ambigua var. amurensis. Eur. J. Med. Chem. 2014, 84, 417–424. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, S.Y.; Chen, L.; Huang, X.J.; Zhang, Q.W.; Jiang, R.W.; Yao, F.; Ye, W.C. New enantiomeric isoquinoline alkaloids from Coptis chinensis. Phytochem. Lett. 2014, 7, 89–92. [Google Scholar] [CrossRef]
- Sai, C.M.; Li, D.H.; Xue, C.M.; Wang, K.B.; Hu, P.; Pei, Y.H.; Bai, J.; Jing, Y.K.; Li, Z.L.; Hua, H.M. Two Pairs of Enantiomeric Alkaloid Dimers from Macleaya cordata. Org. Lett. 2015, 17, 4102–4105. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.; Dan, W.; Schneider, U.; Wang, J. beta-Carboline alkaloid monomers and dimers: Occurrence, structural diversity, and biological activities. Eur. J. Med. Chem. 2018, 157, 622–656. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Feng, Z.L.; Wang, Y.T.; Lin, L.G. Anticancer carbazole alkaloids and coumarins from Clausena plants: A review. Chin. J. Nat. Med. 2017, 15, 881–888. [Google Scholar] [CrossRef]
- Zhao, W.Y.; Zhou, W.Y.; Chen, J.J.; Yao, G.D.; Lin, B.; Wang, X.B.; Huang, X.X.; Song, S.J. Enantiomeric β-carboline dimers from Picrasma quassioides and their anti-hepatoma potential. Phytochemistry 2019, 159, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.M.; Li, F.; Zheng, F.F.; Gong, N.N.; Li, Y.; Feng, W.Z.; Tian, L. (±)-Quassidine K, a pair of cytotoxic bis-β-carboline alkaloid enantiomers from Picrasma quassioides. Nat. Prod. Res. 2020, 34, 489–493. [Google Scholar] [CrossRef]
- Jiao, W.H.; Chen, G.D.; Gao, H.; Li, J.; Gu, B.B.; Xu, T.T.; Yu, H.B.; Shi, G.H.; Yang, F.; Yao, X.S.; et al. (±)-Quassidines I and J, two pairs of cytotoxic bis-β-carboline alkaloid enantiomers from Picrasma quassioides. J. Nat. Prod. 2014, 77, 2707–2712. [Google Scholar] [CrossRef]
- Wang, K.B.; Li, S.G.; Huang, X.Y.; Li, D.H.; Li, Z.L.; Hua, H.M. (±)-Peharmaline A: A Pair of Rare β-Carboline-Vasicinone Hybrid Alkaloid Enantiomers from Peganum harmala. Eur. J. Org. Chem. 2017, 2017, 1876–1879. [Google Scholar] [CrossRef]
- Liu, Y.; Yu, H.Y.; Xu, H.Z.; Liu, J.J.; Meng, X.G.; Zhou, M.; Ruan, H.L. Alkaloids with Immunosuppressive Activity from the Bark of Pausinystalia yohimbe. J. Nat. Prod. 2018, 81, 1841–1849. [Google Scholar] [CrossRef]
- Cao, N.; Chen, Y.; Ma, X.; Zeng, K.; Zhao, M.; Tu, P.; Li, J.; Jiang, Y. Bioactive carbazole and quinoline alkaloids from Clausena dunniana. Phytochemistry 2018, 151, 1–8. [Google Scholar] [CrossRef]
- Ma, X.; Cao, N.; Zhang, C.; Guo, X.; Zhao, M.; Tu, P.; Jiang, Y. Cytotoxic carbazole alkaloid derivatives from the leaves and stems of Murraya microphylla. Fitoterapia 2018, 127, 334–340. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.L.; Li, J.; Zheng, J.; Gu, X.P.; Ferreira, D.; Zjawiony, J.K.; Zhao, M.B.; Guo, X.Y.; Tu, P.F.; Jiang, Y. LC-MS-guided isolation of insulin-secretion-promoting monoterpenoid carbazole alkaloids from Murraya microphylla. J. Nat. Prod. 2018, 81, 2371–2380. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.B.; Gao, J.; Zou, G.A.; Xin, X.L.; Aisa, H.A. Piperidine Alkaloids with Diverse Skeletons from Anacyclus pyrethrum. J. Nat. Prod. 2018, 81, 1474–1482. [Google Scholar] [CrossRef]
- Chen, Q.B.; Aisa, H.A. Alkaloid constituents from Viola tianschanica. Phytochemistry 2017, 144, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Song, W.W.; Zeng, G.Z.; Peng, W.W.; Chen, K.X.; Tan, N.H. Cytotoxic amides and quinolones from Clausena lansium. Helv. Chim. Acta 2014, 97, 298–305. [Google Scholar] [CrossRef]
- Yu, M.Y.; Qin, X.J.; Peng, X.R.; Wang, X.; Tian, X.X.; Li, Z.R.; Qiu, M.H. Macathiohydantoins B-K, novel thiohydantoin derivatives from Lepidium meyenii. Tetrahedron 2017, 73, 4392–4397. [Google Scholar] [CrossRef]
- Sundaram, G.S.M.; Venkatesh, C.; Ila, H.; Junjappa, H. 1-(Methyldithiocarbonyl)imidazole as thiocarbonyl transfer reagent: A facile one-pot three-component synthesis of 3,5- and 1,3,5-substituted 2-thiohydantoins. Synlett 2007, 2, 251–254. [Google Scholar] [CrossRef]
- Al-Khdhairawi, A.A.Q.; Krishnan, P.; Lim, K.H.; Mai, C.W.; Leong, C.O.; Chung, F.F.L.; Leong, C.O.; Yong, K.T.; Chong, K.W.; Low, Y.Y.; et al. A Bis-benzopyrroloisoquinoline Alkaloid Incorporating a Cyclobutane Core and a Chlorophenanthroindolizidine Alkaloid with Cytotoxic Activity from Ficus fistulosa var. tengerensis. J. Nat. Prod. 2017, 80, 2734–2740. [Google Scholar] [CrossRef]
- Ratnagiriswaran, A.N.; Venkatachalam, K. The chemical examination of Tylophora asthmatica and the isolation of the alkaloids tylophorine and tylophorinine. Indian J. Med. Res. 1935, 22, 433–441. [Google Scholar]
- Stoye, A.; Peez, T.E.; Opatz, T. Left, Right, or Both? On the Configuration of the Phenanthroindolizidine Alkaloid Tylophorine from Tylophora indica. J. Nat. Prod. 2013, 76, 275–278. [Google Scholar] [CrossRef]
- Hu, Y.; Zhang, C.; Zhao, X.; Wang, Y.; Feng, D.; Zhang, M.; Xie, H. (±)-Homocrepidine A, a Pair of Anti-inflammatory Enantiomeric Octahydroindolizine Alkaloid Dimers from Dendrobium crepidatum. J. Nat. Prod. 2016, 79, 252–256. [Google Scholar] [CrossRef] [PubMed]
- Li, S.G.; Wang, K.B.; Gong, C.; Bao, Y.; Qin, N.B.; Li, D.H.; Li, Z.L.; Bai, J.; Hua, H.M. Cytotoxic quinazoline alkaloids from the seeds of Peganum harmala. Bioorg. Med. Chem. Lett. 2018, 28, 103–106. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.D.; Cheng, X.M.; Liu, W.; Han, Z.Z.; Chou, G.X.; Wang, Y.; Sun, D.X.; Wang, Z.T.; Wang, C.H. Peganumine B-I and two enantiomers: New alkaloids from the seeds of Peganum harmala Linn. and their potential cytotoxicity and cholinesterase inhibitory activities. RSC Adv. 2016, 6, 15976–15987. [Google Scholar] [CrossRef]
- Shou, Q.; Banbury, L.K.; Renshaw, D.E.; Smith, J.E.; He, X.; Dowell, A.; Griesser, H.J.; Heinrich, M.; Wohlmuth, H. Parvifloranines A and B, Two 11-Carbon Alkaloids from Geijera parviflora. J. Nat. Prod. 2013, 76, 1384–1387. [Google Scholar] [CrossRef]
- Xia, G.Y.; Owusu, D.P.; Ding, L.Q.; Qiu, F.; Xia, G.Y.; Sun, D.J.; Ma, J.H.; Liu, Y.; Chen, L.X.; Zhao, F. (+)/(−)-Phaeocaulin A-D, four pairs of new enantiomeric germacrane-type sesquiterpenes from Curcuma phaeocaulis as natural nitric oxide inhibitors. Sci. Rep. 2017, 7, 43576. [Google Scholar] [CrossRef]
- Han, Q.T.; Li, G.S.; Xiang, K.L.; Ren, Y.; Dai, S.J. Flavonoid alkaloids from Scutellaria moniliorrhiza with anti-inflammatory activities and inhibitory activities against aldose reductase. Phytochemistry 2018, 152, 91–96. [Google Scholar] [CrossRef]
- Zhou, D.; Chen, G.; Ma, Y.P.; Wang, C.G.; Lin, B.; Yang, Y.Q.; Li, W.; Koike, K.; Hou, Y.; Li, N. Isolation, structural elucidation, optical resolution, and antineuroinflammatory activity of phenanthrene and 9,10-dihydrophenanthrene derivatives from Bletilla striata. J. Nat. Prod. 2019, 82, 2238–2245. [Google Scholar] [CrossRef]
- Cheng, Z.Y.; Du, Y.Q.; Zhang, Q.; Lin, B.; Gao, P.Y.; Huang, X.X.; Song, S.J. Two pairs of new alkaloid enantiomers with a spiro [benzofuranone-benzazepine] skeleton from the bark of Juglans mandshurica. Tetrahedron Lett. 2018, 59, 2050–2053. [Google Scholar] [CrossRef]
- Chen, F.; Huang, X.J.; Liang, Q.P.; Huang, Y.P.; Lan, T.; Zhou, G.X. Three new lignanamides from the root of Lycium chinense with anti-inflammatory activity. Nat. Prod. Res. 2019, 33, 3378–3382. [Google Scholar] [CrossRef]
- Zhu, G.Y.; Yang, J.; Yao, X.J.; Yang, X.; Fu, J.; Liu, X.; Bai, L.P.; Liu, L.; Jiang, Z.H. (±)-Sativamides A and B, two pairs of racemic nor-lignanamide enantiomers from the fruits of Cannabis sativa. J. Org. Chem. 2018, 83, 2376–2381. [Google Scholar] [CrossRef]
- Azmi, M.N.; Chan, G.; Peresse, T.; Remeur, C.; Roussi, F.; Litaudon, M.; Awang, K. Kingianins O-Q: Pentacyclic polyketides from Endiandra kingiana as inhibitor of Mcl-1/Bid interaction. Fitoterapia 2016, 109, 190–195. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Song, J.; Liu, A.; Xiao, B.; Li, S.; Wen, Z.; Lu, Y.; Du, G. Research Progress of the Antiviral Bioactivities of Natural Flavonoids. Nat. Prod. Bioprospect. 2020, 10, 271–283. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Xu, P.S.; Xu, K.P.; Zou, Z.X.; Zhou, G.; Li, D.; Li, D.; Li, X.M.; Li, J.; Tan, G.S. Two new biflavanoids from Selaginella trichoclada Alsto. Nat. Prod. Res. 2021, 35, 3410–3416. [Google Scholar] [CrossRef] [PubMed]
- Gaffield, W. Circular dichroism, optical rotatory dispersion, and absolute configuration of flavanones, 3-hydroxyflavanones, and their glycosides. Determination of aglycone chirality in flavanone glycosides. Tetrahedron 1970, 26, 4093–4108. [Google Scholar] [CrossRef]
- Zhang, T.Y.; Lin, J.B.; Li, Q.Z.; Kang, J.C.; Pan, J.L.; Hou, S.H.; Chen, C.; Zhang, S.Y. Copper-Catalyzed Selective ortho-C–H/N–H Annulation of Benzamides with Arynes: Synthesis of Phenanthridinone Alkaloids. Org. Lett. 2017, 19, 1764–1767. [Google Scholar] [CrossRef]
- Li, R.; Cheng, J.; Jiao, M.; Guo, C.; Chen, S.; Li, L.; Liu, A. New phenylpropanoid-substituted flavan-3-ols and flavonols from the leaves of Uncaria rhynchophylla. Fitoterapia 2017, 116, 17–23. [Google Scholar] [CrossRef]
- Zaki, M.A.; Hetta, M.H.; Mohammed, R.; Nanayakkara, N.P.D.; Jacob, M.R.; Khan, S.I.; Ibrahim, M.A.; Samoylenko, V.; Coleman, C.; Ferreira, D.; et al. Bioactive Formylated Flavonoids from Eugenia rigida: Isolation, Synthesis, and X-ray Crystallography. J. Nat. Prod. 2016, 79, 2341–2349. [Google Scholar] [CrossRef]
- Li, Y.; Qin, X.B.; Liu, H.X.; Xu, Z.F.; Tan, H.B.; Qiu, S.X. Two pairs of enantiomeric propylated flavonoids and a new lignan from the aerial parts of Abrus precatorius. Fitoterapia 2019, 133, 125–129. [Google Scholar] [CrossRef]
- He, Q.F.; Wu, Z.L.; Huang, X.J.; Zhong, Y.L.; Jiang, R.W.; Li, Y.L.; Ye, W.C.; Wang, Y.; He, Q.-F.; Wu, Z.L.; et al. Cajanusflavanols A-C, Three Pairs of Flavonostilbene Enantiomers from Cajanus cajan. Org. Lett. 2018, 20, 876–879. [Google Scholar] [CrossRef]
- Xu, L.; Huang, T.; Huang, C.; Wu, C.; Jia, A.; Hu, X. Chiral separation, absolute configuration, and bioactivity of two pairs of flavonoid enantiomers from Morus nigra. Phytochemistry 2019, 163, 33–37. [Google Scholar] [CrossRef]
- Xu, L.J.; Huang, C.Y.; Niu, L.X.; Wu, C.Z.; Yang, P.M.; Yu, M.H.; Wang, Y.F.; Hu, X. Isoprenylated flavonoids from Morus nigra and their PPAR γ agonistic activities. Fitoterapia 2018, 127, 109–114. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.Q.; Liu, Q.X.; Li, H.L.; Han, W. Flavane constituents from branch of Celastrus hindsii. Zhongcaoyao 2014, 45, 2132–2135. [Google Scholar]
- Li, F.F.; Sun, Q.; Wang, D.; Liu, S.; Lin, B.; Liu, C.T.; Li, L.Z.; Huang, X.X.; Song, S.J. Chiral Separation of Cytotoxic Flavan Derivatives from Daphne giraldii. J. Nat. Prod. 2016, 79, 2236–2242. [Google Scholar] [CrossRef] [PubMed]
- Pang, D.R.; Su, X.Q.; Sun, J.; Li, Y.T.; Zhu, Z.X.; Song, Y.L.; Zhao, Y.F.; Tu, P.F.; Zheng, J.; Li, J. Flavonoid dimers from the total phenolic extract of Chinese dragon’s blood, the red resin of Dracaena cochinchinensis. Fitoterapia 2016, 115, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Hiep, N.T.; Lee, D.; Hiep, N.T.; Kwon, J.; Hong, S.; Mar, W.; Kim, N.; Guo, Y.; Hwang, B.Y. Enantiomeric Isoflavones with neuroprotective activities from the Fruits of Maclura tricuspidata. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef]
- Liu, Z.; Zheng, X.; Wang, Y.; Tang, M.; Chen, S.; Zhang, F.; Li, L.; Zhang, C.; Sun, Y. Lignans and isoflavonoids from the stems of Pisonia umbellifera. RSC Adv. 2018, 8, 16383–16391. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.; Wu, P.; Shen, F.; Ji, J.; Rakesh, K.P. Chalcone derivatives and their antibacterial activities: Current development. Bioorg. Chem. 2019, 91, 103133. [Google Scholar] [CrossRef]
- Shi, Y.S.; Hu, W.Z.; Zhang, X.F.; Lv, X.; Shi, Y.S.; Zhang, Y.; Fu, X. Dihydrochalcones and Diterpenoids from Pteris ensiformis and Their Bioactivities. Molecules 2017, 22, 1413. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Zhang, X.; Kelsang, N.; Tu, G.; Kong, D.; Lu, J.; Zhang, Y.; Liang, H.; Tu, P.; Zhang, Q. Structurally diverse cytotoxic dimeric chalcones from Oxytropis chiliophylla. J. Nat. Prod. 2018, 81, 307–315. [Google Scholar] [CrossRef]
- Simard, F.; Gauthier, C.; Chiasson, E.; Lavoie, S.; Mshvildadze, V.; Legault, J.; Pichette, A. Antibacterial balsacones J-M, hydroxycinnamoylated dihydrochalcones from Populus balsamifera buds. J. Nat. Prod. 2015, 78, 1147–1153. [Google Scholar] [CrossRef]
- Ma, Q.; Min, K.; Li, H.L.; Jiang, J.H.; Liu, Y.; Zhan, R.; Chen, Y.G. Horsfiequinones A-F, Dimeric Diarylpropanoids from Horsfieldia tetratepala. Planta Med. 2014, 80, 688–694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Pogam, P.; Boustie, J. Xanthones of Lichen Source: A 2016 Update. Molecules 2016, 21, 294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, C.; Gong, C.; Pu, J.; Li, D.; Li, Z.; Hua, H.; Jia, C.; Jia, C.; Gong, C.; Pu, J.; et al. A pair of new enantiomers of xanthones from the stems and leaves of Cratoxylum cochinchinense. Chin. Med. 2019, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macabeo, A.P.G.; Martinez, F.P.A.; Kurtan, T.; Toth, L.; Mandi, A.; Schmidt, S.; Heilmann, J.; Alejandro, G.J.D.; Knorn, M.; Dahse, H.-M.; et al. Tetrahydroxanthene-1,3(2H)-dione derivatives from Uvaria valderramensis. J. Nat. Prod. 2014, 77, 2717–2721. [Google Scholar] [CrossRef] [PubMed]
- Boonnak, N.; Chantrapromma, S.; Fun, H.K.; Yuenyongsawad, S.; Patrick, B.O.; Maneerat, W.; Williams, D.E.; Andersen, R.J. Three types of cytotoxic natural caged-scaffolds: Pure enantiomers or partial racemates. J. Nat. Prod. 2014, 77, 1562–1571. [Google Scholar] [CrossRef] [PubMed]
- Niu, S.L.; Li, D.H.; Li, X.Y.; Wang, Y.T.; Li, S.G.; Bai, J.; Pei, Y.H.; Jing, Y.K.; Li, Z.L.; Hua, H.M. Bioassay- and chemistry-guided isolation of scalemic caged prenylxanthones from the leaves of Garcinia bracteata. J. Nat. Prod. 2018, 81, 749–757. [Google Scholar] [CrossRef]
- Sriyatep, T.; Andersen, R.J.; Patrick, B.O.; Pyne, S.G.; Muanprasat, C.; Seemakhan, S.; Borwornpinyo, S.; Laphookhieo, S. Scalemic Caged Xanthones Isolated from the Stem Bark Extract of Garcinia propinqua. J. Nat. Prod. 2017, 80, 1658–1667. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.F.; Liu, Z.P.; Wang, F.P. Natural Sesquiterpenoids as Cytotoxic Anticancer Agents. Mini-Rev. Med. Chem. 2011, 11, 1153–1164. [Google Scholar] [CrossRef]
- Xi, F.M.; Ma, S.-G.; Liu, Y.-B.; Li, L.; Yu, S.-S. Artaboterpenoids A and B, Bisabolene-Derived Sesquiterpenoids from Artabotrys hexapetalus. Org. Lett. 2016, 18, 3374–3377. [Google Scholar] [CrossRef]
- Yan, J.; Shi, X.; Donkor, P.O.; Qiu, F.; Yan, J.; Shi, X.; Donkor, P.O.; Gao, X.; Ding, L.; Qiu, F.; et al. Nine pairs of megastigmane enantiomers from the leaves of Eucommia ulmoides Oliver. J. Nat. Med. 2017, 71, 780–790. [Google Scholar] [CrossRef]
- Zhang, R.; Feng, X.; Su, G.; Mu, Z.; Zhang, H.; Zhao, Y.; Jiao, S.; Cao, L.; Chen, S.; Tu, P.; et al. Bioactive Sesquiterpenoids from the Peeled Stems of Syringa pinnatifolia. J. Nat. Prod. 2018, 81, 1711–1720. [Google Scholar] [CrossRef] [PubMed]
- Ge, C.Y.; Zhang, J.L.; Ge, C.Y.; Zhang, J.L. Bioactive sesquiterpenoids and steroids from the resinous exudates of Commiphora myrrha. Nat. Prod. Res. 2019, 33, 309–315. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.Y.; Song, Y.N.; Zhang, L.J.; Zhang, M.; Ye, Y.; Zhang, H.; Zhu, J.Y.; Luo, L.; Xia, J.; Rahman, K. Sesquiterpenes and lignans from the flower buds of Daphne genkwa and their nitric oxide inhibitory activities. Nat. Prod. Res. 2018, 32, 2893–2899. [Google Scholar] [CrossRef] [PubMed]
- Dong, K.; Pu, J.X.; Du, X.; Li, X.N.; Sun, H.D. Two new guaianolide-type sesquiterpenoids from Kadsura interior. Chin. Chem. Lett. 2013, 24, 111–113. [Google Scholar] [CrossRef]
- Li, H.; Jiao, R.; Mu, J.; Xu, S.; Li, X.; Wang, X.; Li, Z.; Xu, J.; Hua, H.; Li, D. Bioactive Natural Spirolactone-Type 6,7-seco-ent-Kaurane Diterpenoids and Synthetic Derivatives. Molecules 2018, 23, 2914. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.Y.; Gao, K.; Yue, J.M.; Zhao, J.X.; Sheng, L.; Fan, Y.Y.; Li, J.Y.; Yue, J.M. Mangelonoids A and B, Two Pairs of Macrocyclic Diterpenoid Enantiomers from Croton mangelong. Org. Lett. 2018, 20, 4040–4043. [Google Scholar] [CrossRef]
- Feng, L.; Mandi, A.; Tang, C.; Kurtan, T.; Tang, S.; Ke, C.Q.; Shen, N.; Lin, G.; Yao, S.; Ye, Y. A Pair of Enantiomeric Bis-seco-abietane Diterpenoids from Cryptomeria fortunei. J. Nat. Prod. 2018, 81, 2667–2672. [Google Scholar] [CrossRef]
- Jiang, Y.J.; Zhang, Y.; He, J.; Wu, X.D.; Shao, L.D.; Li, X.N.; Su, J.; Peng, L.Y.; Li, R.T.; Zhao, Q.-S. (±)-Salviaprione, a pair of unprecedented abietane-type diterpenoids from Salvia prionitis. Tetrahedron Lett. 2015, 56, 5457–5459. [Google Scholar] [CrossRef]
- Geng, C.A.; Chen, X.L.; Zhou, N.J.; Chen, H.; Ma, Y.B.; Huang, X.Y.; Zhang, X.M.; Chen, J.J. LC-MS Guided Isolation of (±)-Sweriledugenin A, a Pair of Enantiomeric Lactones from Swertia leducii. Org. Lett. 2014, 16, 370–373. [Google Scholar] [CrossRef]
- Liang, W.J.; Geng, C.A.; Zhang, X.M.; Chen, H.; Yang, C.Y.; Rong, G.Q.; Zhao, Y.; Xu, H.B.; Wang, H.; Zhou, N.J.; et al. (±)-Paeoveitol, a Pair of New Norditerpene Enantiomers from Paeonia veitchii. Org. Lett. 2014, 16, 424–427. [Google Scholar] [CrossRef]
- Hu, L.; Zhu, H.; Li, L.; Huang, J.; Sun, W.; Liu, J.; Li, H.; Luo, Z.; Wang, J.; Xue, Y.; et al. (±)-Japonones A and B, two pairs of new enantiomers with anti-KSHV activities from Hypericum japonicum. Sci. Rep. 2016, 6, 27588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, N.; Kobayashi, J.i. Prenylated acylphloroglucinols and meroterpenoids from Hypericum plants. Heterocycles 2015, 90, 23–40. [Google Scholar]
- Huang, G.H.; Hu, Z.; Lei, C.; Wang, P.-P.; Yang, J.; Li, J.Y.; Li, J.; Hou, A.J. Enantiomeric Pairs of Meroterpenoids with Diverse Heterocyclic Systems from Rhododendron nyingchiense. J. Nat. Prod. 2018, 81, 1810–1818. [Google Scholar] [CrossRef] [PubMed]
- Liao, H.B.; Huang, G.H.; Yu, M.H.; Lei, C.; Hou, A.J. Five Pairs of Meroterpenoid Enantiomers from Rhododendron capitatum. J. Org. Chem. 2017, 82, 1632–1637. [Google Scholar] [CrossRef] [PubMed]
- Liao, H.B.; Lei, C.; Gao, L.X.; Li, J.Y.; Li, J.; Hou, A.J. Two Enantiomeric Pairs of Meroterpenoids from Rhododendron capitatum. Org. Lett. 2015, 17, 5040–5043. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Li, L.; Zhu, R.; Zhang, J.; Zhou, J.; Lou, H. Bibenzyl-based meroterpenoid enantiomers from the Chinese liverwort Radula sumatrana. J. Nat. Prod. 2017, 80, 3143–3150. [Google Scholar] [CrossRef]
- Li, C.; Li, C.J.; Ma, J.; Huang, J.W.; Wang, X.Y.; Wang, X.-L.; Ye, F.; Zhang, D.M. Magmenthanes A-H: Eight new meroterpenoids from the bark of Magnolia officinalis var. Biloba. Bioorg. Chem. 2019, 88, 102948. [Google Scholar] [CrossRef]
- Bridi, H.; Meirelles, G.d.C.; von Poser, G.L. Structural diversity and biological activities of phloroglucinol derivatives from Hypericum species. Phytochemistry 2018, 155, 203–232. [Google Scholar] [CrossRef]
- Cheng, M.J.; Yang, X.Y.; Cao, J.Q.; Liu, C.; Zhong, L.P.; Wang, Y.; You, X.F.; Li, C.C.; Wang, L.; Ye, W.C. Isolation, structure elucidation, and total synthesis of myrtuspirone A from Myrtus communis. Org. Lett. 2019, 21, 1583–1587. [Google Scholar] [CrossRef]
- Tantapakul, C.; Maneerat, W.; Sripisut, T.; Ritthiwigrom, T.; Andersen, R.J.; Cheng, P.; Cheenpracha, S.; Raksat, A.; Laphookhieo, S. New Benzophenones and Xanthones from Cratoxylum sumatranum ssp. neriifolium and Their Antibacterial and Antioxidant Activities. J. Agric. Food Chem. 2016, 64, 8755–8762. [Google Scholar] [CrossRef]
- Hu, L.; Xue, Y.; Zhang, J.; Zhu, H.; Chen, C.; Li, X.-N.; Liu, J.; Wang, Z.; Zhang, Y.; Zhang, Y. (±)-Japonicols A-D, Acylphloroglucinol-Based Meroterpenoid Enantiomers with Anti-KSHV Activities from Hypericum japonicum. J. Nat. Prod. 2016, 79, 1322–1328. [Google Scholar] [CrossRef] [PubMed]
- Cheenpracha, S.; Pyne, S.G.; Patrick, B.O.; Andersen, R.J.; Maneerat, W.; Laphookhieo, S. Mallopenins A-E, antibacterial phenolic derivatives from the fruits of Mallotus philippensis. J. Nat. Prod. 2019, 82, 2174–2180. [Google Scholar] [CrossRef] [PubMed]
- Hans, M.; Charpentier, M.; Huch, V.; Jauch, J.; Bruhn, T.; Bringmann, G.; Quandt, D. Stereoisomeric Composition of Natural Myrtucommulone A. J. Nat. Prod. 2015, 78, 2381–2389. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Tian, H.Y.; Huang, X.L.; Wang, W.J.; Li, N.P.; He, J.; Ye, W.C.; Wang, L. Xanthchrysones A-C: Rearranged Phenylpropanoyl-Phloroglucinol Dimers with Unusual Skeletons from Xanthostemon chrysanthus. J. Org. Chem. 2019, 84, 15355–15361. [Google Scholar] [CrossRef] [PubMed]
- Su, J.C.; Wang, S.; Cheng, W.; Huang, X.J.; Li, M.M.; Jiang, R.W.; Li, Y.L.; Wang, L.; Ye, W.C.; Wang, Y. Phloroglucinol Derivatives with Unusual Skeletons from Cleistocalyx operculatus and Their in Vitro Antiviral Activity. J. Org. Chem. 2018, 83, 8522–8532. [Google Scholar] [CrossRef] [PubMed]
- Oya, A.; Tanaka, N.; Kusama, T.; Kim, S.Y.; Hayashi, S.; Kojoma, M.; Hishida, A.; Kawahara, N.; Sakai, K.; Gonoi, T.; et al. Prenylated Benzophenones from Triadenum japonicum. J. Nat. Prod. 2015, 78, 258–264. [Google Scholar] [CrossRef]
- Delle Monache, F.; Delle Monache, G.; Pinheiro, R.M.; Radics, L. Chemistry of Clusia genus. Part 3. Nemorosonol, a derivative of tricyclo-[4.3.1.03,7]-decane-7-hydroxy-2,9-dione from Clusia nemorosa. Phytochemistry 1988, 27, 2305–2308. [Google Scholar] [CrossRef]
- Fan, Y.-M.; Yi, P.; Li, Y.; Yan, C.; Huang, T.; Gu, W.; Ma, Y.; Huang, L.J.; Zhang, J.X.; Yang, C.L.; et al. Two Unusual Polycyclic Polyprenylated Acylphloroglucinols, Including a Pair of Enantiomers from Garcinia multiflora. Org. Lett. 2015, 17, 2066–2069. [Google Scholar] [CrossRef]
- Tian, D.S.; Yi, P.; Xia, L.; Xiao, X.; Fan, Y.M.; Gu, W.; Huang, L.J.; Ben-David, Y.; Di, Y.T.; Yuan, C.M.; et al. Garmultins A-G, biogenetically related polycyclic Acylphloroglucinols from Garcinia multiflora. Org. Lett. 2016, 18, 5904–5907. [Google Scholar] [CrossRef]
- Li, Q.; Deng, A.J.; Qin, H.L.; Ji, M.; Li, Z.H.; Chen, X.G. Racemic 3,4-dihydro-4-naphthyl-naphthalen-1(2H)-ones from Juglans regia flowers. Fitoterapia 2019, 139, 104401. [Google Scholar] [CrossRef]
- Zhao, S.M.; Wang, Z.; Chen, X.Q.; Huang, M.B.; Tan, N.H. (±)-Rubioncolin D, a pair of enantiomeric naphthohydroquinone dimers from Rubia oncotricha. Tetrahedron Lett. 2017, 58, 3041–3043. [Google Scholar] [CrossRef]
- Zhao, S.M.; Wang, Z.; Zeng, G.Z.; Song, W.W.; Chen, X.Q.; Li, X.N.; Tan, N.H. New cytotoxic naphthohydroquinone dimers from Rubia alata. Org. Lett. 2014, 16, 5576–5579. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Xie, L.Z.; Li, J.; Chen, G.D.; Aisa, H.A. A pair of new tetrahydro-naphthalenone enantiomers from Eremurus altaicus (Pall.). Stev. Phytochem. Lett. 2015, 13, 330–333. [Google Scholar] [CrossRef]
- Qiao, M.M.; Liu, F.; Liu, Y.; Guo, L.; Zhou, Q.M.; Peng, C.; Xiong, L. Curcumane C and (±)-curcumane D, an unusual seco-cadinane sesquiterpenoid and a pair of unusual nor-bisabolane enantiomers with significant vasorelaxant activity from Curcuma longa. Bioorg. Chem. 2019, 92, 103275. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Zheng, R.R.; Liu, Z.W.; Wang, W.J.; Li, G.Q.; Fan, C.L.; Zhang, X.Q.; Ye, W.C.; Che, C.T. Enantiomeric chromones from Harrisonia perforata. Phytochem. Lett. 2014, 10, 295–299. [Google Scholar] [CrossRef]
- Yuan, W.J.; Gao, W.F.; Zhang, J.H.; Cao, P.; Zhang, Y.; Chen, D.Z.; Li, S.L.; Di, Y.T.; Hao, X.J. (±)-Perforison A, A Pair of New Chromone Enantiomers from Harrisonia perforata. Nat. Prod. Commun. 2017, 12, 63–65. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.F.; Han, C.; Xu, Q.-Q.; Wang, X.B.; Zhao, H.J.; Xue, G.M.; Luo, J.G.; Kong, L.Y. Isolation, chiral-phase resolution, and determination of the absolute configurations of a complete series of stereoisomers of a rearranged acetophenone with three stereocenters. J. Nat. Prod. 2019, 82, 1399–1404. [Google Scholar] [CrossRef]
- Tang, Y.Q.; Li, Y.Q.; Xie, Y.B.; Zhang, J.S.; Li, W.; Lou, L.L.; Zhang, G.; Yin, S. Evodialones A and B: Polyprenylated Acylcyclopentanone Racemates with a 3-Ethyl-1,1-diisopentyl-4-methylcyclopentane Skeleton from Evodia lepta. J. Nat. Prod. 2018, 81, 1483–1487. [Google Scholar] [CrossRef]
- Li, W.; Rao, L.; Liu, Y.; He, Q.; Fan, Y.; You, Y.-X.; Su, Y.; Hu, F.; Xu, Y.K.; Lin, B.; et al. (±)-Meliviticines A and B: Rearranged prenylated acetophenone derivatives from Melicope viticina and their antimicrobial activity. Bioorg. Chem. 2019, 90, 103099. [Google Scholar] [CrossRef]
- Wang, W.J.; Wang, L.; Huang, X.J.; Jiang, R.W.; Yang, X.L.; Zhang, D.M.; Chen, W.M.; Tang, B.Q.; Wang, Y.; Zhang, X.Q.; et al. Two pairs of new benzofuran enantiomers with unusual skeletons from Eupatorium chinense. Tetrahedron Lett. 2013, 54, 3321–3324. [Google Scholar] [CrossRef]
- Xu, J.F.; Zhao, H.J.; Wang, X.B.; Li, Z.R.; Luo, J.; Yang, M.H.; Yang, L.; Yu, W.Y.; Yao, H.Q.; Luo, J.G.; et al. (±)-Melicolones A and B, rearranged prenylated acetophenone stereoisomers with an unusual 9-oxatricyclo[3.2.1.13,8]nonane core from the leaves of Melicope ptelefolia. Org. Lett. 2015, 17, 146–149. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.J.; Zhu, L.J.; Zhao, Y.Q.; Zhen, Y.Q.; Zhang, L.; Lin, C.C.; Chen, L.X. Diarylheptanoid: A privileged structure in drug discovery. Fitoterapia 2020, 142. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wu, Z.L.; Huang, X.J.; Peng, Y.; Huang, X.; Shi, L.; Wang, Y.; Ye, W.C. Evaluation of diarylheptanoid-terpene adduct enantiomers from Alpinia officinarum for neuroprotective activities. J. Nat. Prod. 2018, 81, 162–170. [Google Scholar] [CrossRef] [PubMed]
- Dong, S.H.; Nikolic, D.; Simmler, C.; Qiu, F.; van Breemen, R.B.; Soejarto, D.D.; Pauli, G.F.; Chen, S.N. Diarylheptanoids from Dioscorea villosa (Wild Yam). J. Nat. Prod. 2012, 75, 2168–2177. [Google Scholar] [CrossRef] [Green Version]
- Mondal, S.; Verma, A.; Saha, S. Conformationally Restricted Triarylmethanes: Synthesis, Photophysical Studies, and Applications. Eur. J. Org. Chem. 2019, 2019, 864–894. [Google Scholar] [CrossRef]
- Zhou, B.; Yuan, X.J.; Li, J.Y.; Xu, Y.C.; Li, J.; Yue, J.M.; Liu, D.X.; Li, Y. (−)- and (+)-Securidanes A and B, Natural Triarylmethane Enantiomers: Structure and Bioinspired Total Synthesis. Research 2018, 2018, 2674182. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Yao, Y.; Huang, X.J.; Oberer, L.; Wagner, T.; Guo, J.M.; Gu, W.; Liu, W.D.; Lv, G.X.; Shen, Y.N.; et al. Four new selaginellin derivatives from Selaginella pulvinata: Mechanism of racemization process in selaginellins with quinone methide. Tetrahedron 2015, 71, 1581–1587. [Google Scholar] [CrossRef]
- Zou, J.; Chen, G.-D.; Zhao, H.; Huang, Y.; Luo, X.; Xu, W.; He, R.-R.; Hu, D.; Yao, X.S.; Gao, H. Triligustilides A and B: Two Pairs of Phthalide Trimers from Angelica sinensis with a Complex Polycyclic Skeleton and Their Activities. Org. Lett. 2018, 20, 884–887. [Google Scholar] [CrossRef]
- Mattio, L.M.; Catinella, G.; Pinto, A.; Dallavalle, S. Natural and nature-inspired stilbenoids as antiviral agents. Eur. J. Med. Chem. 2020, 202, 112541. [Google Scholar] [CrossRef]
- Li, X.L.; Zhao, B.X.; Huang, X.J.; Zhang, D.M.; Jiang, R.W.; Li, Y.J.; Jian, Y.Q.; Wang, Y.; Li, Y.L.; Ye, W.C. (+)- and (-)-Cajanusine, a Pair of New Enantiomeric Stilbene Dimers with a New Skeleton from the Leaves of Cajanus cajan. Org. Lett. 2014, 16, 224–227. [Google Scholar] [CrossRef]
- Ito, T.; Endo, H.; Shinohara, H.; Oyama, M.; Akao, Y.; Iinuma, M. Occurrence of stilbene oligomers in Cyperus rhizomes. Fitoterapia 2012, 83, 1420–1429. [Google Scholar] [CrossRef] [PubMed]
- Ito, T.; Endo, H.; Oyama, M.; Iinuma, M. Novel isolation of stilbenoids with enantiomeric and meso forms from a cyperus rhizome. Phytochem. Lett. 2012, 5, 267–270. [Google Scholar] [CrossRef]
- Zhang, X.; Feng, Z.M.; Yang, Y.N.; Jiang, J.S.; Zhang, P.C. Phenolic acid derivatives from Ligusticum chuanxiong. Phytochem. Lett. 2019, 33, 114–118. [Google Scholar] [CrossRef]
- Gao, Y.P.; Shen, Y.H.; Zhang, S.D.; Tian, J.M.; Zeng, H.W.; Ye, J.; Li, H.L.; Shan, L.; Zhang, W.-D. Incarvilleatone, a New Cyclohexylethanoid Dimer from Incarvillea younghusbandii and Its Inhibition against Nitric Oxide (NO) Release. Org. Lett. 2012, 14, 1954–1957. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.M.; Zheng, C.J.; Chen, G.Y.; Zhang, X.P.; Song, X.P.; Li, G.N.; Gan, L.S.; Sun, C.G. Bioactive Phenanthrene and Bibenzyl Derivatives from the Stems of Dendrobium nobile. J. Nat. Prod. 2016, 79, 1791–1797. [Google Scholar] [CrossRef]
- Malebo, H.M.; Kihampa, C.; Mgina, C.A.; Sung’hwa, F.; Jonker, S.A.; Nkunya, M.H.H.; Waibel, R. Antifungal Enantiomeric Styrylpyrones from Sanrafaelia ruffonammari and Ophrypetalum odoratum. Nat. Prod. Bioprospect. 2014, 4, 129–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, R.; Huang, X.; Zhang, Y.; Wu, L.; Nie, H.; Zhou, D.; Liu, B.; Deng, S.; Yang, R.; Huang, S.; et al. Structural Characterization and Assessment of the Cytotoxicity of 2,3-Dihydro-1H-indene Derivatives and Coumarin Glucosides from the Bark of Streblus indicus. J. Nat. Prod. 2016, 79, 2472–2478. [Google Scholar] [CrossRef]
- Wei, X.; Feng, C.; Li, X.H.; Mao, X.X.; Rong, L.; Yu, X.; Zhang, C.X.; Luo, H.B.; Zhang, D.M.; Ye, W.C.; et al. Enantiomeric Polyketides from the Starfish-Derived Symbiotic Fungus Penicillium sp. GGF16-1-2. Chem. Biodivers. 2019, 16, e1900052. [Google Scholar] [CrossRef]
- Ma, L.Y.; Zhang, H.B.; Kang, H.H.; Zhong, M.J.; Liu, D.S.; Liu, W.Z.; Ren, H. New Butenolides and Cyclopentenones from Saline Soil-Derived Fungus Aspergillus Sclerotiorum. Molecules 2019, 24, 2642. [Google Scholar] [CrossRef] [Green Version]
- He, J.W.; Wang, C.X.; Yang, L.; Chen, G.D.; Hu, D.; Guo, L.D.; Yao, X.S.; Gao, H. A Pair of New Polyketide Enantiomers from Three Endolichenic Fungal Strains Nigrospora sphaerica, Alternaria alternata, and Phialophora sp. Nat. Prod. Commun. 2016, 11, 829–831. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Fu, P.; Zhang, Y.; Liu, X.; Ren, F.; Che, Y. Sporulosol, a new ketal from the fungus Paraconiothyrium sporulosum. Molecules 2018, 23, 1263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, J.; Li, X.M.; Li, X.; Li, H.L.; Meng, L.H.; Wang, B.-G. New lactone and isocoumarin derivatives from the marine mangrove-derived endophytic fungus Penicillium coffeae MA-314. Phytochem. Lett. 2019, 32, 1–5. [Google Scholar] [CrossRef]
- Liu, X.; Chen, C.; Zheng, Y.; Zhang, M.; Tong, Q.; Liu, J.; Zhou, Q.; Wang, J.; Luo, Z.; Zhu, H.; et al. (±)-Peniorthoesters A and B, Two Pairs of Novel Spiro-Orthoester en-antiomers With an Unusual 1,4,6-Trioxaspi-ro[4.5]decane-7-One Unit From Penicillium minioluteum. Front. Chem. 2018, 6, 605. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.J.; Chen, S.F.; Xu, X.W.; Zhao, D.; Wang, H.F.; Bai, J.; Hua, H.M.; Chen, G.; Pei, Y.H.; Lu, X.J.; et al. One pair of new cyclopentaisochromenone enantiomer from Alternaria sp. TNXY-P-1 and their cytotoxic activity. J. Asian Nat. Prod. Res. 2018, 20, 328–336. [Google Scholar] [CrossRef]
- Tang, J.W.; Xu, H.C.; Wang, W.G.; Hu, K.; Zhou, Y.F.; Chen, R.; Li, X.N.; Du, X.; Sun, H.D.; Puno, P.-T. (+)- And (-)-Alternarilactone A: Enantiomers with a Diepoxy-Cage-like Scaffold from an Endophytic Alternaria sp. J. Nat. Prod. 2019, 82, 735–740. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, X.B.; Li, T.X.; Yang, M.H.; Kong, L.Y. Bioactive metabolites from the endophytic fungus Alternaria alternata. Fitoterapia 2014, 99, 153–158. [Google Scholar] [CrossRef]
- Intaraudom, C.; Bunbamrung, N.; Dramae, A.; Boonyuen, N.; Choowong, W.; Rachtawee, P.; Pittayakhajonwut, P. Chromone derivatives, R- and S- taeniolin, from the marine-derived fungus Taeniolella sp. BCC31839. Nat. Prod. Res. 2021, 35, 392–398. [Google Scholar] [CrossRef]
- Wu, Y.H.; Xiao, G.K.; Chen, G.D.; Wang, C.X.; Hu, D.; Lian, Y.Y.; Lin, F.; Guo, L.D.; Yao, X.S.; Gao, H. Pericocins A-D, New Bioactive Compounds from Periconia sp. Nat. Prod. Commun. 2015, 10, 2127–2130. [Google Scholar] [CrossRef] [Green Version]
- Guo, D.L.; Li, X.H.; Feng, D.; Jin, M.Y.; Cao, Y.M.; Cao, Z.X.; Deng, F.; Deng, Y.; Gu, Y.C.; Geng, Z. Novel Polyketides Produced by the Endophytic Fungus Aspergillus Fumigatus from Cordyceps Sinensis. Molecules 2018, 23, 1709. [Google Scholar] [CrossRef] [Green Version]
- Li, E.; Zhang, F.; Niu, S.; Liu, X.; Liu, G.; Che, Y. A Spiro[chroman-3,7’-isochromene]-4,6’(8’H)-dione from the Cordyceps-Colonizing Fungus Fimetariella sp. Org. Lett. 2012, 14, 3320–3323. [Google Scholar] [CrossRef]
- Li, X.H.; Han, X.H.; Qin, L.L.; He, J.L.; Cao, Z.X.; Guo, D.L.; Deng, Y.; Gu, Y.C. Isochromanes from Aspergillus fumigatus, an endophytic fungus from Cordyceps sinensis. Nat. Prod. Res. 2019, 33, 1870–1875. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Gao, W.; Zhang, M.; Li, Y.L.; Li, L.; Li, X.B.; Chang, W.Q.; Zhao, Z.T.; Lou, H.X. p-Terphenyl Derivatives from the Endolichenic Fungus Floricola striata. J. Nat. Prod. 2016, 79, 2188–2194. [Google Scholar] [CrossRef] [PubMed]
- Arunrattiyakorn, P.; Kuno, M.; Aree, T.; Laphookhieo, S.; Sriyatep, T.; Kanzaki, H.; Garcia Chavez, M.A.; Wang, Y.A.; Andersen, R.J. Biotransformation of β-Mangostin by an Endophytic Fungus of Garcinia mangostana to Furnish Xanthenes with an Unprecedented Heterocyclic Skeleton. J. Nat. Prod. 2018, 81, 2244–2250. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Chen, S.; Qiu, P.; Tan, C.; Long, Y.; Lu, Y.; She, Z. (+)- and (-)-Ascomlactone A: A pair of novel dimeric polyketides from a mangrove endophytic fungus Ascomycota sp. SK2YWS-L. Org. Biomol. Chem. 2017, 15, 10276–10280. [Google Scholar] [CrossRef]
- Liu, Z.; Qiu, P.; Li, J.; Chen, G.; Chen, Y.; Liu, H.; She, Z. Anti-inflammatory polyketides from the mangrove-derived fungus Ascomycota sp. SK2YWS-L. Tetrahedron 2018, 74, 746–751. [Google Scholar] [CrossRef]
- Zhang, X.; Wu, Z.; Lai, Y.; Li, D.; Wang, J.; Luo, Z.; Xue, Y.; Zhu, H.; Chen, C.; Zhang, Y. (±)-Terreinlactone A, a pair of 3-substituted δ-lactone enantiomers derived from terrein from the fungus Aspergillus terreus. Chem. Pharm. Bull. 2018, 66, 764–767. [Google Scholar] [CrossRef] [Green Version]
- Hu, C.H.; Zhou, Y.H.; Xie, F.; Li, Y.L.; Zhao, Z.T.; Lou, H.X. Two new α-pyrone derivatives from an endolichenic fungus Tolypocladium sp. J. Asian. Nat. Prod. Res. 2017, 19, 786–792. [Google Scholar] [CrossRef]
- Sang, X.N.; Chen, S.F.; Chen, G.; An, X.; Li, S.G.; Lu, X.J.; Zhao, D.; Bai, J.; Wang, H.F.; Pei, Y.H. Two pairs of enantiomeric α-pyrone dimers from the endophytic fungus Phoma sp. YN02-P-3. RSC Adv. 2017, 7, 1943–1946. [Google Scholar] [CrossRef] [Green Version]
- Miyano, R.; Matsuo, H.; Nonaka, K.; Mokudai, T.; Niwano, Y.; Shiomi, K.; Takahashi, Y.; Omura, S.; Nakashima, T. Pochoniolides A and B, new antioxidants from the fungal strain Pochonia chlamydosporia var. spinulospora FKI-7537. J. Biosci. Bioeng. 2018, 126, 661–666. [Google Scholar]
- Qi, B.; Liu, X.; Mo, T.; Li, S.S.; Wang, J.; Shi, X.P.; Wang, X.H.; Zhu, Z.X.; Zhao, Y.F.; Jin, H.W.; et al. Nitric oxide inhibitory polyketides from Penicillium chrysogenum MT-12, an endophytic fungus isolated from Huperzia serrata. Fitoterapia 2017, 123, 35–43. [Google Scholar] [CrossRef]
- He, F.; Li, X.; Yu, J.H.; Zhang, X.; Nong, X.; Chen, G.; Zhu, K.; Wang, Y.Y.; Bao, J.; Zhang, H. Secondary metabolites from the mangrove sediment-derived fungus Penicillium pinophilum SCAU037. Fitoterapia 2019, 136, 104177. [Google Scholar] [CrossRef] [PubMed]
- Zang, Y.; Gong, Y.H.; Li, X.W.; Li, X.N.; Liu, J.J.; Chen, C.M.; Zhou, Y.; Gu, L.H.; Luo, Z.W.; Wang, J.P.; et al. Canescones A-E: Aromatic polyketide dimers with PTP1B inhibitory activity from Penicillium canescens. Org. Chem. Front. 2019, 6, 3274–3281. [Google Scholar] [CrossRef]
- Xu, L.L.; Chen, H.L.; Hai, P.; Gao, Y.; Xie, C.D.; Yang, X.L.; Abe, I. (+)- And (−)-Preuisolactone A: A Pair of Caged Norsesquiterpenoidal Enantiomers with a Tricyclo[4.4.01,6.02,8]decane Carbon Skeleton from the Endophytic Fungus Preussia isomera. Org. Lett. 2019, 21, 1078–1081. [Google Scholar] [CrossRef] [PubMed]
- Song, R.Y.; Liu, Y.; Liu, R.H.; Wang, X.B.; Li, T.X.; Kong, L.Y.; Yang, M.H. Benzophenone derivatives from the plant endophytic fungus, Pestalotiopsis sp. Phytochem. Lett. 2017, 22, 189–193. [Google Scholar] [CrossRef]
- Meng, L.H.; Li, X.M.; Liu, Y.; Wang, B.G.; Meng, L.H.; Liu, Y.; Mandi, A.; Kurtan, T. Isolation, Stereochemical Study, and Antioxidant Activity of Benzofuranone Derivatives from a Mangrove-derived Fungus Eurotium rubrum MA-150. Chirality 2016, 28, 581–584. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.Y.; Kuang, R.Q.; Chen, G.D.; Qin, S.Y.; Wang, C.X.; Hu, D.; Wu, B.; Liu, X.Z.; Yao, X.S.; Gao, H. Three pairs of new isopentenyl dibenzo[b,e]oxepinone enantiomers from Talaromyces flavus, a wetland soil-derived fungus. Molecules 2016, 21, 1184. [Google Scholar] [CrossRef] [Green Version]
- Hammerschmidt, L.; Ola, A.; Mueller, W.E.G.; Lin, W.; Mandi, A.; Kurtan, T.; Proksch, P.; Aly, A.H. Two new metabolites from the endophytic fungus Xylaria sp. isolated from the medicinal plant Curcuma xanthorrhiza. Tetrahedron Lett. 2015, 56, 1193–1197. [Google Scholar] [CrossRef]
- Liu, H.; Tan, H.; Wang, W.; Zhang, W.; Chen, Y.; Li, S.; Liu, Z.; Li, H.; Zhang, W. Cytorhizophins A and B, benzophenone-hemiterpene adducts from the endophytic fungus Cytospora rhizophorae. Org. Chem. Front. 2019, 6, 591–596. [Google Scholar] [CrossRef]
- Wu, J.C.; Hou, Y.; Xu, Q.; Fang, J.; Wu, Q.X.; Jin, X.J.; Chen, Y.; Hu, B. (±)-Alternamgin, a Pair of Enantiomeric Polyketides, from the Endophytic Fungi Alternaria sp. MG1. Org. Lett. 2019, 21, 1551–1554. [Google Scholar] [CrossRef]
- Zhong, W.M.; Wang, J.F.; Wei, X.Y.; Zeng, Q.; Chen, X.Y.; Xiang, Y.; Tian, X.P.; Zhang, S.; Long, L.J.; Wang, F.-Z. (+)- And (−)-Eurotone A: A pair of enantiomeric polyketide dimers from a marine-derived fungus Eurotium sp. SCSIO F452. Tetrahedron Lett. 2019, 60, 1600–1603. [Google Scholar] [CrossRef]
- Shaker, S.; Fan, R.Z.; Lan, W.J.; Li, H.J. A pair of novel bisindole alkaloid enantiomers from marine fungus Fusarium sp. XBB-9. Nat. Prod. Res. 2021, 35, 1497–1503. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Wang, L.; Wang, B.; Xu, Y.; Zhu, G.; Lan, M.; Zhu, W.; Sun, K. Diketopiperazine and diphenylether derivatives from marine algae-derived Aspergillus versicolor OUCMDZ-2738 by epigenetic activation. Mar. Drugs 2019, 17, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, J.; Li, X.M.; Meng, L.H.; Konuklugil, B.; Li, X.; Li, H.L.; Wang, B.G. Isolation and characterization of three pairs of indolediketopiperazine enantiomers containing infrequent N-methoxy substitution from the marine algal-derived endophytic fungus Acrostalagmus luteoalbus TK-43. Bioorg. Chem. 2019, 90, 103030. [Google Scholar] [CrossRef] [PubMed]
- Zhong, W.; Wang, J.; Wei, X.; Fu, T.; Chen, Y.; Zeng, Q.; Huang, Z.; Huang, X.; Zhang, W.; Zhang, S.; et al. Three pairs of new spirocyclic alkaloid enantiomers from the marine-derived fungus Eurotium sp. SCSIO F452. Front. Chem. 2019, 7, 350. [Google Scholar] [CrossRef]
- Gao, H.; Liu, W.; Zhu, T.; Mo, X.; Mandi, A.; Kurtan, T.; Li, J.; Ai, J.; Gu, Q.; Li, D. Diketopiperazine alkaloids from a mangrove rhizosphere soil derived fungus Aspergillus effuses H1-1. Org. Biomol. Chem. 2012, 10, 9501–9506. [Google Scholar] [CrossRef] [Green Version]
- Cai, R.; Jiang, H.; Xiao, Z.; Liu, Z.; Lin, S.e.; She, Z.; Cao, W.; Yan, T.; Long, Y.; She, Z. (−)- and (+)-Asperginulin A, a Pair of Indole Diketopiperazine Alkaloid Dimers with a 6/5/4/5/6 Pentacyclic Skeleton from the Mangrove Endophytic Fungus Aspergillus sp. SK-28. Org. Lett. 2019, 21, 9633–9636. [Google Scholar] [CrossRef]
- Chen, G.D.; Bao, Y.R.; Huang, Y.F.; Hu, D.; Li, X.X.; Guo, L.D.; Li, J.; Yao, X.S.; Gao, H. Three pairs of variecolortide enantiomers from Eurotium sp. with caspase-3 inhibitory activity. Fitoterapia 2014, 92, 252–259. [Google Scholar] [CrossRef]
- Zhong, W.; Wang, J.; Wei, X.; Chen, Y.; Fu, T.; Xiang, Y.; Huang, X.; Tian, X.; Xiao, Z.; Zhang, W.; et al. Variecolortins A-C, Three Pairs of Spirocyclic Diketopiperazine Enantiomers from the Marine-Derived Fungus Eurotium sp. SCSIO F452. Org. Lett. 2018, 20, 4593–4596. [Google Scholar] [CrossRef]
- Han, J.; Liu, C.; Li, L.; Zhou, H.; Liu, L.; Bao, L.; Chen, Q.; Song, F.; Zhang, L.; Li, E.; et al. Decalin-Containing Tetramic Acids and 4-Hydroxy-2-pyridones with Antimicrobial and Cytotoxic Activity from the Fungus Coniochaeta cephalothecoides Collected in Tibetan Plateau (Medog). J. Org. Chem. 2017, 82, 11474–11486. [Google Scholar] [CrossRef]
- Li, J.; Wang, W.X.; Chen, H.P.; Li, Z.H.; He, J.; Zheng, Y.S.; Sun, H.; Huang, R.; Yuan, Q.X.; Feng, T.; et al. (±)-Xylaridines A and B, Highly Conjugated Alkaloids from the Fungus Xylaria longipes. Org. Lett. 2019, 21, 1511–1514. [Google Scholar] [CrossRef]
- Li, J.; Wang, W.X.; Li, Z.H.; He, J.; Huang, R.; Zheng, Y.S.; Li, L.Q.; Wang, X.; Feng, T.; Liu, J.K. Xylaridines C and D, unusual thiopyranodipyridine alkaloids from the fungus Xylaria longipes. Org. Lett. 2019, 21, 6145–6148. [Google Scholar] [CrossRef]
- Chen, S.; Jiang, M.; Chen, B.; Niaz, S.I.; He, J.; Liu, L.; Chen, S.; Liu, L.; Salaenoi, J.; Niaz, S.I. Penicamide A, A Unique N,N’-Ketal Quinazolinone Alkaloid from Ascidian-Derived Fungus Penicillium sp. 4829. Mar. Drugs 2019, 17, 522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, H.; Li, F.; Ji, N. Alkaloids from an algicolous strain of Talaromyces sp. Chin. J. Oceanol. Limnol. 2016, 34, 367–371. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.K.; Gao, T.; Yang, M.Y.; Zhao, G.Z.; Zhu, H.J.; Cao, F.; Zhang, B.; Liu, L. A pair of enantiomeric 5-oxabicyclic[4.3.0]lactam derivatives and one new polyketide from the marine-derived fungus Penicillium griseofulvum. Nat. Prod. Res. 2018, 32, 2366–2369. [Google Scholar] [CrossRef] [PubMed]
- Almeida, C.; Hemberger, Y.; Schmitt, S.M.; Bouhired, S.; Natesan, L.; Kehraus, S.; Dimas, K.; Gutschow, M.; Bringmann, G.; Konig, G.M. Marilines A-C: Novel phthalimidines from the sponge-derived fungus Stachylidium sp. Chemistry 2012, 18, 8827–8834. [Google Scholar] [CrossRef]
- Kong, Z.; Jing, R.; Geng, Y.; Ji, J.; Wu, Y.; Guo, Y.; Qin, L.; Zheng, C. Trichodermadiones A and B from the solid culture of Trichoderma atroviride S361, an endophytic fungus in Cephalotaxus fortunei. Fitoterapia 2018, 127, 362–366. [Google Scholar] [CrossRef]
- Wen, H.; Li, Y.; Liu, X.; Ye, W.; Yao, X.; Che, Y. Fusagerins A-F, New Alkaloids from the Fungus Fusarium sp. Nat. Prod. Bioprospect. 2015, 5, 195–203. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Chen, X.; Li, D.; Zhang, Y.; Li, L.; Guo, L.; Cao, Y.; Che, Y. Bisabolane Sesquiterpenoids from the Plant Endophytic Fungus Paraconiothyrium brasiliense. J. Nat. Prod. 2015, 78, 746–753. [Google Scholar] [CrossRef]
- Jia, Y.L.; Wei, M.Y.; Guan, F.F.; Wang, C.Y.; Shao, C.L.; Jia, Y.L.; Wei, M.Y.; Chen, H.Y. (+)- and (−)-Pestaloxazine A, a Pair of Antiviral Enantiomeric Alkaloid Dimers with a Symmetric Spiro[oxazinane-piperazinedione] Skeleton from Pestalotiopsis sp. Org. Lett. 2015, 17, 4216–4219. [Google Scholar] [CrossRef]
- Yan, Y.M.; Zhang, H.X.; Liu, H.; Wu, J.B.; Li, Y.P.; Cheng, Y.X.; Wang, Y. (+/−)-Lucidumone, a COX-2 Inhibitory Caged Fungal Meroterpenoid from Ganoderma lucidum. Org. Lett. 2019, 21, 8523–8527. [Google Scholar] [CrossRef]
- Peng, X.R.; Liu, J.Q.; Wan, L.S.; Li, X.N.; Yan, Y.X.; Qiu, M.H. Four New Polycyclic Meroterpenoids from Ganoderma cochlear. Org. Lett. 2014, 16, 4838–4841. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.M.; Ai, J.; Zhou, L.L.; Chung, A.C.K.; Li, R.; Nie, J.; Fang, P.; Wang, X.L.; Luo, J.; Hu, Q.; et al. Lingzhiols, Unprecedented Rotary Door-Shaped Meroterpenoids as Potent and Selective Inhibitors of p-Smad3 from Ganoderma lucidum. Org. Lett. 2013, 15, 5488–5491. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Chen, L.; Li, S.; Zhao, J. Meroterpenoids from the fruiting bodies of higher fungus Ganoderma resinaceum. Phytochem. Lett. 2017, 22, 214–218. [Google Scholar] [CrossRef]
- Luo, Q.; Di, L.; Yang, X.H.; Cheng, Y.X. Applanatumols A and B, meroterpenoids with unprecedented skeletons from Ganoderma applanatum. RSC Adv. 2016, 6, 45963–45967. [Google Scholar] [CrossRef]
- Luo, Q.; Wang, X.L.; Di, L.; Yan, Y.M.; Lu, Q.; Yang, X.H.; Hu, D.B.; Cheng, Y.X. Isolation and identification of renoprotective substances from the mushroom Ganoderma lucidum. Tetrahedron 2015, 71, 840–845. [Google Scholar] [CrossRef]
- Wang, X.F.; Yan, Y.M.; Wang, X.L.; Ma, X.J.; Fu, X.Y.; Cheng, Y.X. Two new compounds from Ganoderma lucidum. J. Asian Nat. Prod. Res. 2015, 17, 329–332. [Google Scholar] [CrossRef]
- Luo, Q.; Tu, Z.C.; Cheng, Y.X. Two rare meroterpenoidal rotamers from Ganoderma applanatum. RSC Adv. 2017, 7, 3413–3418. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Wang, F.; Xu, F.; Ding, L.Q.; Zhang, Q.; Li, H.X.; Zhao, F.; Wang, L.Q.; Zhu, L.H.; Chen, L.X.; et al. Two pairs of farnesyl phenolic enantiomers as natural nitric oxide inhibitors from Ganoderma sinense. Bioorg. Med. Chem. Lett. 2016, 26, 3342–3345. [Google Scholar] [CrossRef]
- Li, L.; Li, H.; Peng, X.R.; Hou, B.; Yu, M.Y.; Dong, J.R.; Li, X.N.; Zhou, L.; Yang, J.; Qiu, M.H. (±)-Ganoapplanin, a Pair of Polycyclic Meroterpenoid Enantiomers from Ganoderma applanatum. Org. Lett. 2016, 18, 6078–6081. [Google Scholar] [CrossRef]
- Luo, Q.; Wang, Z.; Luo, J.-F.; Tu, Z.C.; Cheng, Y.X. (±)-Applanatumines B-D: Novel dimeric meroterpenoids from Ganoderma applanatum as inhibitors of JAK3. RSC Adv. 2017, 7, 38037–38043. [Google Scholar] [CrossRef] [Green Version]
- Qin, F.Y.; Cheng, Y.X.; Qin, F.Y.; Cheng, Y.X.; Yan, Y.M.; Cheng, Y.X.; Tu, Z.C. (±) Gancochlearols A and B: Cytotoxic and COX-2 inhibitory meroterpenoids from Ganoderma cochlear. Nat. Prod. Res. 2020, 34, 2269–2275. [Google Scholar] [CrossRef] [PubMed]
- Qin, F.Y.; Cheng, Y.X.; Qin, F.Y.; Cheng, Y.X.; Yan, Y.M.; Cheng, Y.X.; Tu, Z.C. (±) Cochlearoids N-P: Three pairs of phenolic meroterpenoids from the fungus Ganoderma cochlear and their bioactivities. J. Asian Nat. Prod. Res. 2019, 21, 542–550. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.J.; Nian, Y.; Yan, Y.; Gong, Y.; Luo, Q.; Zhang, Y.; Hou, B.; Zuo, Z.L.; Wang, S.M.; Jiang, H.H.; et al. Two new classes of T-type calcium channel inhibitors with new chemical scaffolds from Ganoderma cochlear. Org. Lett. 2015, 17, 3082–3085. [Google Scholar] [CrossRef] [PubMed]
- Li, X.C.; Liu, F.; Su, H.G.; Guo, L.; Zhou, Q.M.; Huang, Y.J.; Peng, C.; Xiong, L. Two pairs of alkaloid enantiomers from Ganoderma luteomarginatum. Biochem. Syst. Ecol. 2019, 86, 103930. [Google Scholar] [CrossRef]
- Nord, C.; Menkis, A.; Broberg, A. Cytotoxic illudane sesquiterpenes from the fungus Granulobasidium vellereum (Ellis and Cragin) Jülich. J. Nat. Prod. 2015, 78, 2559–2564. [Google Scholar] [CrossRef]
- McMorris, T.C.; Kelner, M.J.; Chadha, R.K.; Siegel, J.S.; Moon, S.S.; Moya, M.M. Structure and reactivity of illudins. Tetrahedron 1989, 45, 5433–5440. [Google Scholar] [CrossRef]
- Kinder, F.R., Jr.; Bair, K.W. Total Synthesis of (±)-Illudin M. J. Org. Chem. 1994, 59, 6965–6967. [Google Scholar] [CrossRef]
- Zhang, S.; Zhang, L.; Fu, X.; Li, Z.; Guo, L.; Kou, L.; Liu, M.; Xie, Z. (+)- and (−)-actinoxocine, and actinaphthorans A-B, C-ring expansion and cleavage angucyclinones from a marine-derived Streptomyces sp. Org. Chem. Front. 2019, 6, 3925–3928. [Google Scholar] [CrossRef]
- Yi, W.; Li, Q.; Song, T.; Chen, L.; Li, X.C.; Zhang, Z.; Lian, X.Y. Isolation, structure elucidation, and antibacterial evaluation of the metabolites produced by the marine-sourced Streptomyces sp. ZZ820. Tetrahedron 2019, 75, 1186–1193. [Google Scholar] [CrossRef]
- Zhang, S.; Yang, Q.; Guo, L.; Zhang, Y.; Feng, L.; Zhou, L.; Yang, S.; Yao, Q.; Pescitelli, G.; Xie, Z. Isolation, structure elucidation and racemization of (+)- and (−)-pratensilins A-C: Unprecedented spiro indolinone-naphthofuran alkaloids from a marine Streptomyces sp. Chem. Commun. 2017, 53, 10066–10069. [Google Scholar] [CrossRef]
- Liang, Y.Q.; Liao, X.J.; Zhao, B.X.; Xu, S.H.; Liang, Y.Q.; Xu, S.H. (+)- and (−)-Spongiterpene, a pair of new valerenane sesquiterpene enantiomers from the marine sponge Spongia sp. Nat. Prod. Res. 2021, 35, 2178–2183. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.Y.; Han, G.Y.; Yang, N.N.; Lan, L.F.; Li, X.W.; Guo, Y.W. Racemic trinorsesquiterpenoids from the Beihai sponge Spongia officinalis: Structure and biomimetic total synthesis. Org. Chem. Front. 2018, 5, 1022–1027. [Google Scholar] [CrossRef]
- Jiao, W.H.; Hong, L.L.; Sun, J.B.; Piao, S.J.; Chen, G.D.; Deng, H.; Wang, S.P.; Yang, F.; Lin, H.-W. (±)-Hippolide J, A Pair of Unusual Antifungal Enantiomeric Sesterterpenoids from the Marine Sponge Hippospongia lachne. Eur. J. Org. Chem. 2017, 2017, 3421–3426. [Google Scholar] [CrossRef]
- Afifi, A.H.; Kagiyama, I.; El-Desoky, A.H.; Kato, H.; Mangindaan, R.E.P.; de Voogd, N.J.; Ammar, N.M.; Hifnawy, M.S.; Tsukamoto, S. Sulawesins A-C, Furanosesterterpene Tetronic Acids That Inhibit USP7, from a Psammocinia sp. Marine Sponge. J. Nat. Prod. 2017, 80, 2045–2050. [Google Scholar] [CrossRef]
- Zhu, Y.; Wang, Y.; Gu, B.B.; Yang, F.; Jiao, W.H.; Hu, G.H.; Yu, H.B.; Han, B.N.; Zhang, W.; Shen, Y.; et al. Antifungal bromopyrrole alkaloids from the South China Sea sponge Agelas sp. Tetrahedron 2016, 72, 2964–2971. [Google Scholar] [CrossRef]
- Allen, E.E.; Zhu, C.; Panek, J.S.; Schaus, S.E. Multicomponent Condensation Reactions via ortho-Quinone Methides. Org. Lett. 2017, 19, 1878–1881. [Google Scholar] [CrossRef] [Green Version]
- Chu, M.J.; Tang, X.L.; Qin, G.F.; Sun, Y.T.; Li, L.; de Voogd, N.J.; Li, P.L.; Li, G.Q. Pyrrole Derivatives and Diterpene Alkaloids from the South China Sea Sponge Agelas nakamurai. Chem. Biodivers. 2017, 14. [Google Scholar] [CrossRef]
- Wang, Q.; Tang, X.; Luo, X.; de Voogd, N.J.; Li, P.; Li, G. (+)- and (−)-Spiroreticulatine, A Pair of Unusual Spiro Bisheterocyclic Quinoline-imidazole Alkaloids from the South China Sea Sponge Fascaplysinopsis reticulata. Org. Lett. 2015, 17, 3458–3461. [Google Scholar] [CrossRef]
- Shirouzu, T.; Watari, K.; Ono, M.; Koizumi, K.; Saiki, I.; Tanaka, C.; van Soest, R.W.M.; Miyamoto, T. Structure, synthesis, and biological activity of a C-20 bisacetylenic alcohol from a marine sponge Callyspongia sp. J. Nat. Prod. 2013, 76, 1337–1342. [Google Scholar] [CrossRef]
- Jang, K.H.; Lee, Y.; Sim, C.J.; Oh, K.B.; Shin, J. Bioactive lipids from the sponge Spirastrella abata. Bioorg. Med. Chem. Lett. 2012, 22, 1078–1081. [Google Scholar] [CrossRef]
- Alam, N.; Wang, W.; Hong, J.; Lee, C.-O.; Im, K.S.; Jung, J.H. Cytotoxic sphingosine 4-sulfates from the sponge Spirastrella abata. J. Nat. Prod. 2002, 65, 944–945. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.M.; Li, L.J.; Qin, X.C.; Lu, Q.; Tu, Z.C.; Cheng, Y.X. Compounds from the insect Blaps japanensis with COX-1 and COX-2 inhibitory activities. Bioorg. Med. Chem. Lett. 2015, 25, 2469–2472. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.M.; Ai, J.; Shi, Y.N.; Zuo, Z.L.; Hou, B.; Luo, J.; Cheng, Y.X. (±)-Aspongamide A, an N-Acetyldopamine Trimer Isolated from the Insect Aspongopus chinensis, Is an Inhibitor of p-Smad3. Org. Lett. 2014, 16, 532–535. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.J.; Yan, Y.M.; Tu, Z.C.; Luo, J.F.; Liang, R.; Yang, T.H.; Cheng, Y.X.; Wang, S.M. Compounds from Polyphaga plancyi and their inhibitory activities against JAK3 and DDR1 kinases. Fitoterapia 2016, 114, 163–167. [Google Scholar] [CrossRef]
- Chan, S.T.S.; Nani, R.R.; Schauer, E.A.; Martin, G.E.; Williamson, R.T.; Sauri, J.; Buevich, A.V.; Schafer, W.A.; Joyce, L.A.; Goey, A.K.L.; et al. Characterization and Synthesis of Eudistidine C, a Bioactive Marine Alkaloid with an Intriguing Molecular Scaffold. J. Org. Chem. 2016, 81, 10631–10640. [Google Scholar] [CrossRef]
- Wang, L.; Ma, Y.T.; Sun, Q.Y.; Li, X.N.; Yan, Y.; Yang, J.; Yang, F.M.; Liu, F.Y.; Zang, Z.; Wu, X.H.; et al. Structurally diversified diterpenoids from Euphorbia dracunculoides. Tetrahedron 2015, 71, 5484–5493. [Google Scholar] [CrossRef]
- Marin-Saez, J.; Romero-Gonzalez, R.; Garrido, F.A. Enantiomeric determination and evaluation of the racemization process of atropine in Solanaceae seeds and contaminated samples by high performance liquid chromatography-tandem mass spectrometry. J. Chromatogr. A 2016, 1474, 79–84. [Google Scholar] [CrossRef] [Green Version]
- Capon, R.J. Extracting value: Mechanistic insights into the formation of natural product artifacts-case studies in marine natural products. Nat. Prod. Rep. 2020, 37, 55–79. [Google Scholar] [CrossRef]
- Li, L.-Z.; Sun, X.; Qi, X.-L.; Song, S.-J.; Liang, X.; Wang, J.; Zhao, Q.C.; Song, J. Bioactive norditerpenoids and neolignans from the roots of salvia miltiorrhiza. Org. Biomo.l Chem. 2016, 14, 10050–10057. [Google Scholar] [CrossRef]
- Song, X.-Q.; Zhu, K.; Yu, J.-H.; Zhang, Q.; Zhang, Y.; He, F.; Cheng, Z.-Q.; Jiang, C.-S.; Bao, J.; Zhang, H. New Octadecanoid Enantiomers from the Whole Plants of Plantago depressa. Molecules 2018, 23, 1723. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Jiang, J.; Liu, Z.; Lin, S.; Xia, G.; Xia, X.; Ding, B.; He, L.; Lu, Y.; She, Z. Peniphenones A-D from the mangrove fungus Penicillium dipodomyicola HN4-3A as inhibitors of Mycobacterium tuberculosis phosphatase MptpB. J. Nat. Prod. 2014, 77, 800–806. [Google Scholar] [CrossRef]
- Cao, W.-W.; Luo, Q.; Cheng, Y.-X.; Wang, S.-M. Meroterpenoid enantiomers from Ganoderma sinensis. Fitoterapia 2016, 110, 110–115. [Google Scholar] [CrossRef]
- Zhang, X.; Li, P.-L.; Qin, G.-F.; Li, S.; de Voogd, N.J.; Tang, X.-L.; Li, G.Q. Isolation and absolute configurations of diversiform C17, C21 and C25 terpenoids from the marine sponge Cacospongia sp. Mar. Drugs 2019, 17, 14. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Structures of 8,4′-oxyneolignans.

Figure 2.
Structures of other acyclic lignans.

Scheme 1.
Plausible biosynthetic pathway for 39.

Figure 3.
Structures of Furan-incorporating lignans.

Figure 4.
Structures of other cyclic lignans.

Figure 5.
Structures of sesquineolignans.

Scheme 2.
Plausible biosynthetic pathways for 81a/81b.

Figure 6.
Structures of coumarins.

Figure 7.
Structures of simple phenylpropanoids.

Figure 8.
Structures of indole alkaloids.

Figure 9.
Structures of quinoline and isoquinoline alkaloids.

Scheme 3.
Plausible biosynthetic pathways for 171 and 172.

Figure 10.
Structures of β-carboline and carbazole alkaloids.

Figure 11.
Structures of piperidine alkaloids.

Figure 12.
Structures of thiohydantoin alkaloids.

Figure 13.
Structures of indolizidine and quinolizidine alkaloids.

Figure 14.
Structures of other alkaloids.

Figure 15.
Structures of flavones and isoflavones.

Figure 16.
Structures of chalcones.

Figure 17.
Structures of xanthones.

Scheme 4.
Plausible biosynthetic pathways for 300a/300b−302a/302b.

Figure 18.
Structures of sesquiterpenoids.

Figure 19.
Structures of diterpenoids.

Figure 20.
Structures of meroterpenoids.

Figure 21.
Structures of phloroglucinols.

Scheme 5.
Plausible biosynthetic pathways for 378a/378b and 379a/379b.

Scheme 6.
Plausible biosynthetic pathways for 381a/381b.

Figure 22.
Structures of naphthalene and phenanthrenes.

Figure 23.
Structures of chromanes (* abs. configs. undetermined).

Figure 24.
Structures of acetophenones.

Figure 25.
Structures of diarylheptanoids.

Figure 26.
Structures of triarylmethanes.

Figure 27.
Structures of fatty acids.

Figure 28.
Structures of miscellaneous classes.

Scheme 7.
Plausible biosynthetic pathways for 457a/457b.

Figure 29.
(a) Structures of nonalkaloids from phylum Ascomycota (Part 1). (b) Structures of nonalkaloids from phylum Ascomycota (Part 2).
Figure 29.
(a) Structures of nonalkaloids from phylum Ascomycota (Part 1). (b) Structures of nonalkaloids from phylum Ascomycota (Part 2).


Scheme 8.
Plausible biosynthetic pathways for 499a/499b and 500a/500b.

Figure 30.
Structures of alkaloids from phylum Ascomycota.

Scheme 9.
Plausible biosynthetic pathways for 526/526b−529a/529b.

Scheme 10.
Plausible biosynthetic pathway for 530a/530b.

Scheme 11.
Plausible biosynthetic pathways for 535a/535b−537a/537b.

Figure 31.
(a) Structures of metabolites from phylum Basidiomycota (Part 1); (b) Structures of metabolites from phylum Basidiomycota (Part 2).
Figure 31.
(a) Structures of metabolites from phylum Basidiomycota (Part 1); (b) Structures of metabolites from phylum Basidiomycota (Part 2).


Figure 32.
Structures of metabolites from actinomycetes.

Figure 33.
Structures of metabolites from phylum Porifera.

Figure 34.
Structures of metabolites from phyla Arthropoda and Chordata.

Figure 35.
A comparison of enantiomeric pairs from plants and other sources.

Figure 36.
Distributions of enantiomers in kingdoms Plantae, Fungi, Animalia and Prokaryota.

Figure 37.
Statistics of different types of enantiomeric pairs from plants.


{kind=link}
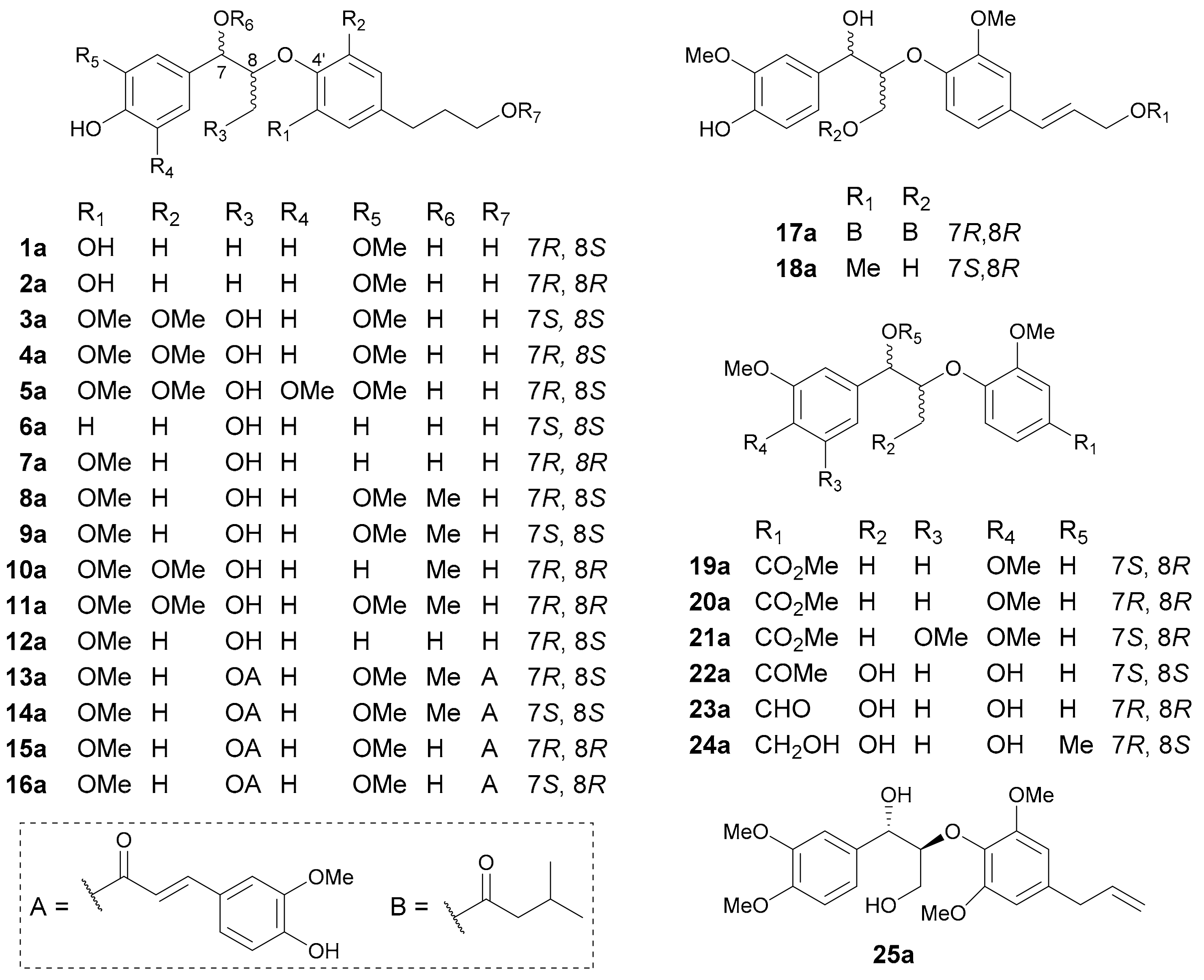{kind=link}
{kind=link}
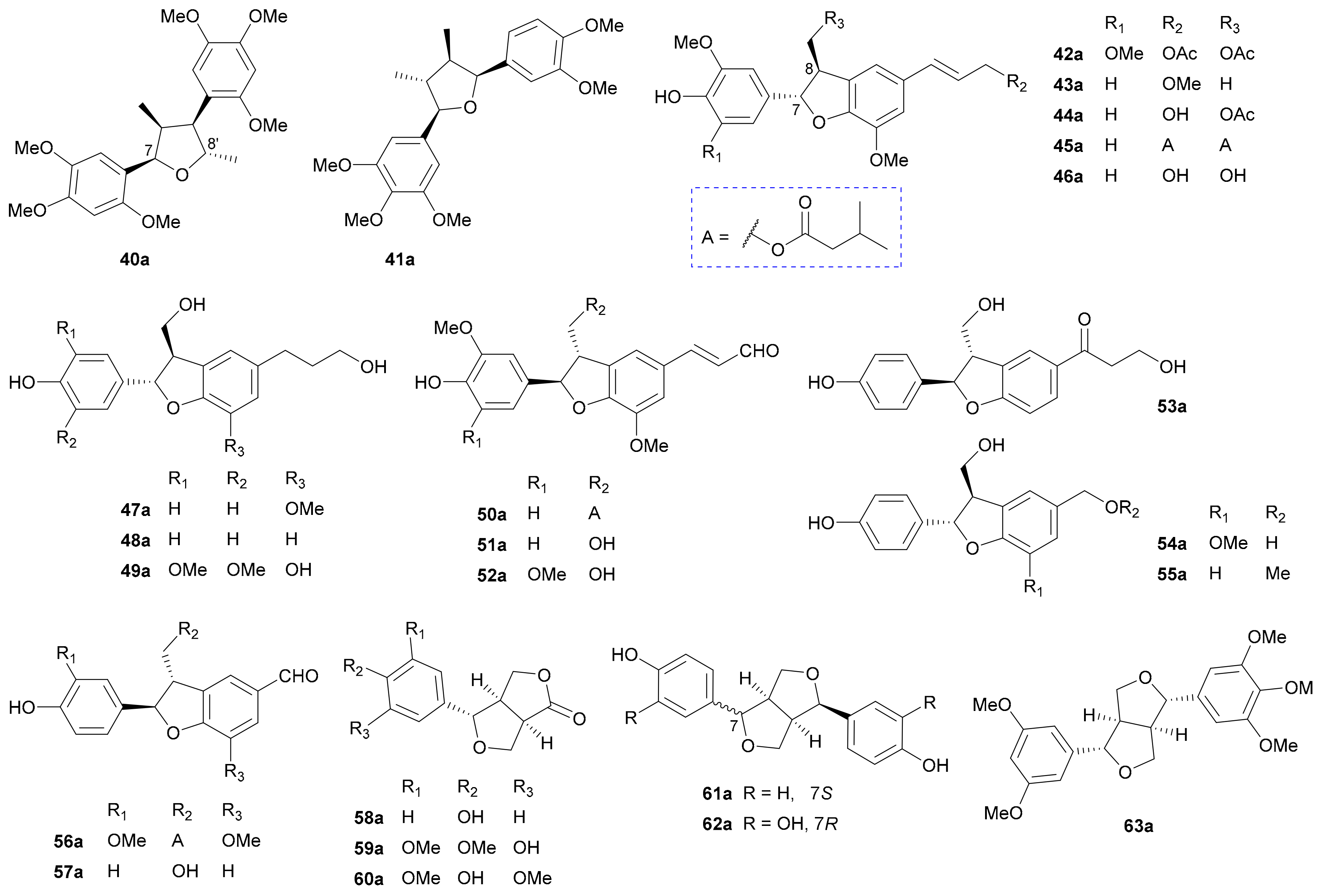{kind=link}
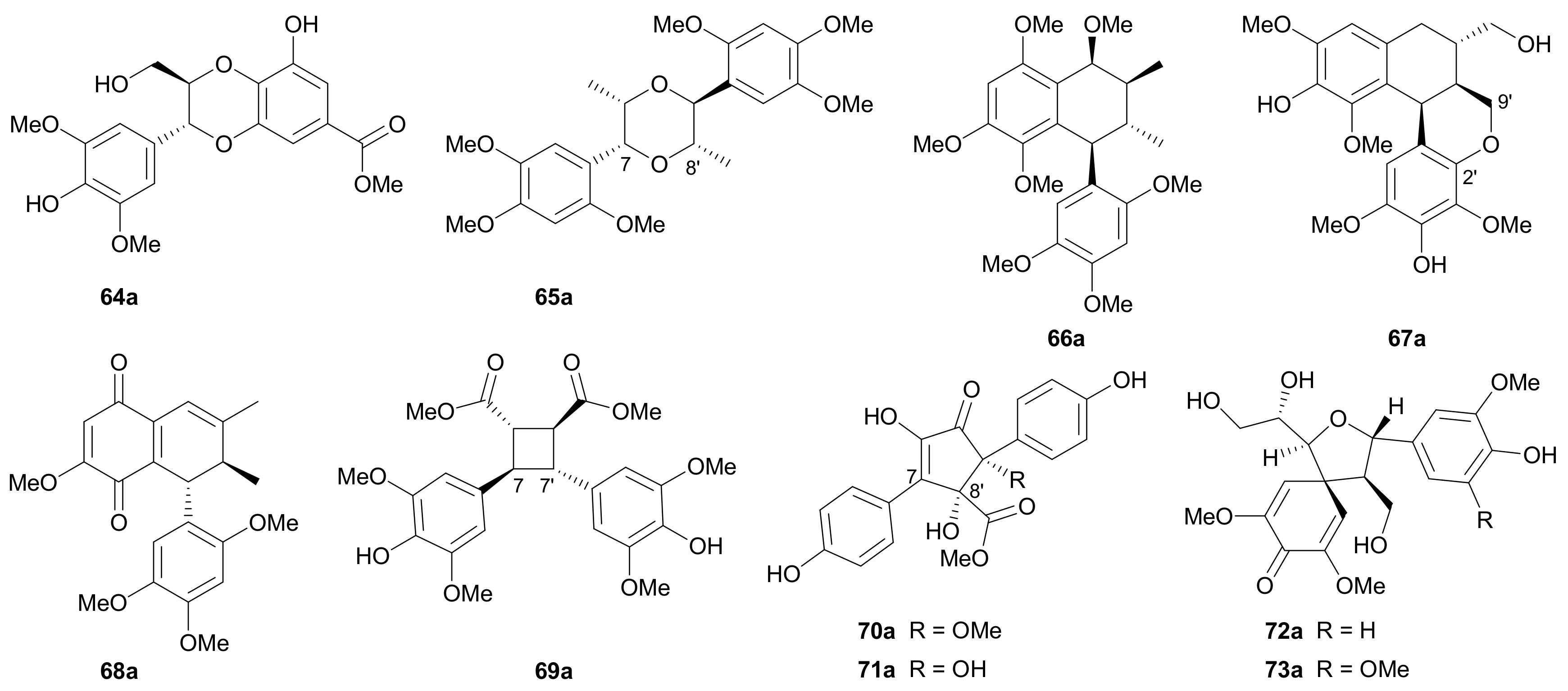{kind=link}
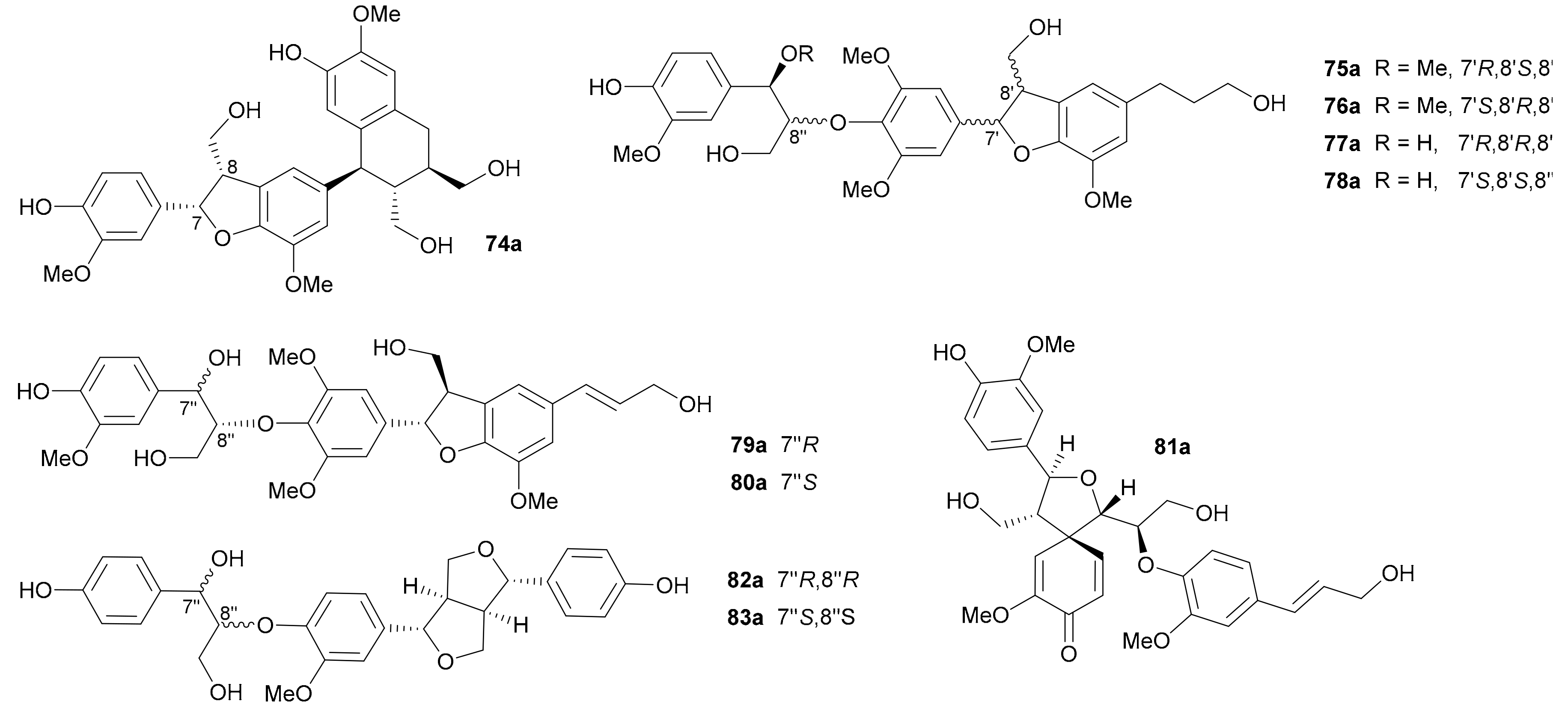{kind=link}
{kind=link}
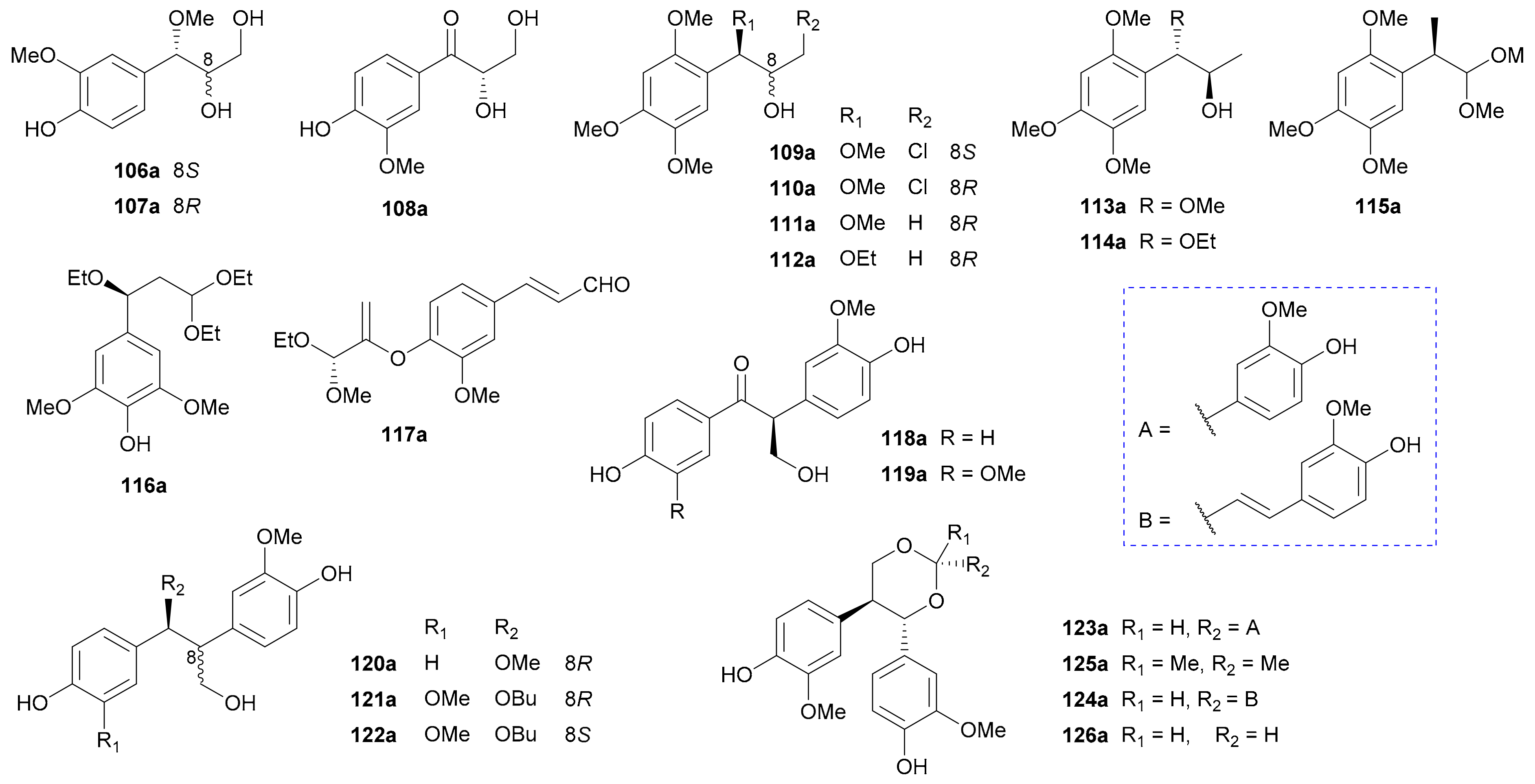{kind=link}
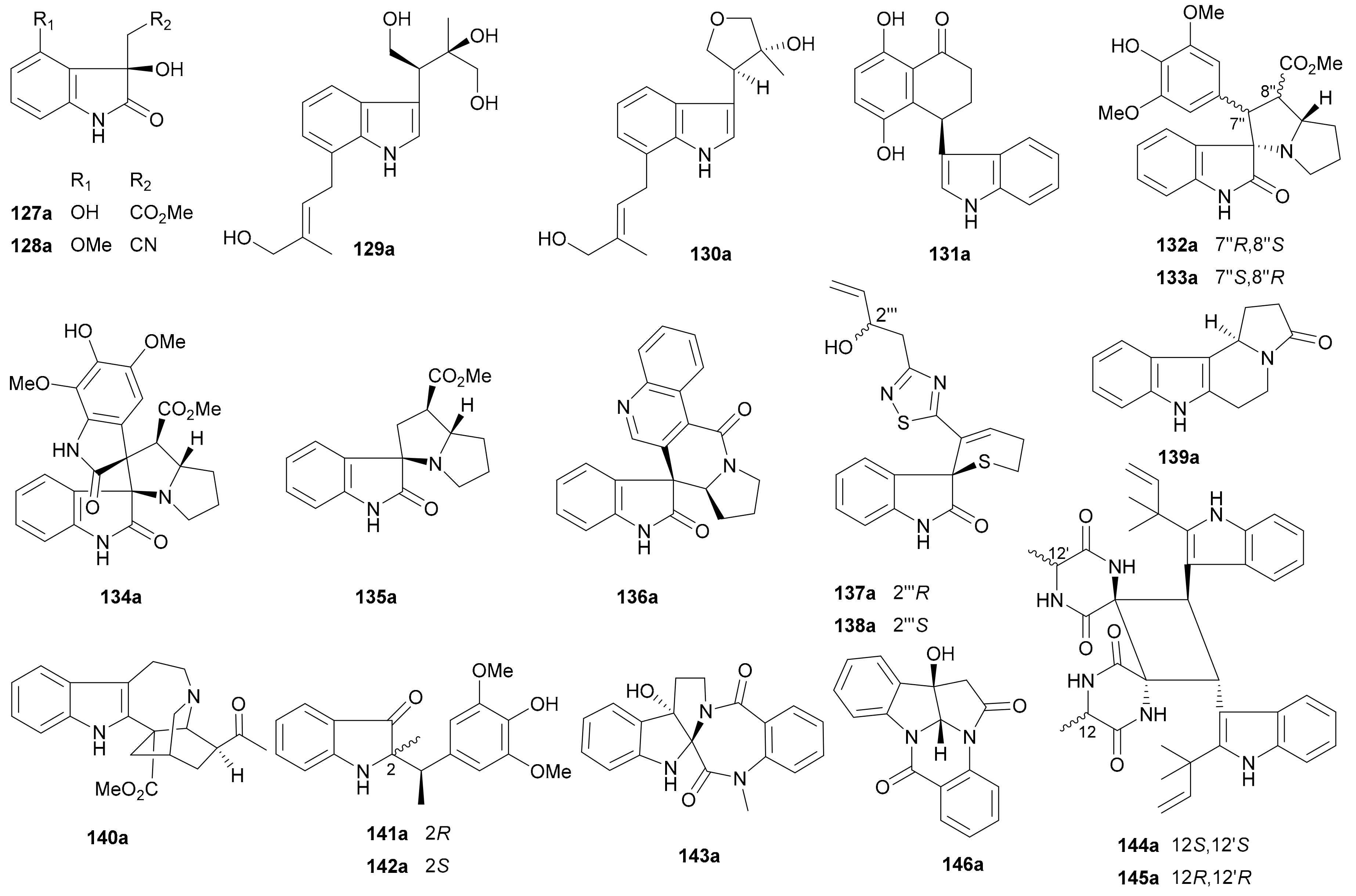{kind=link}
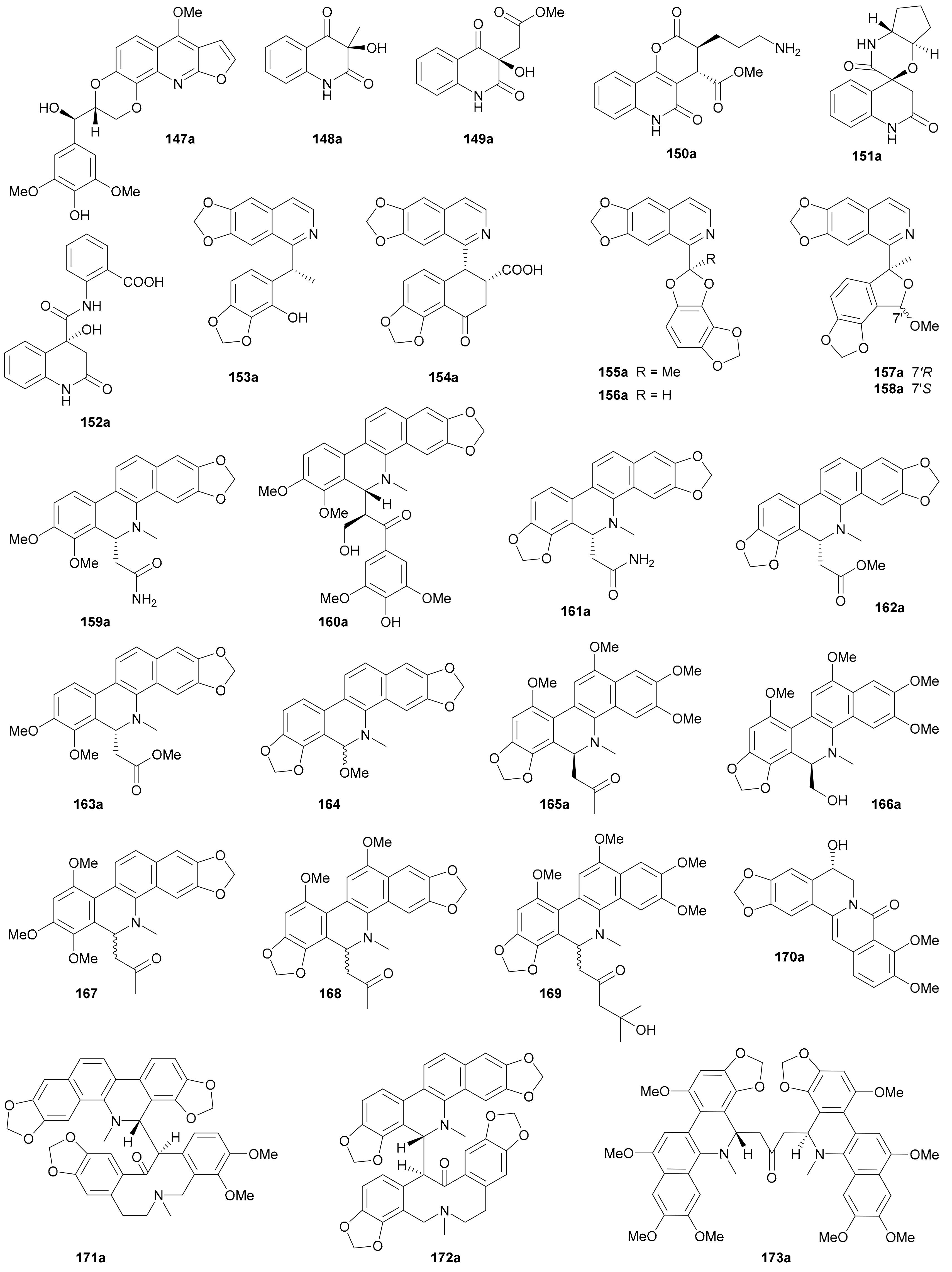{kind=link}
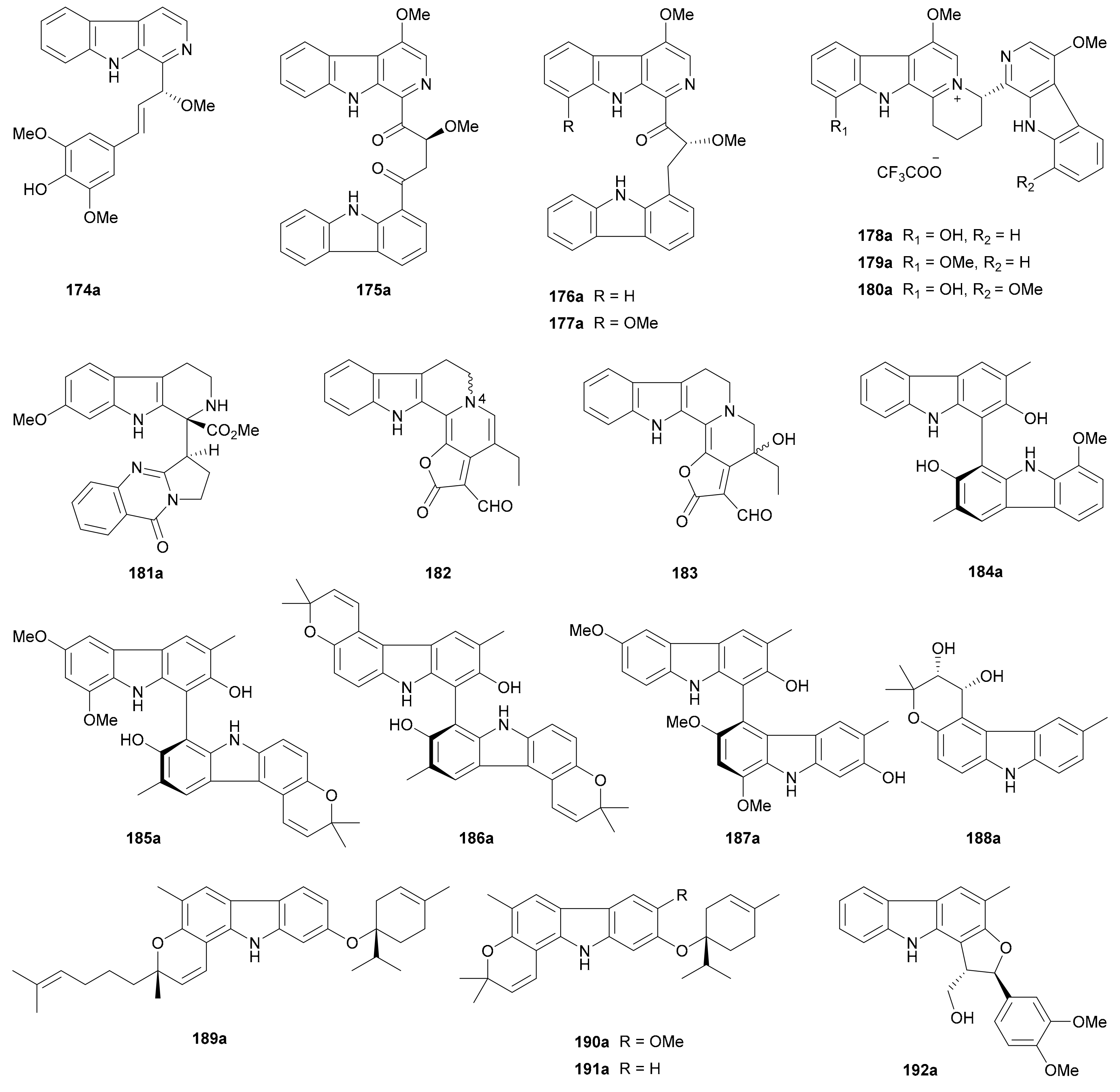{kind=link}
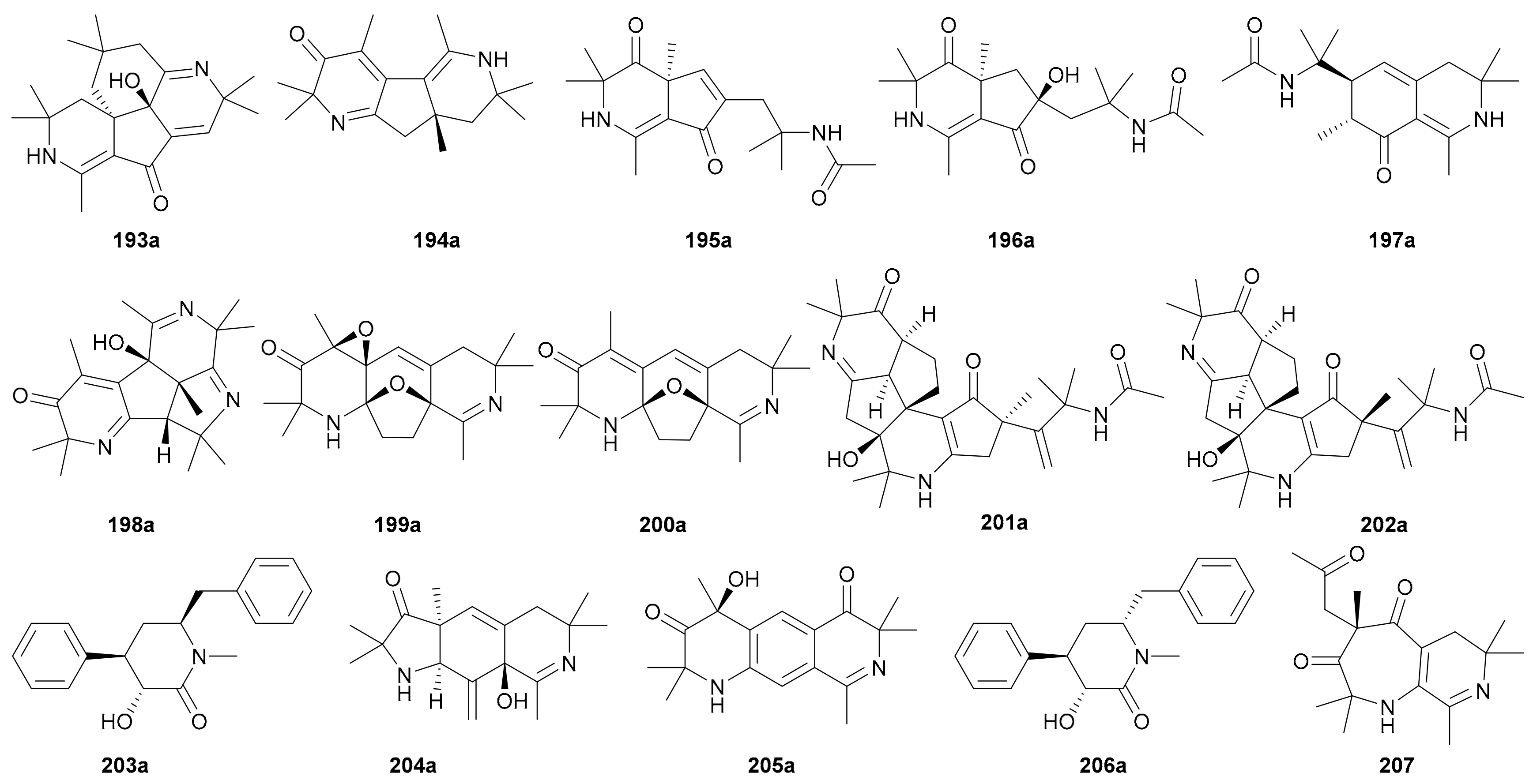{kind=link}
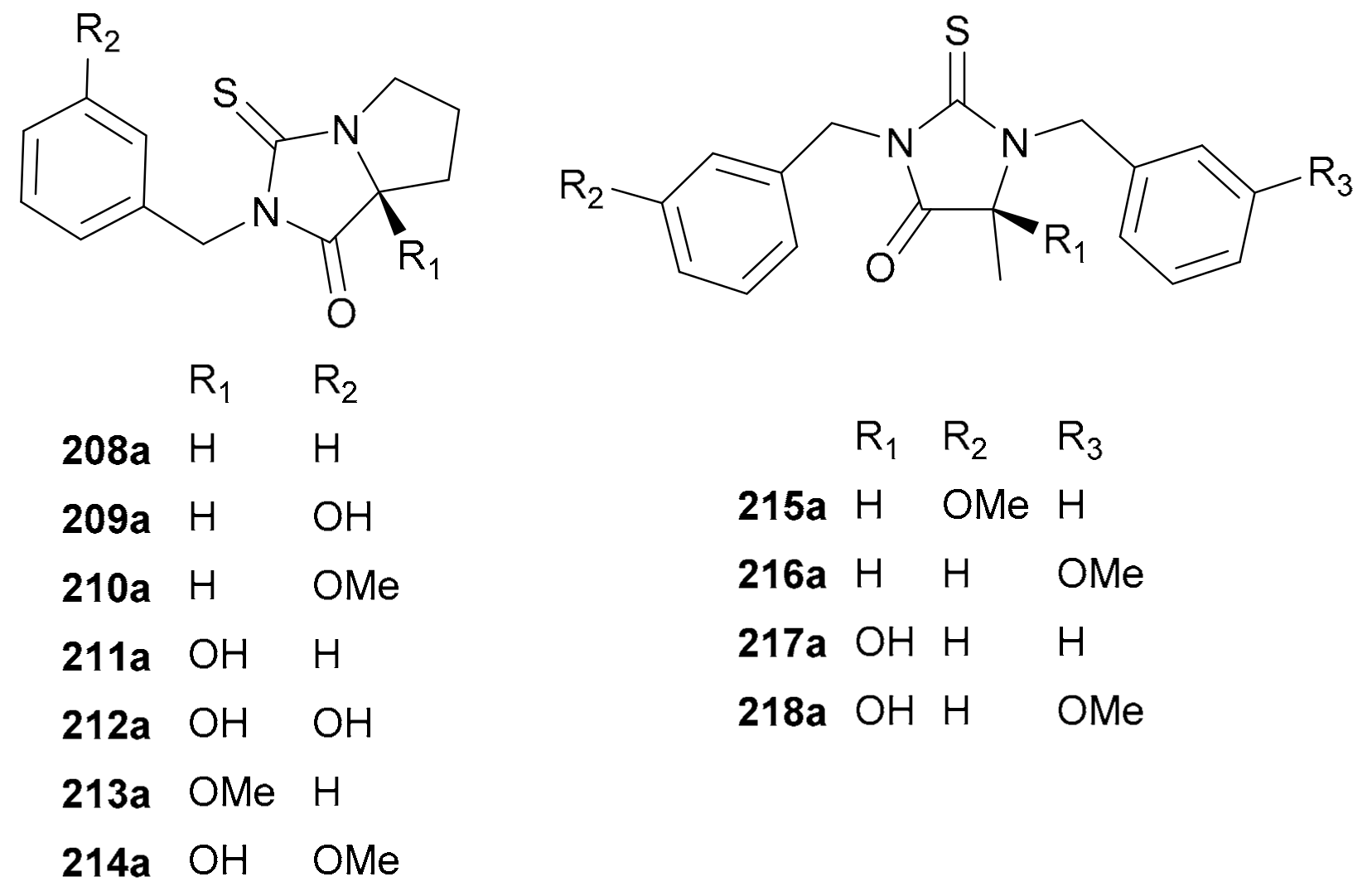{kind=link}
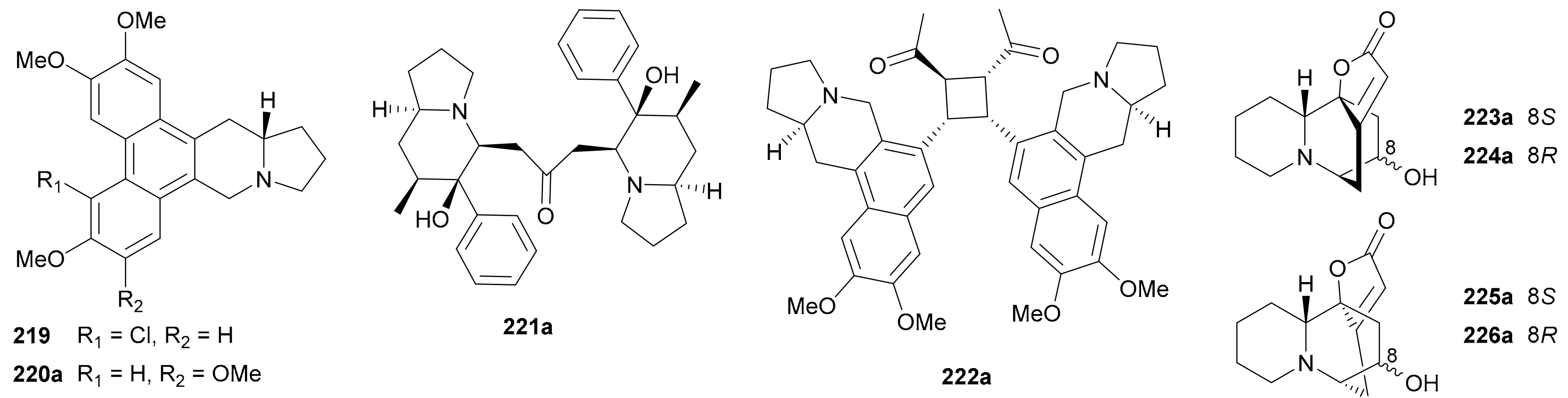{kind=link}
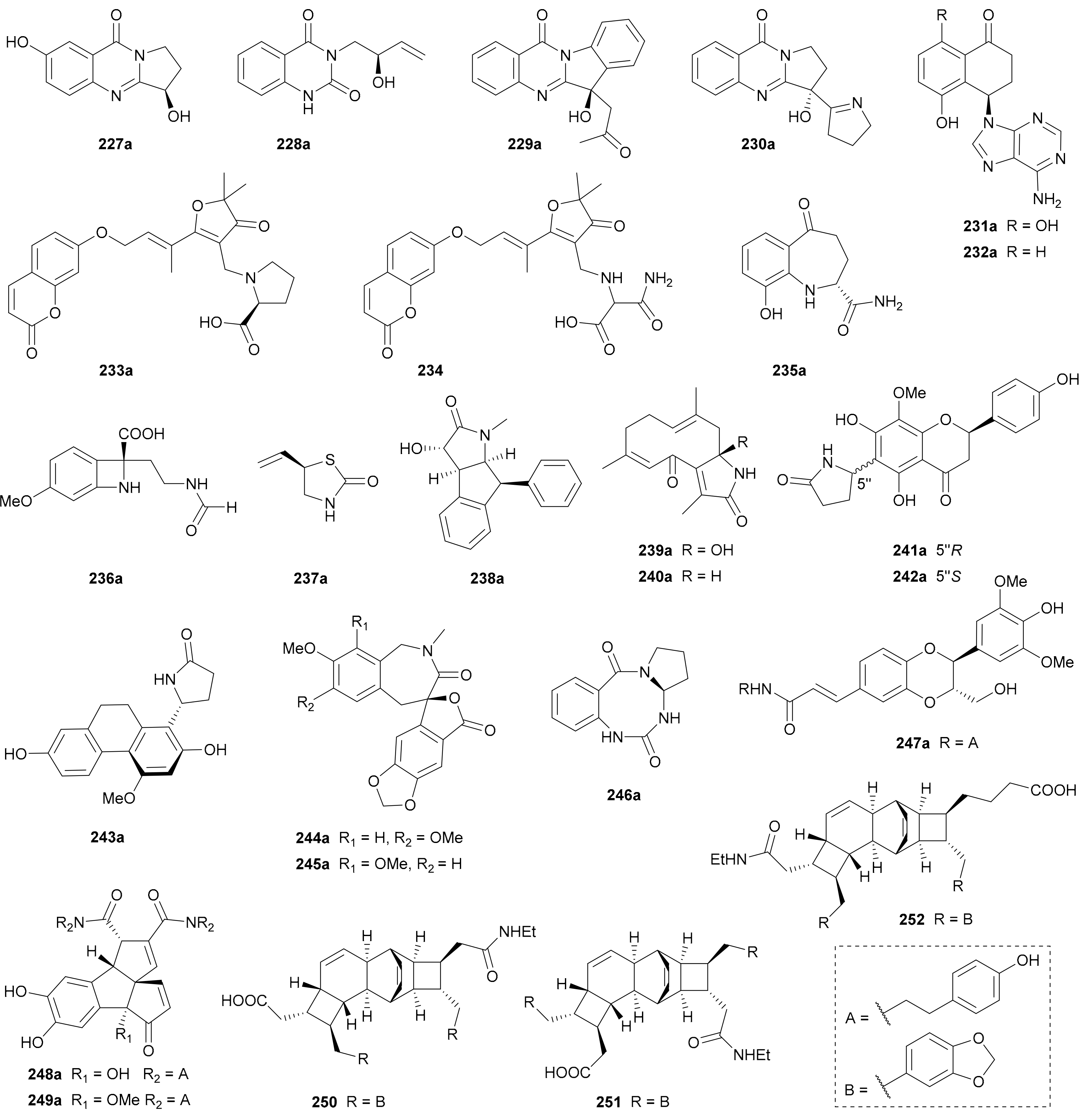{kind=link}
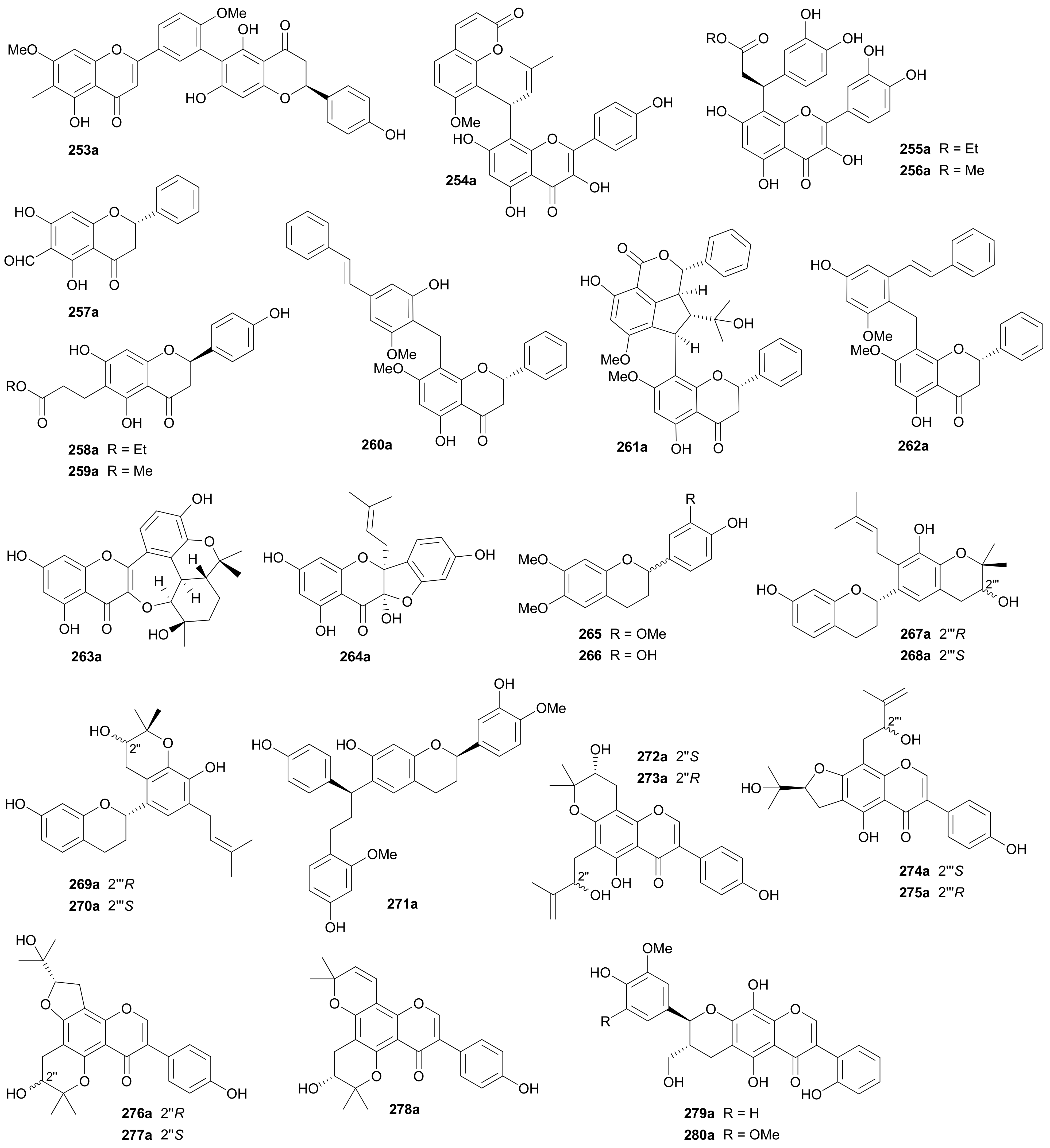{kind=link}
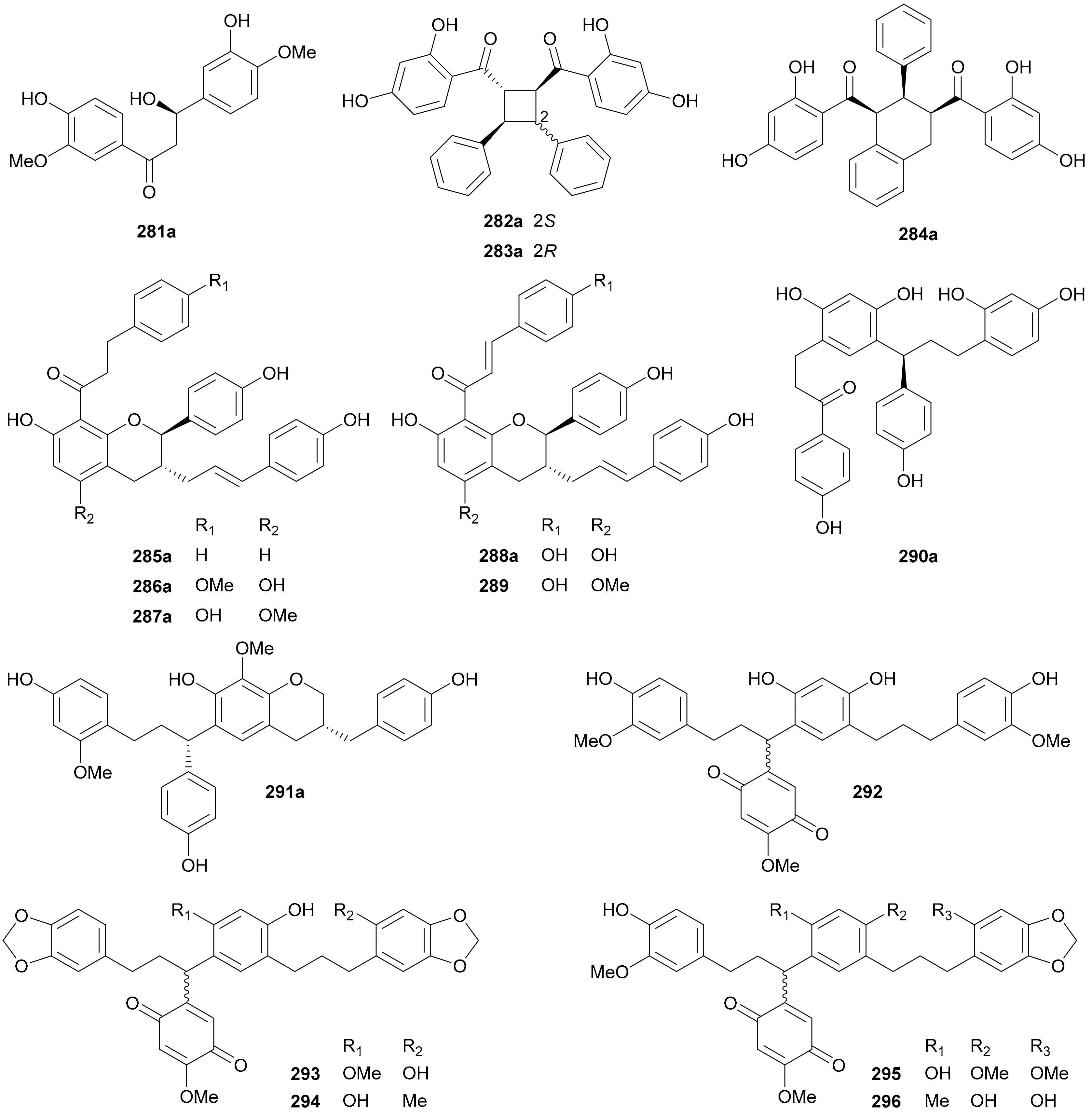{kind=link}
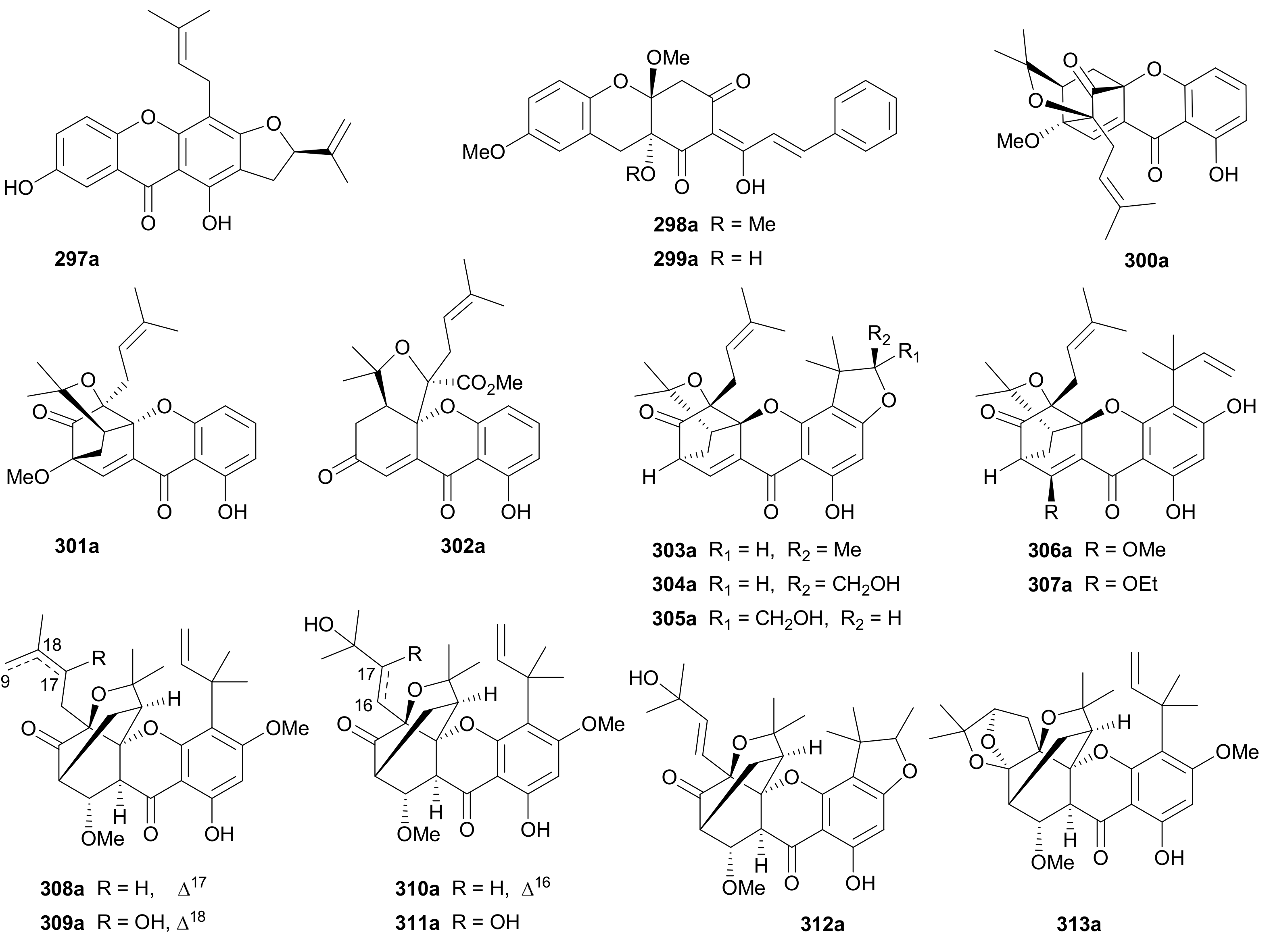{kind=link}
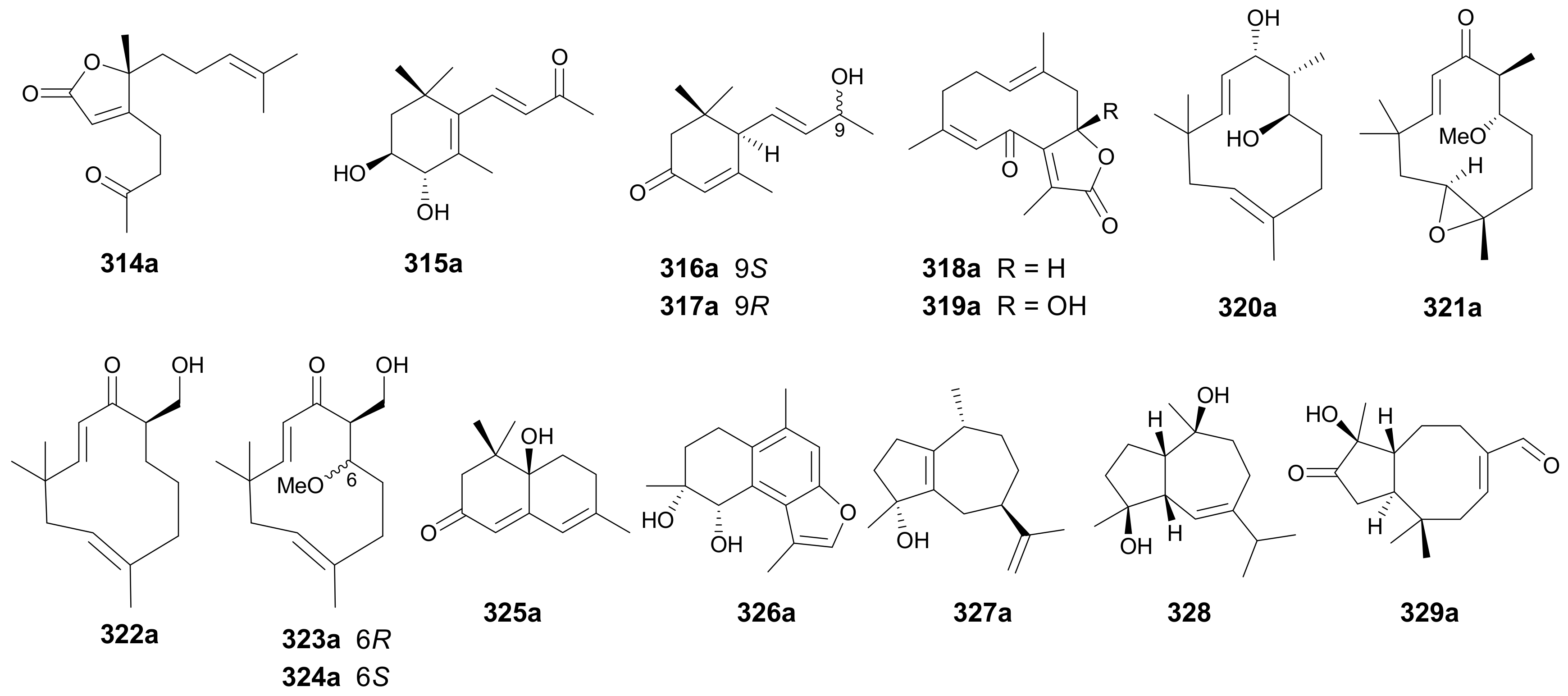{kind=link}
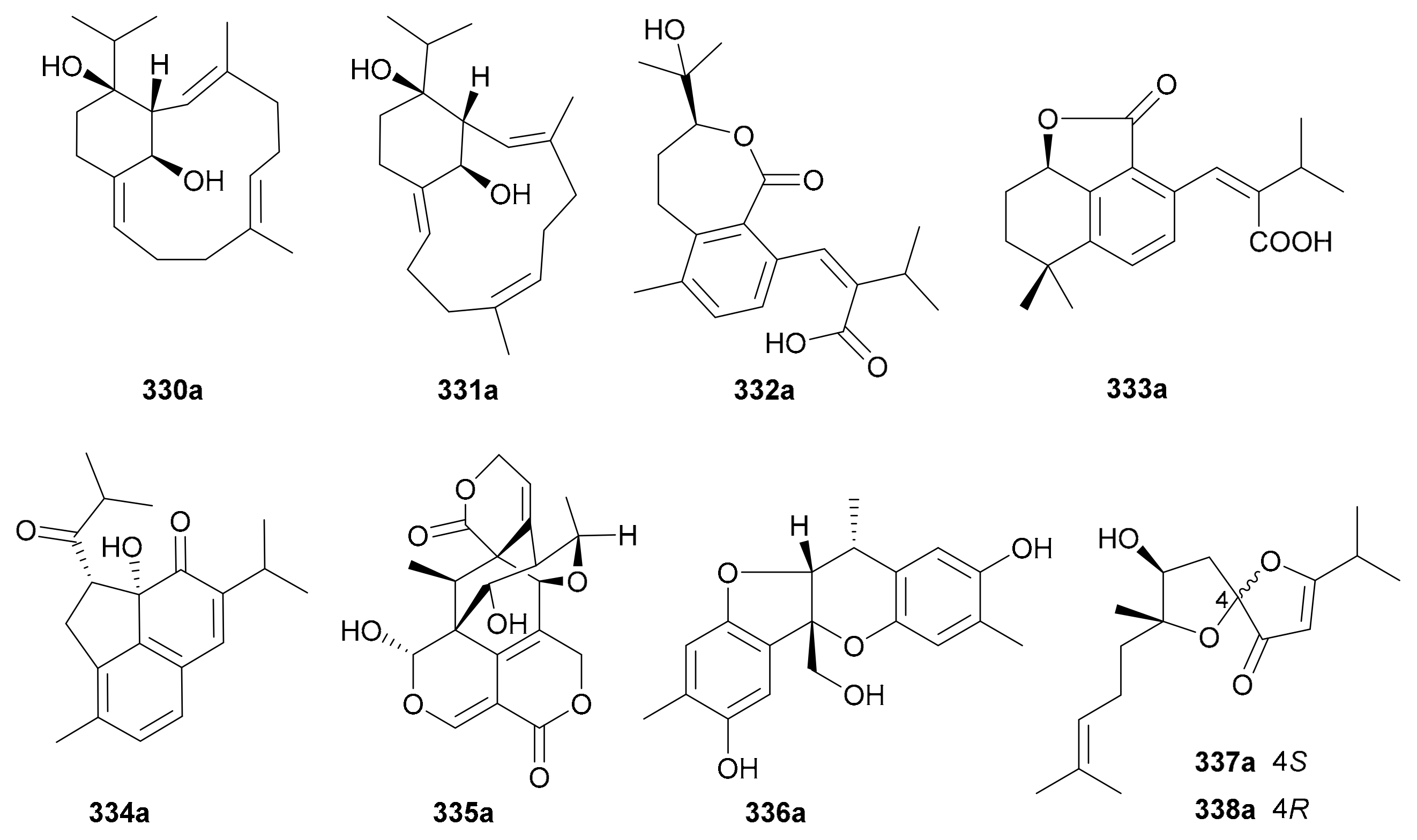{kind=link}
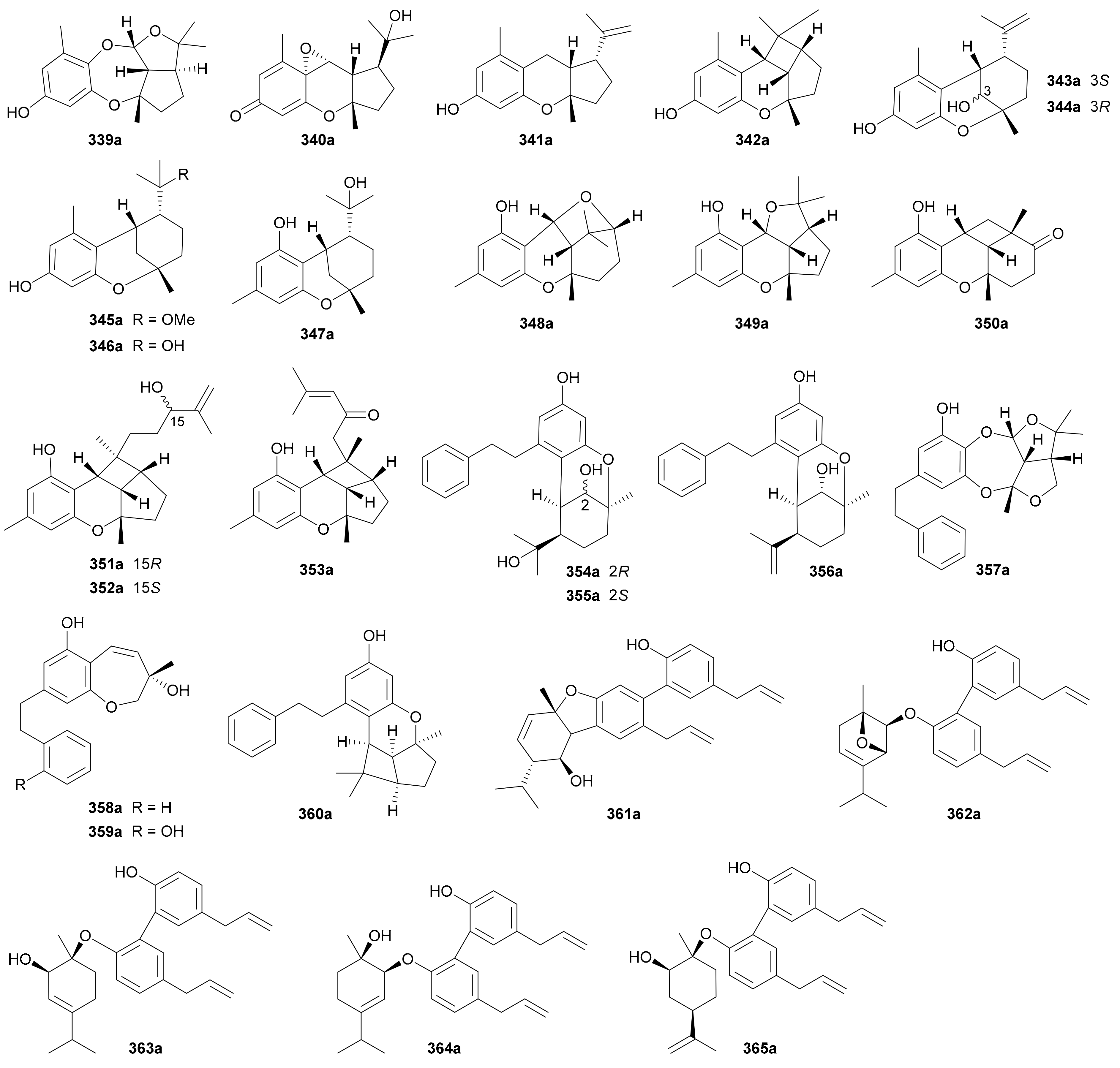{kind=link}
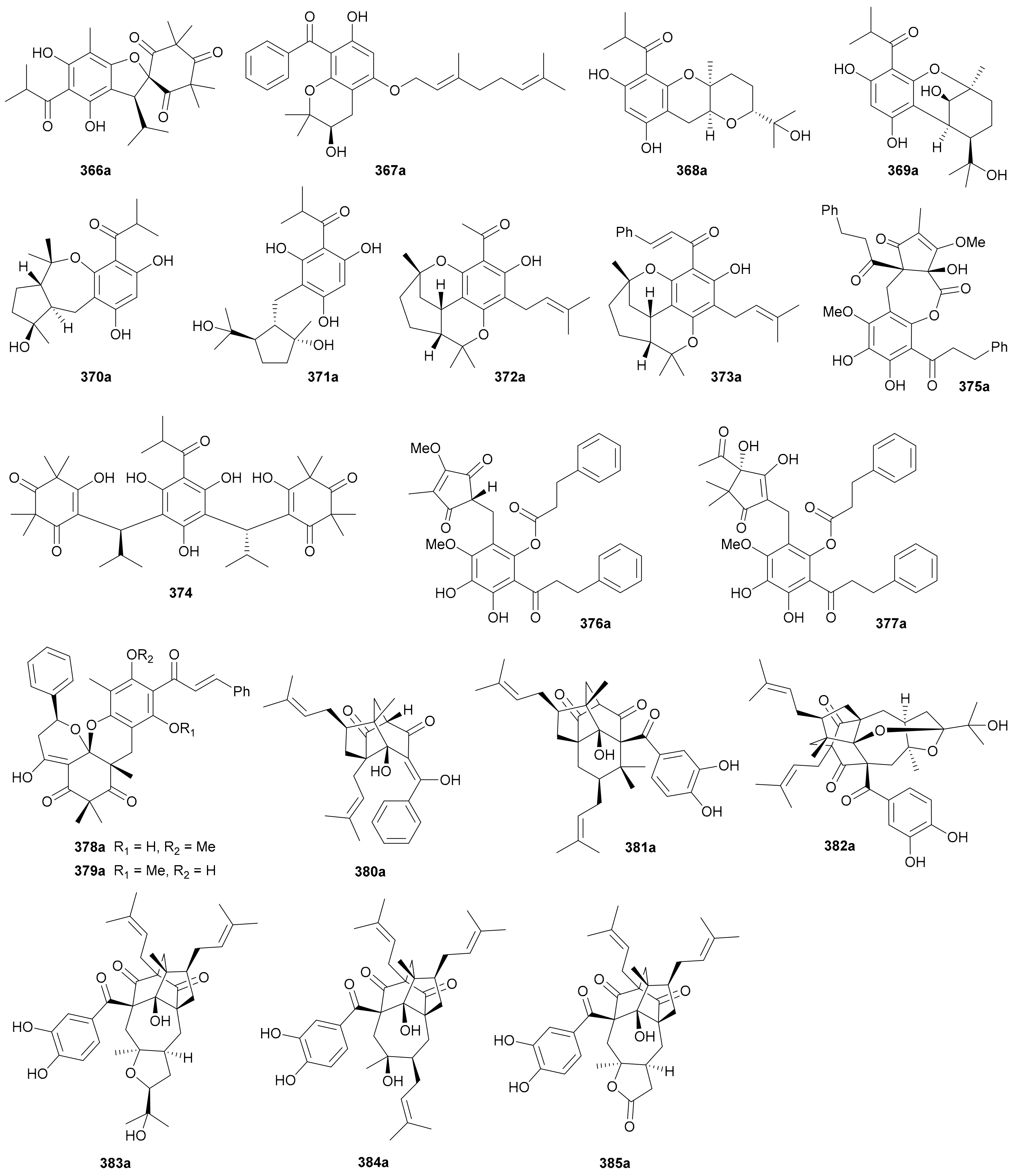{kind=link}
{kind=link}
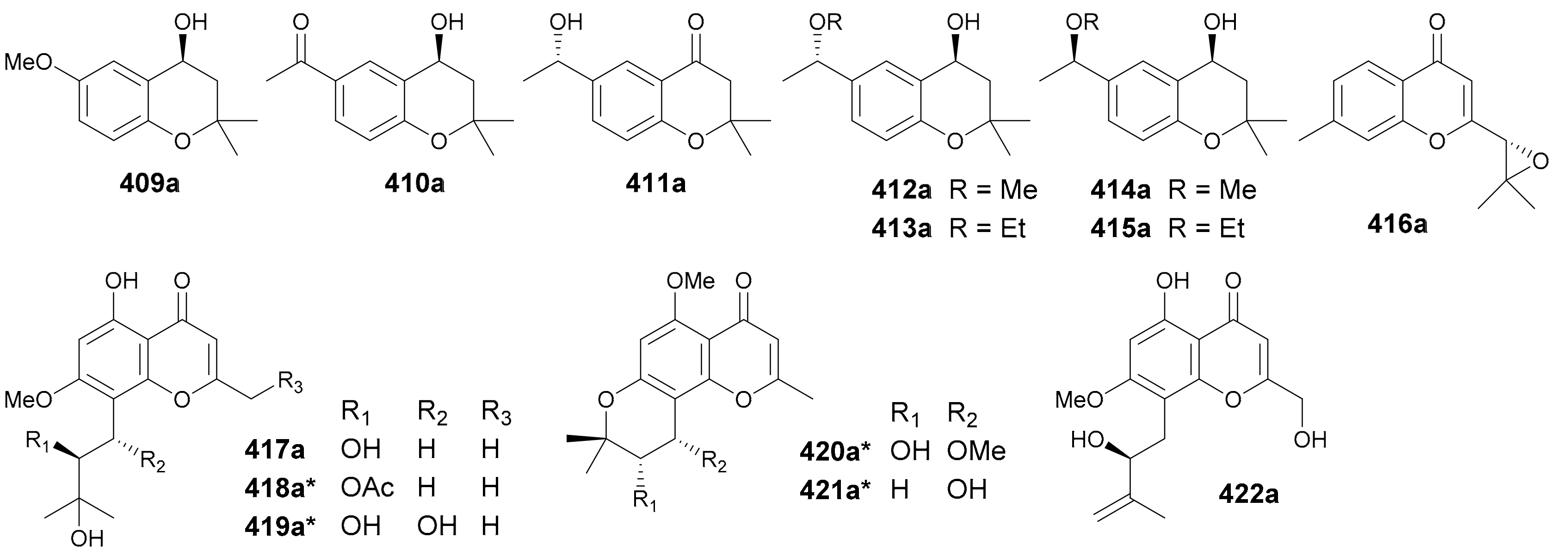{kind=link}
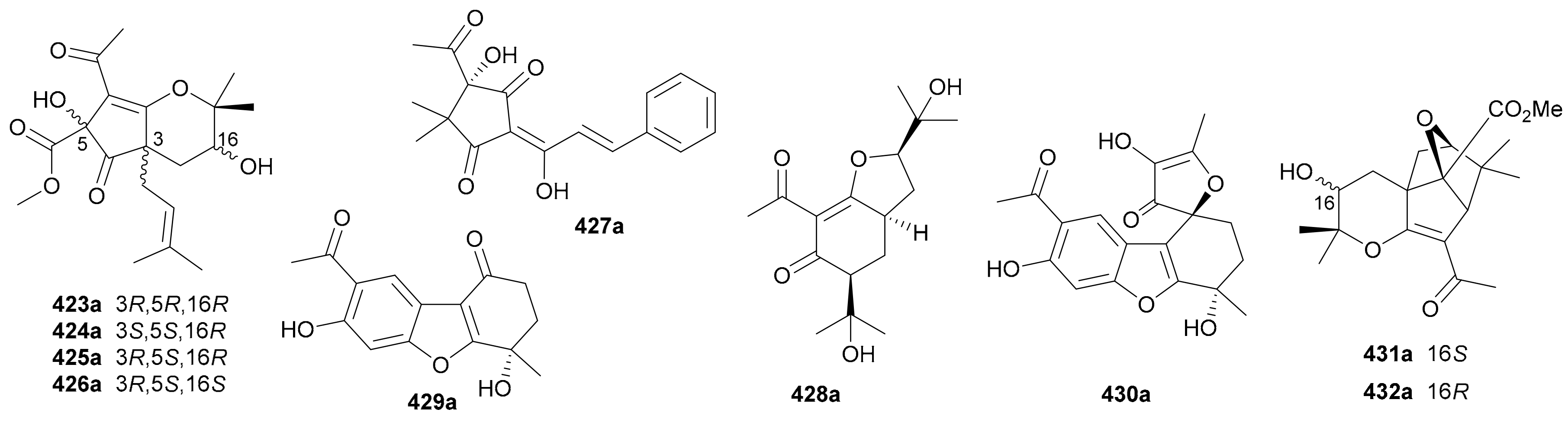{kind=link}
{kind=link}
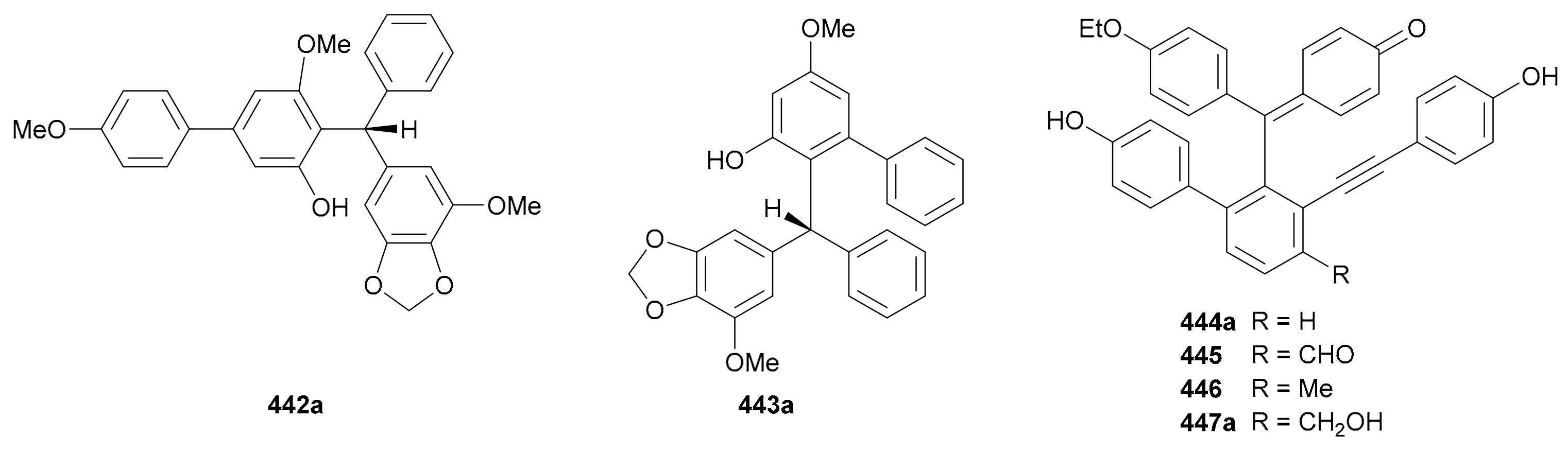{kind=link}
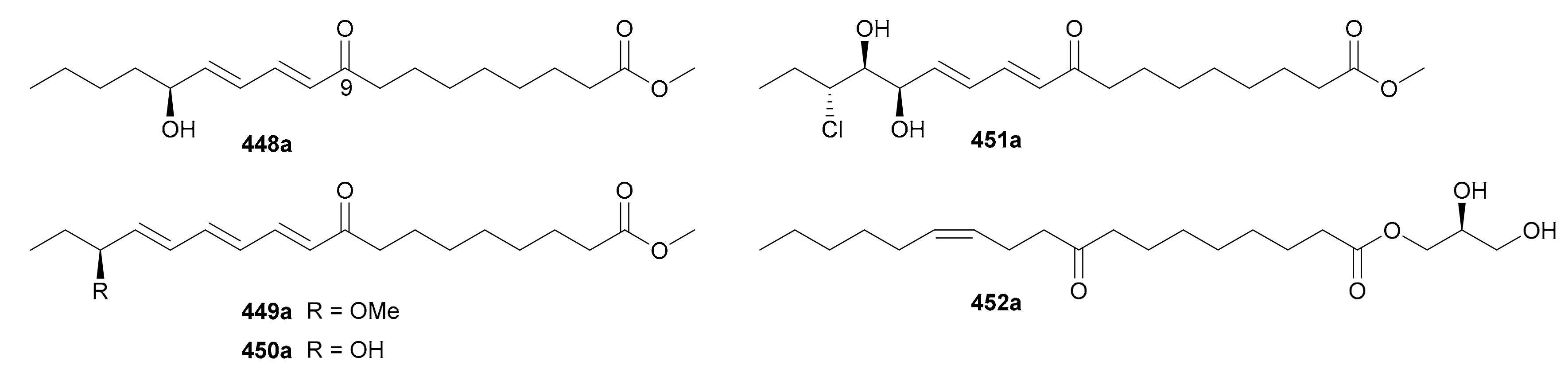{kind=link}
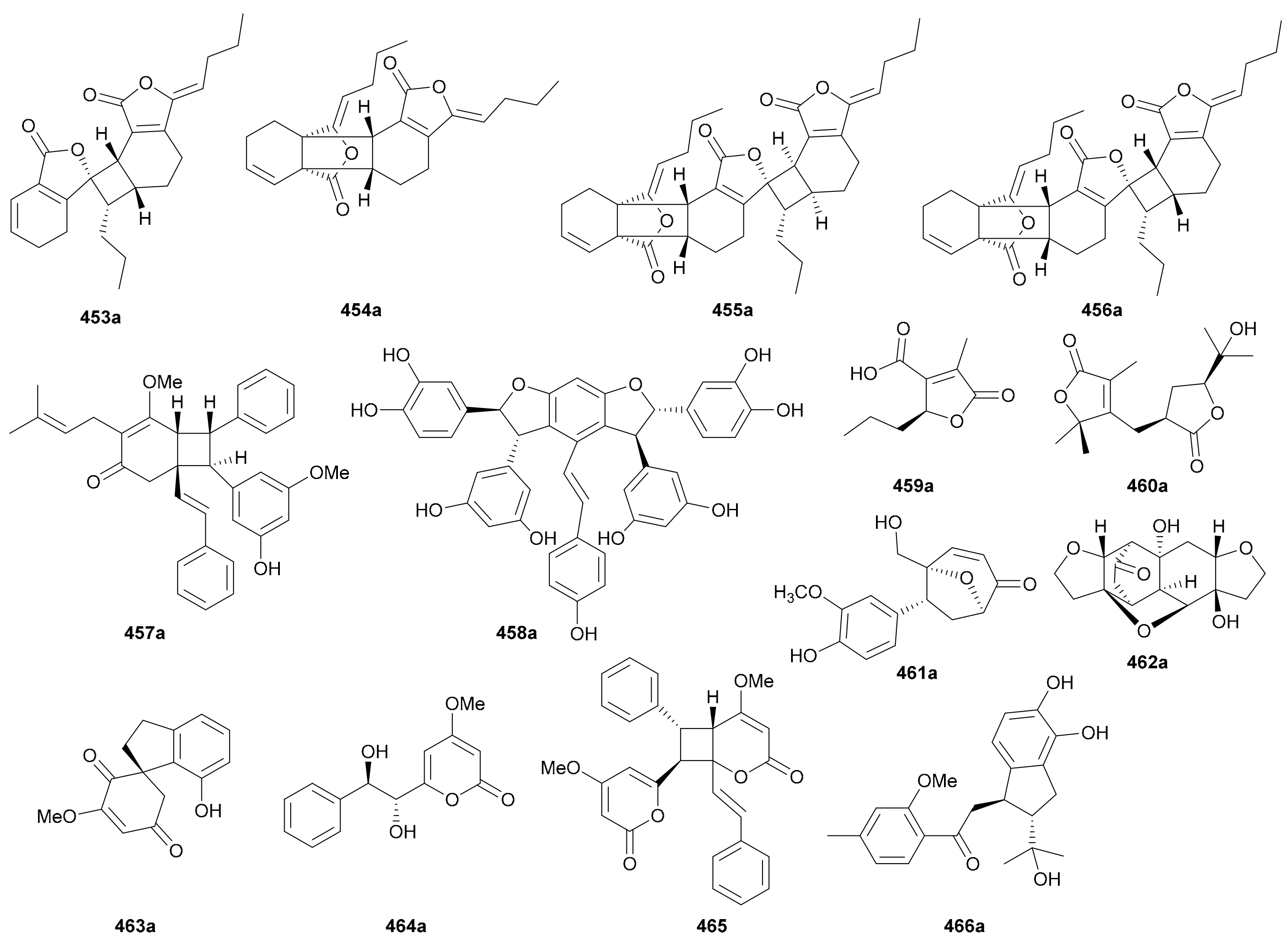{kind=link}
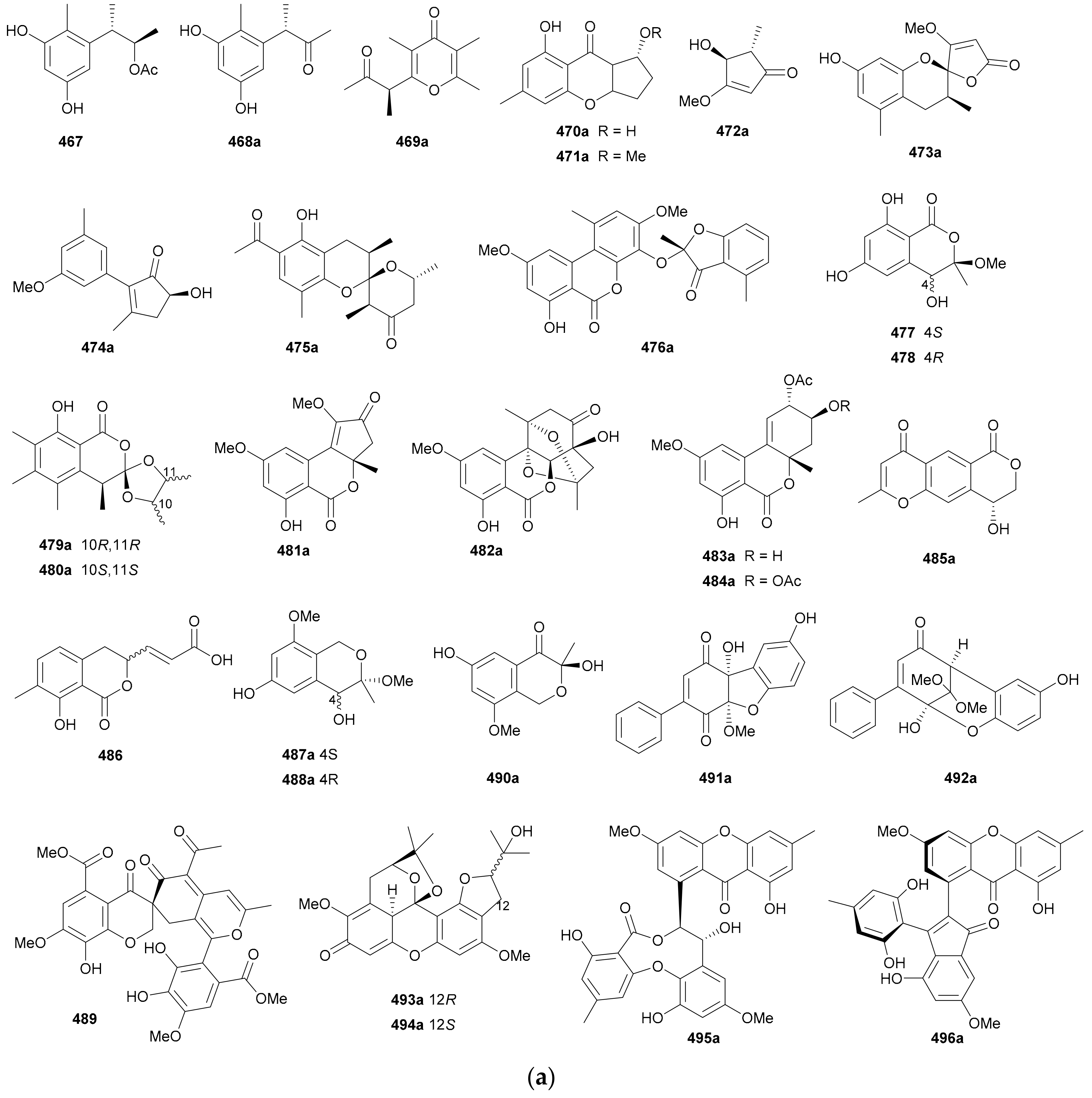{kind=link}
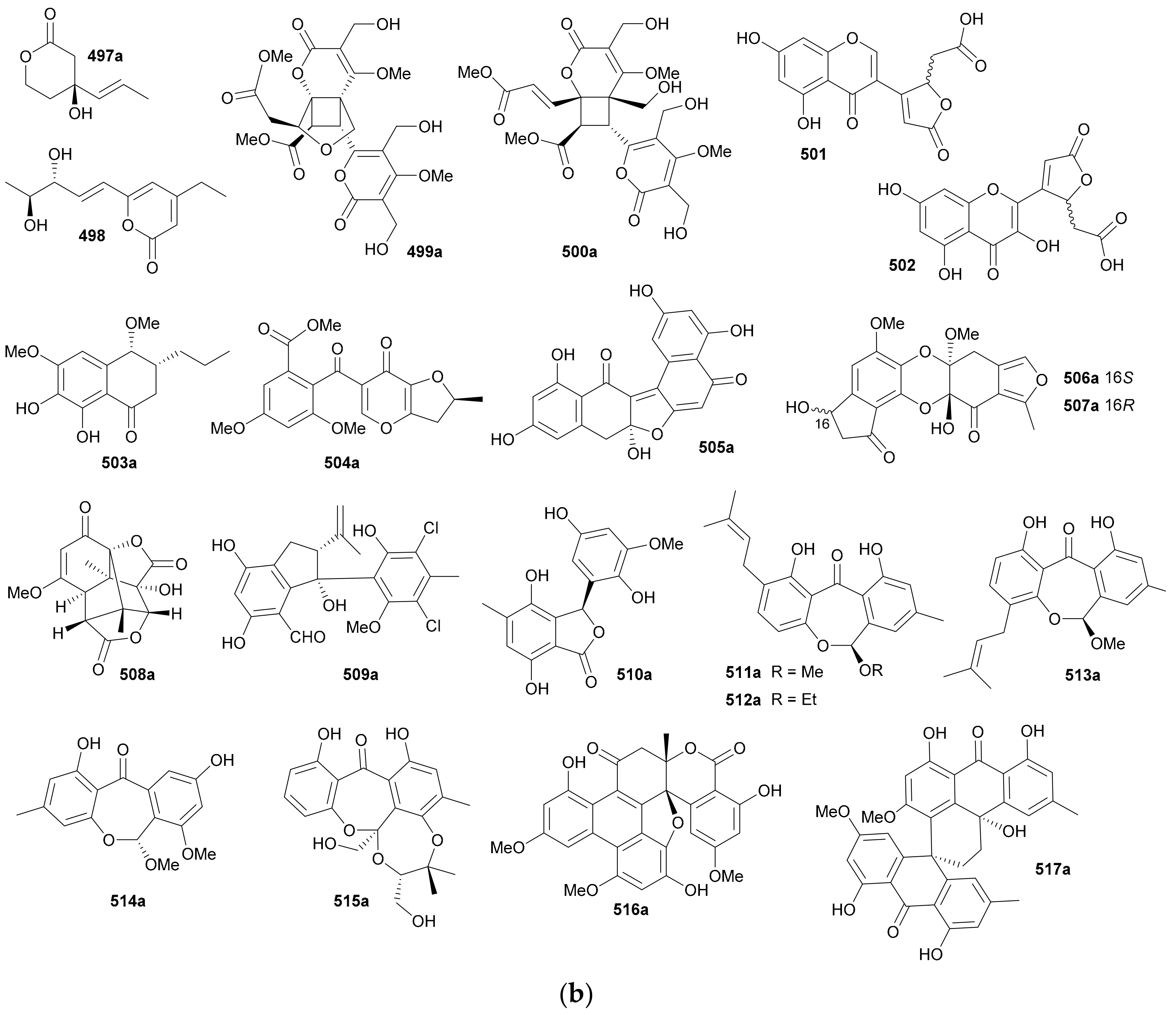{kind=link}
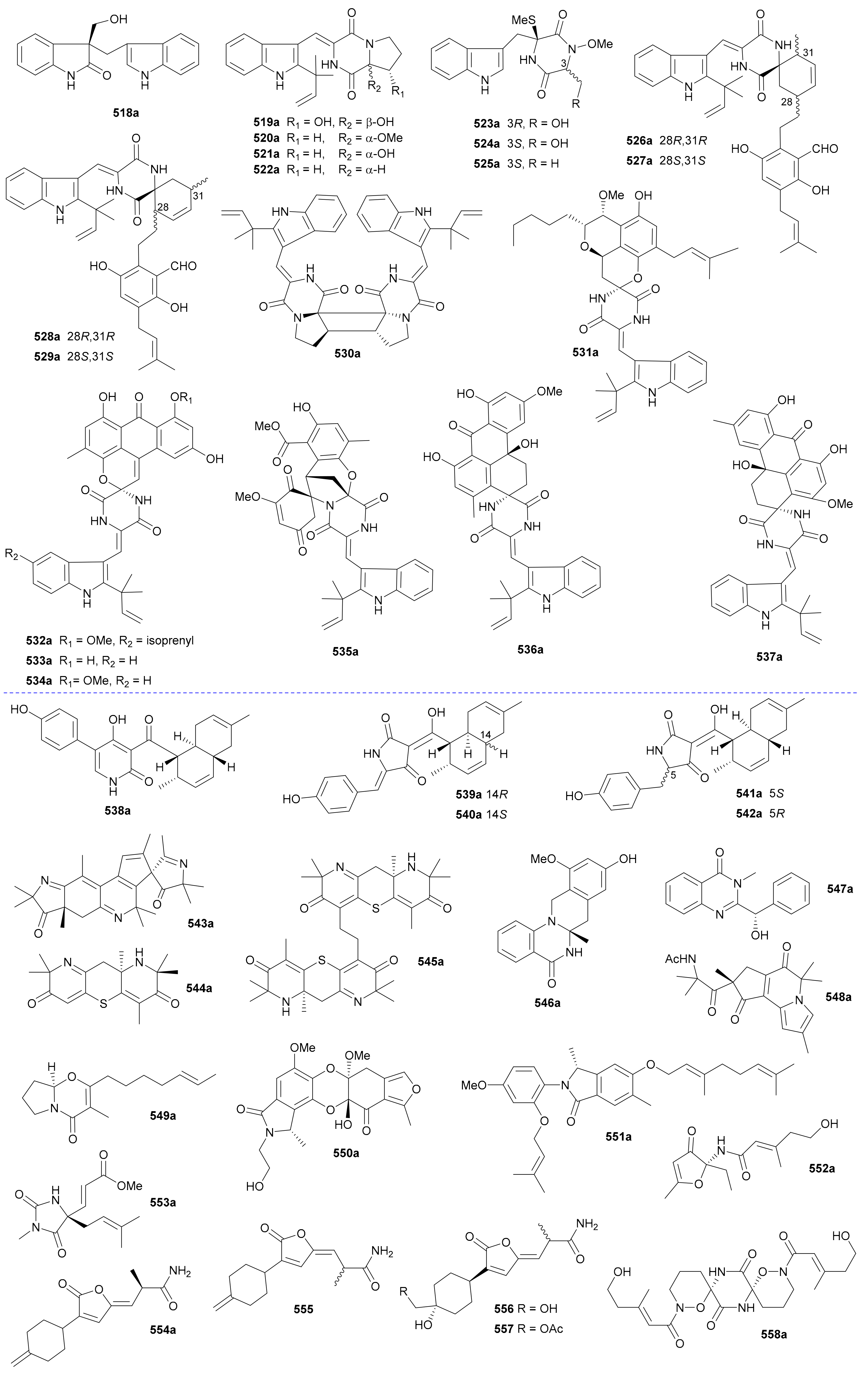{kind=link}
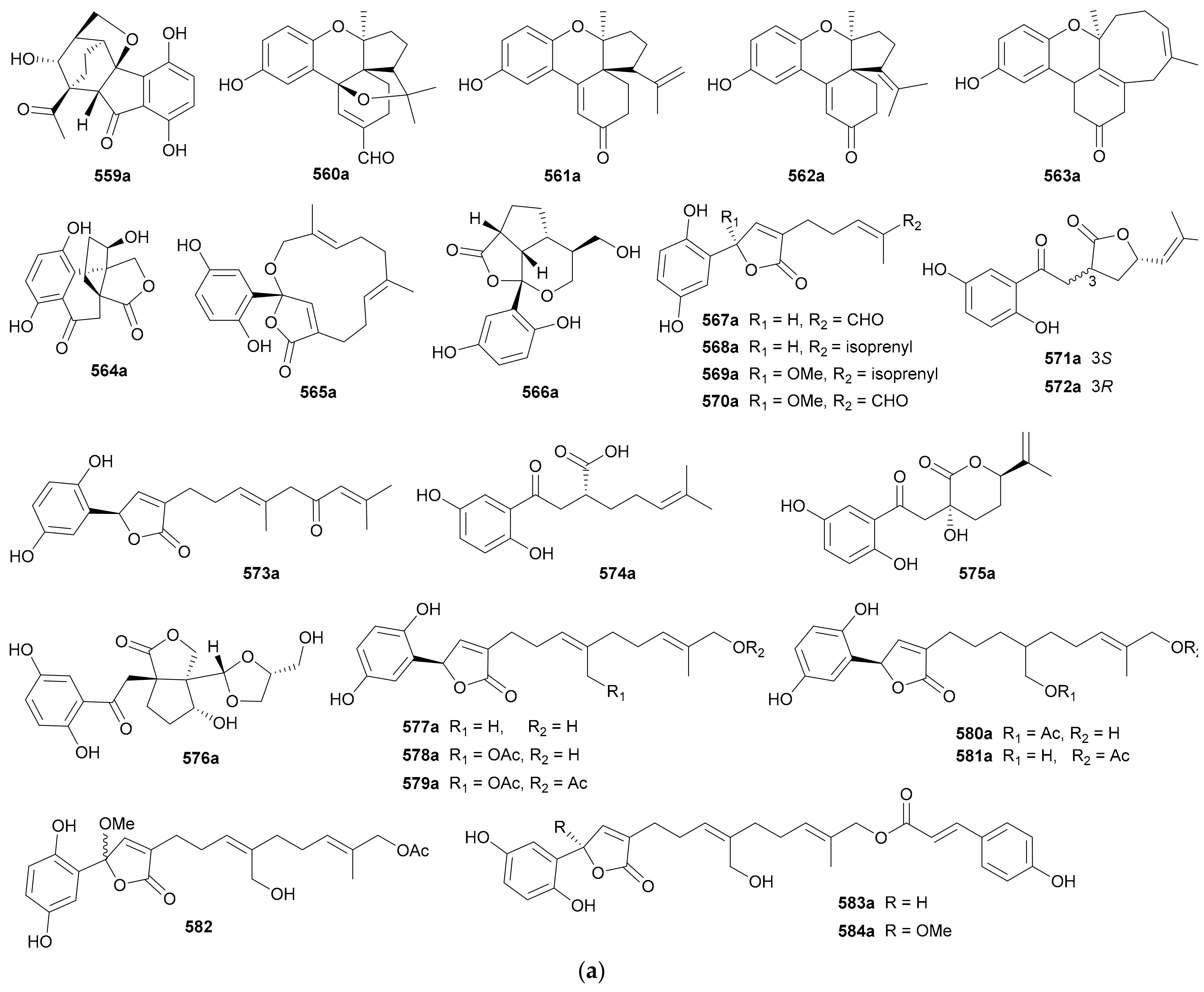{kind=link}
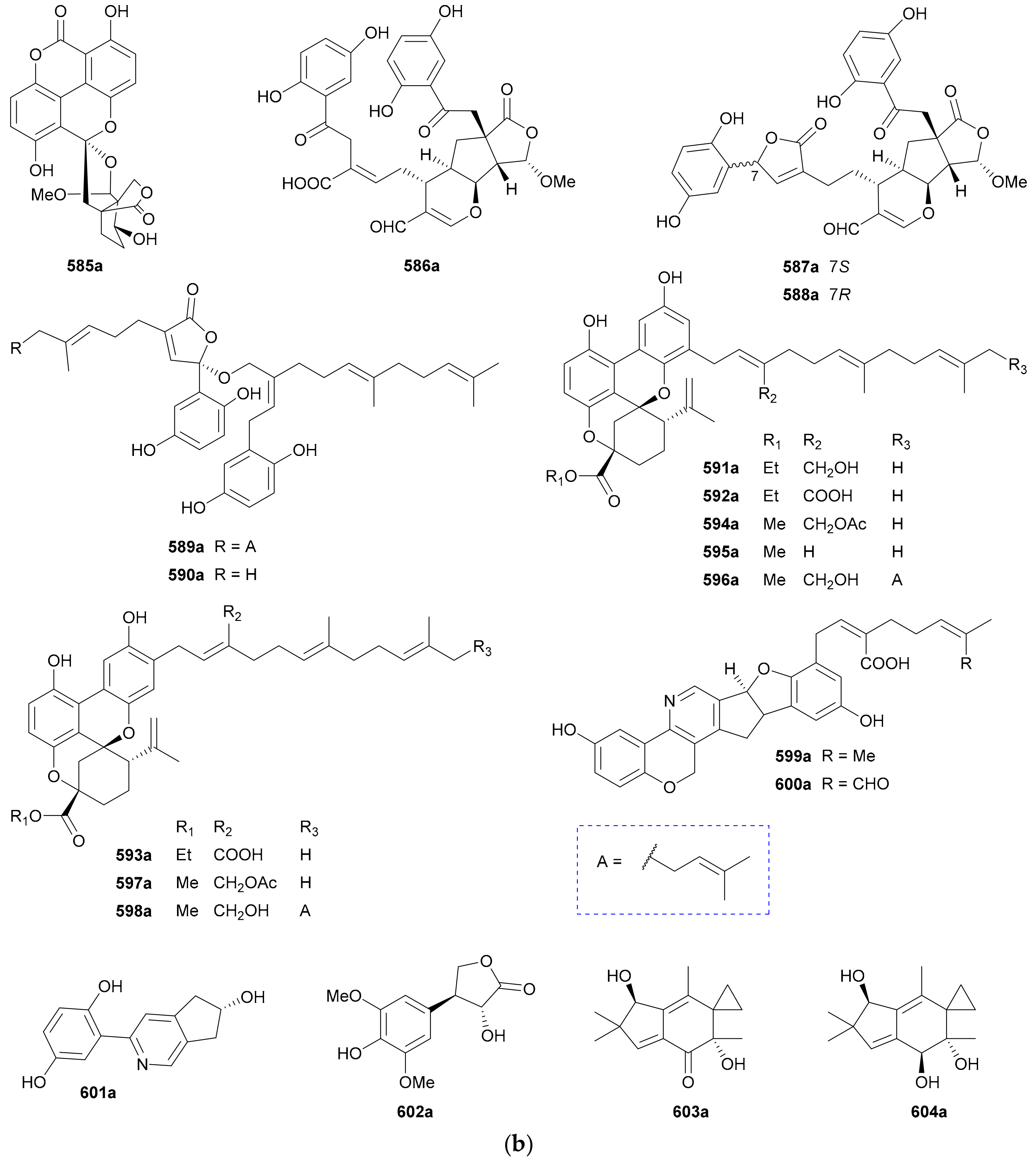{kind=link}
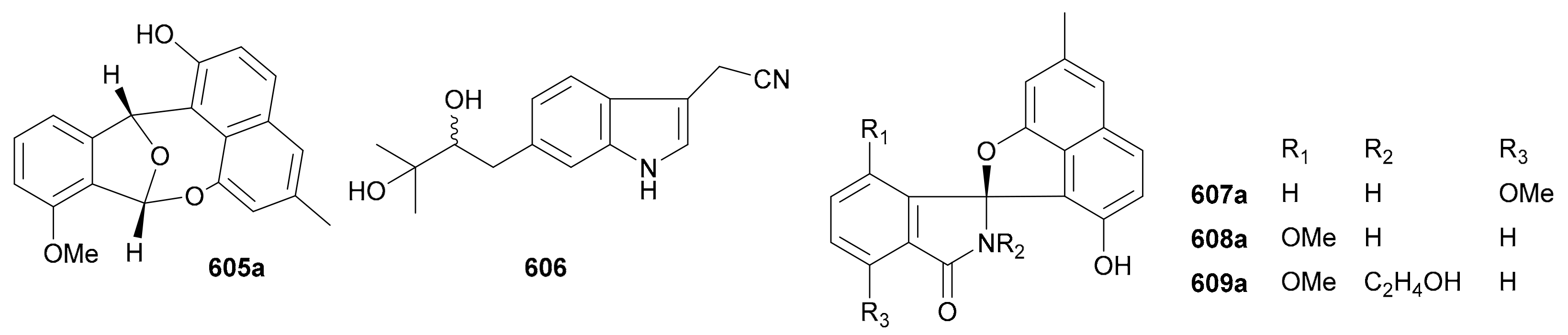{kind=link}
{kind=link}
{kind=link}
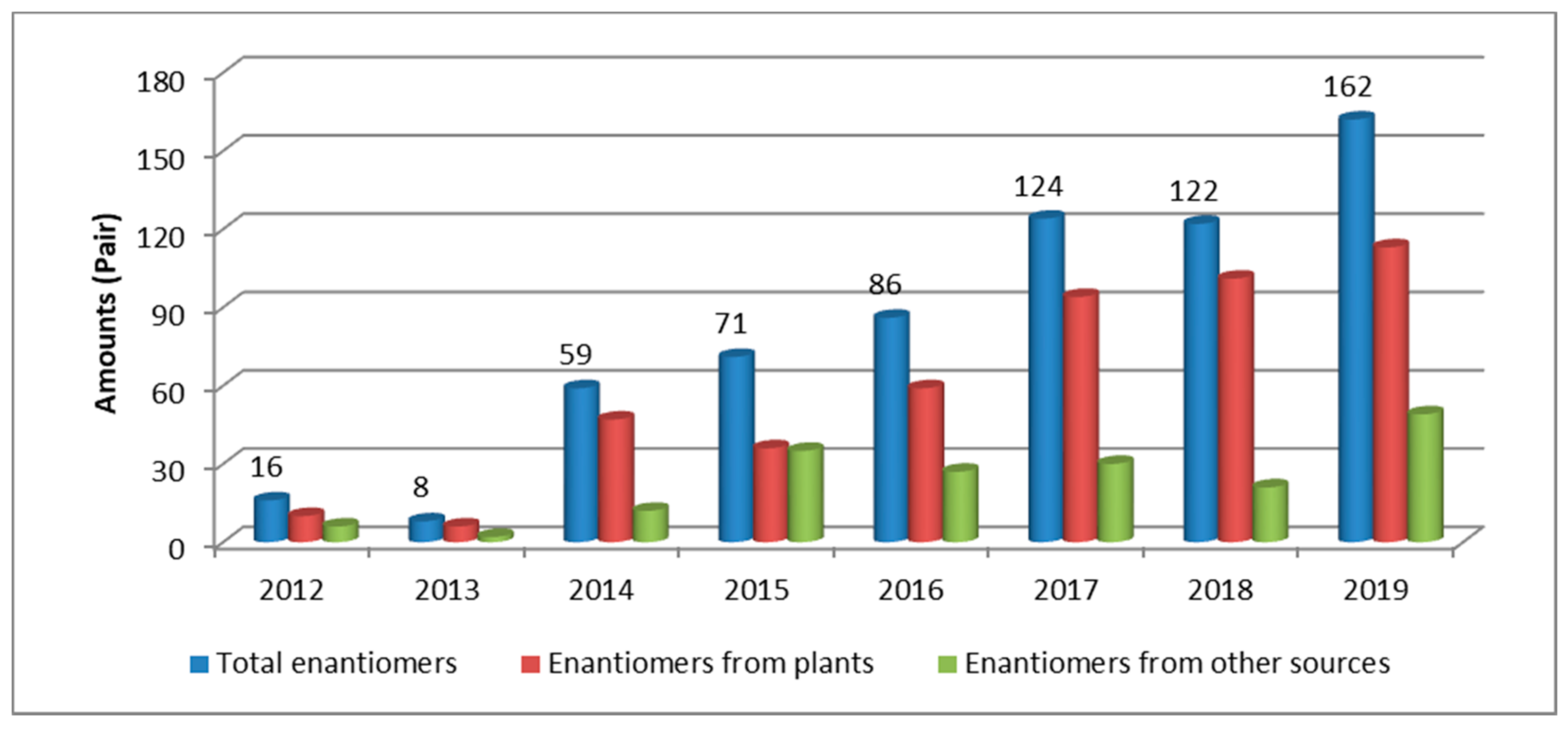{kind=link}
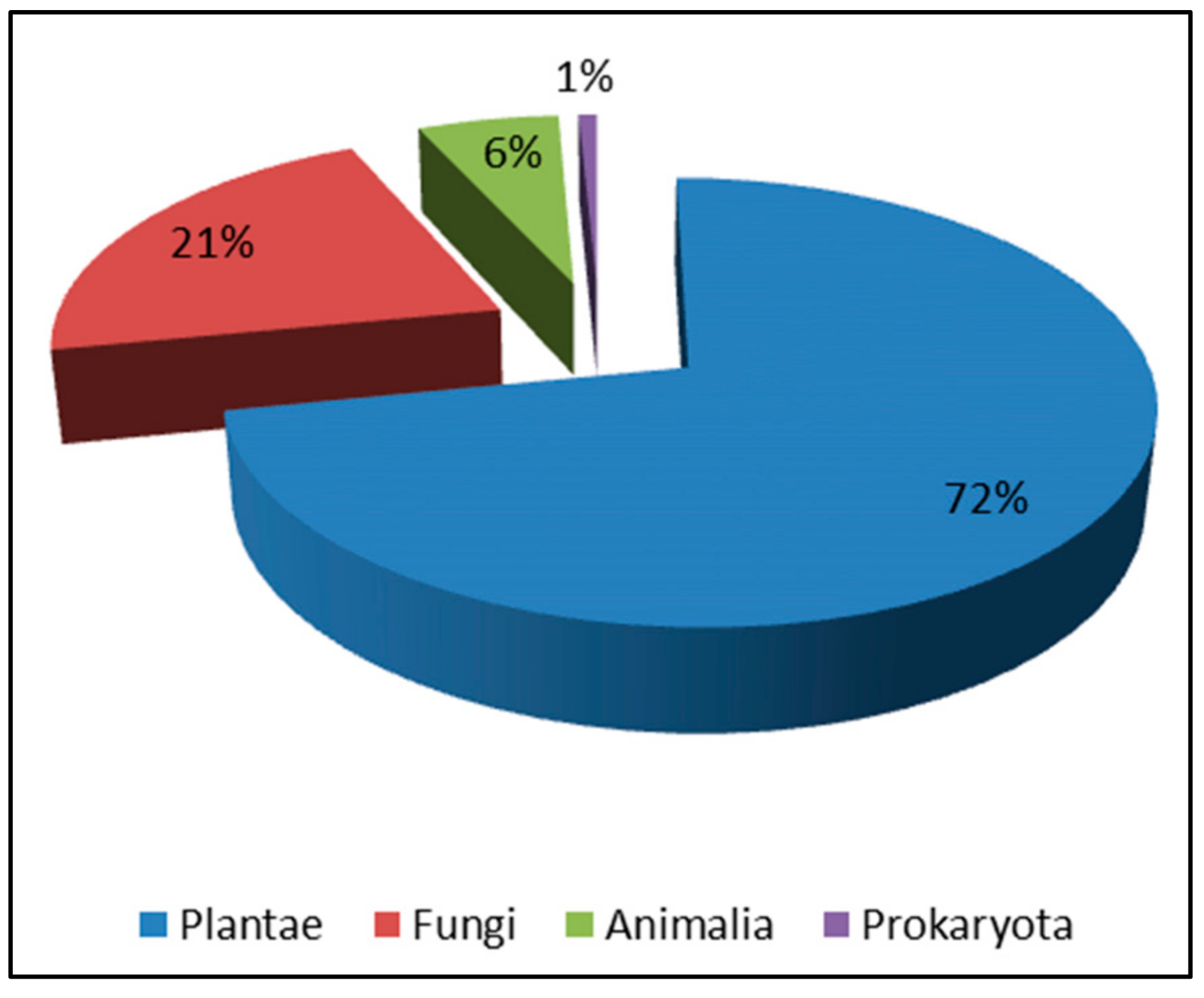{kind=link}
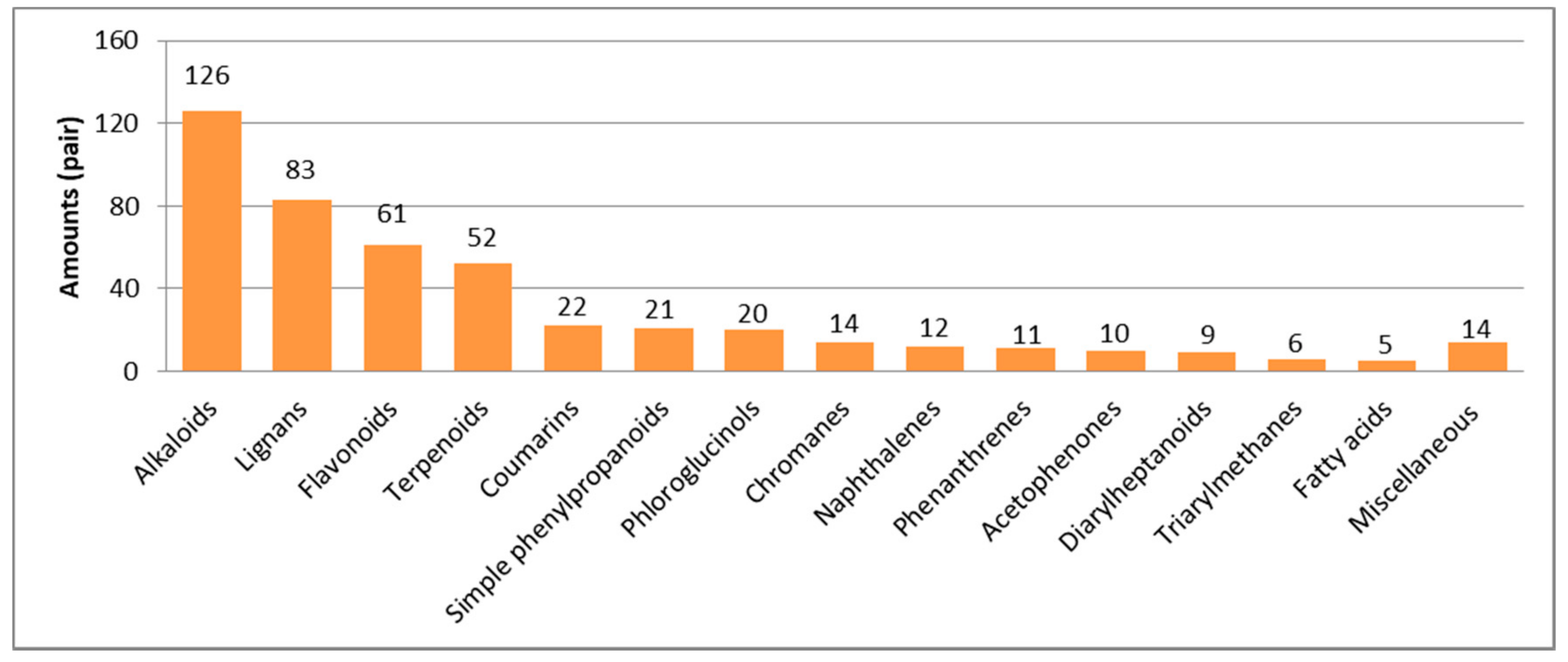{kind=link}
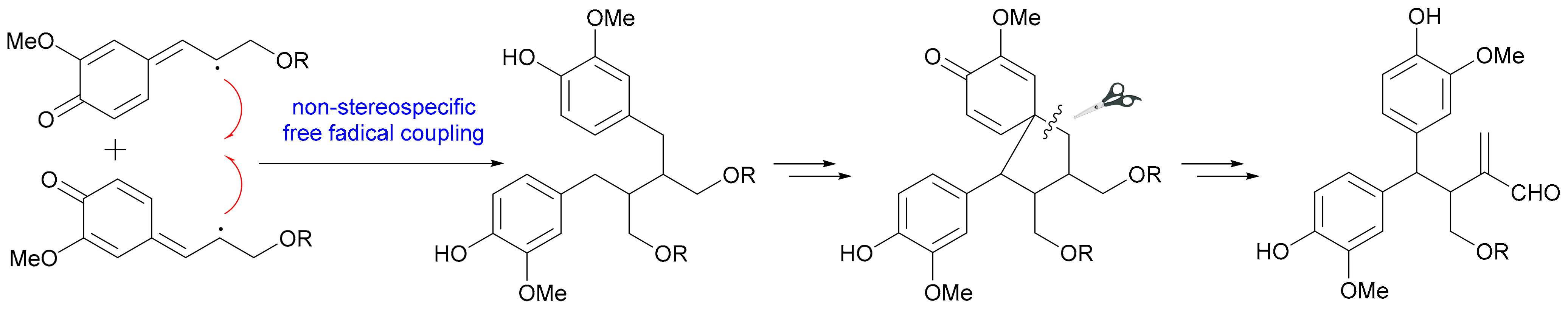{kind=link}
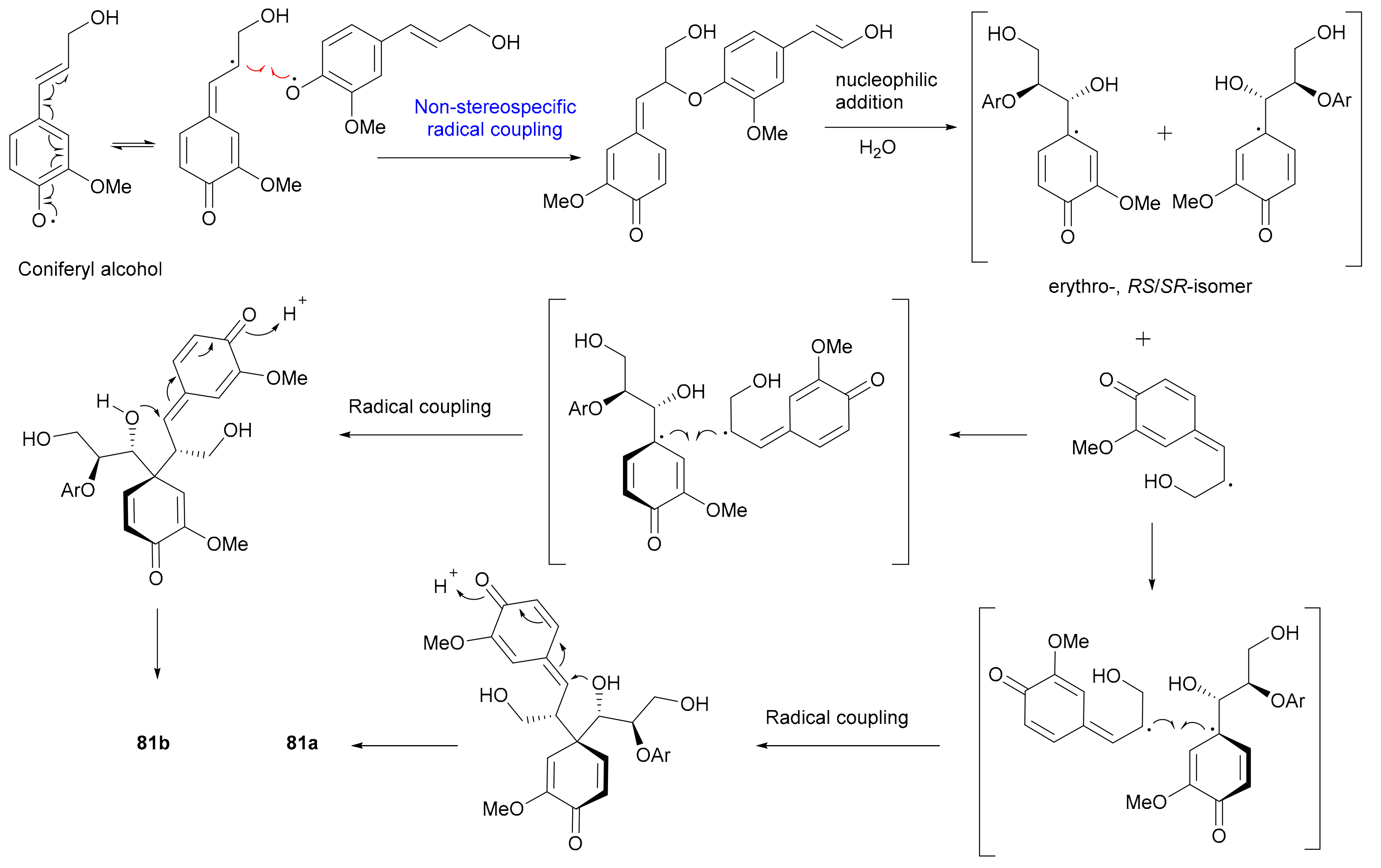{kind=link}
{kind=link}
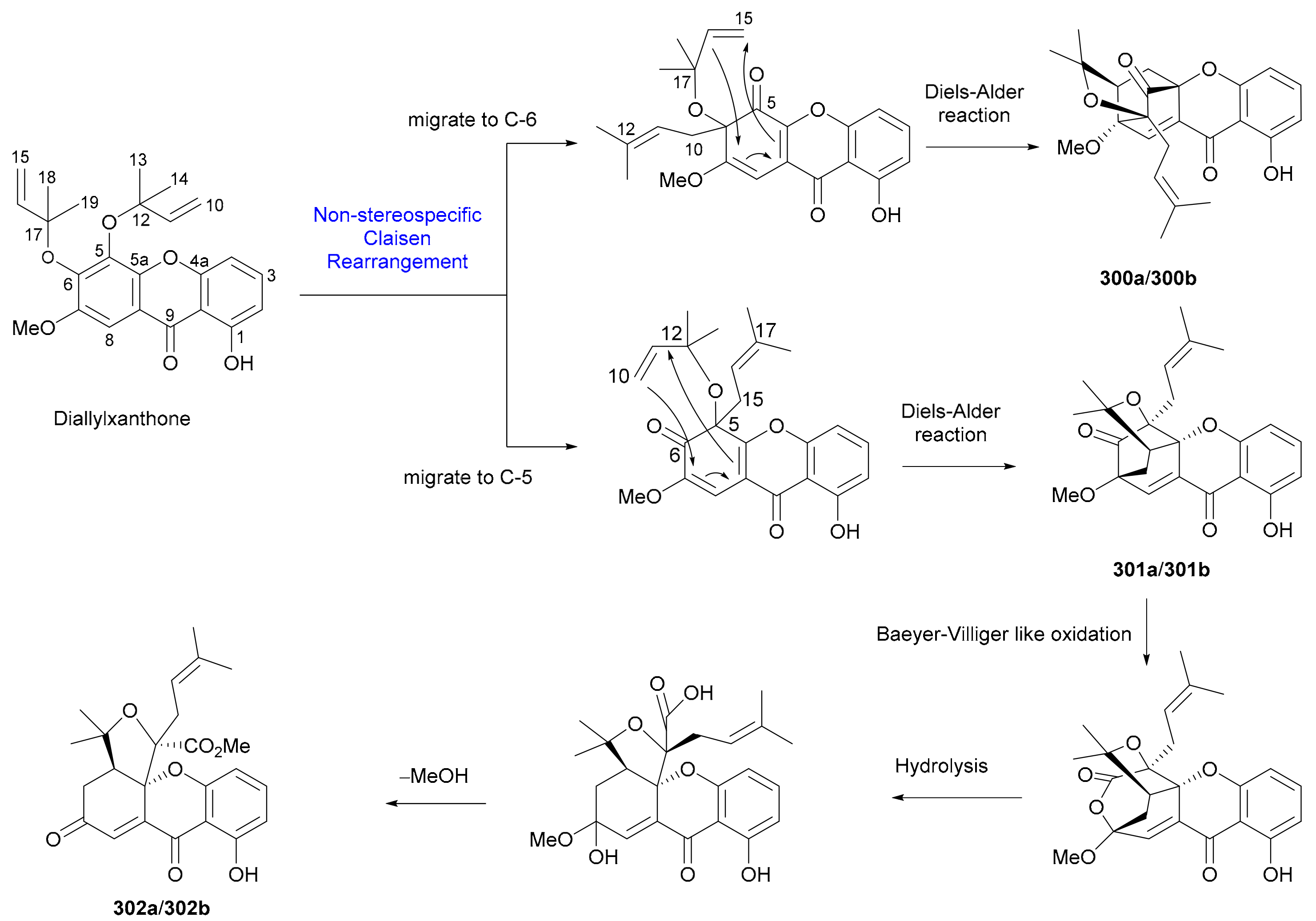{kind=link}
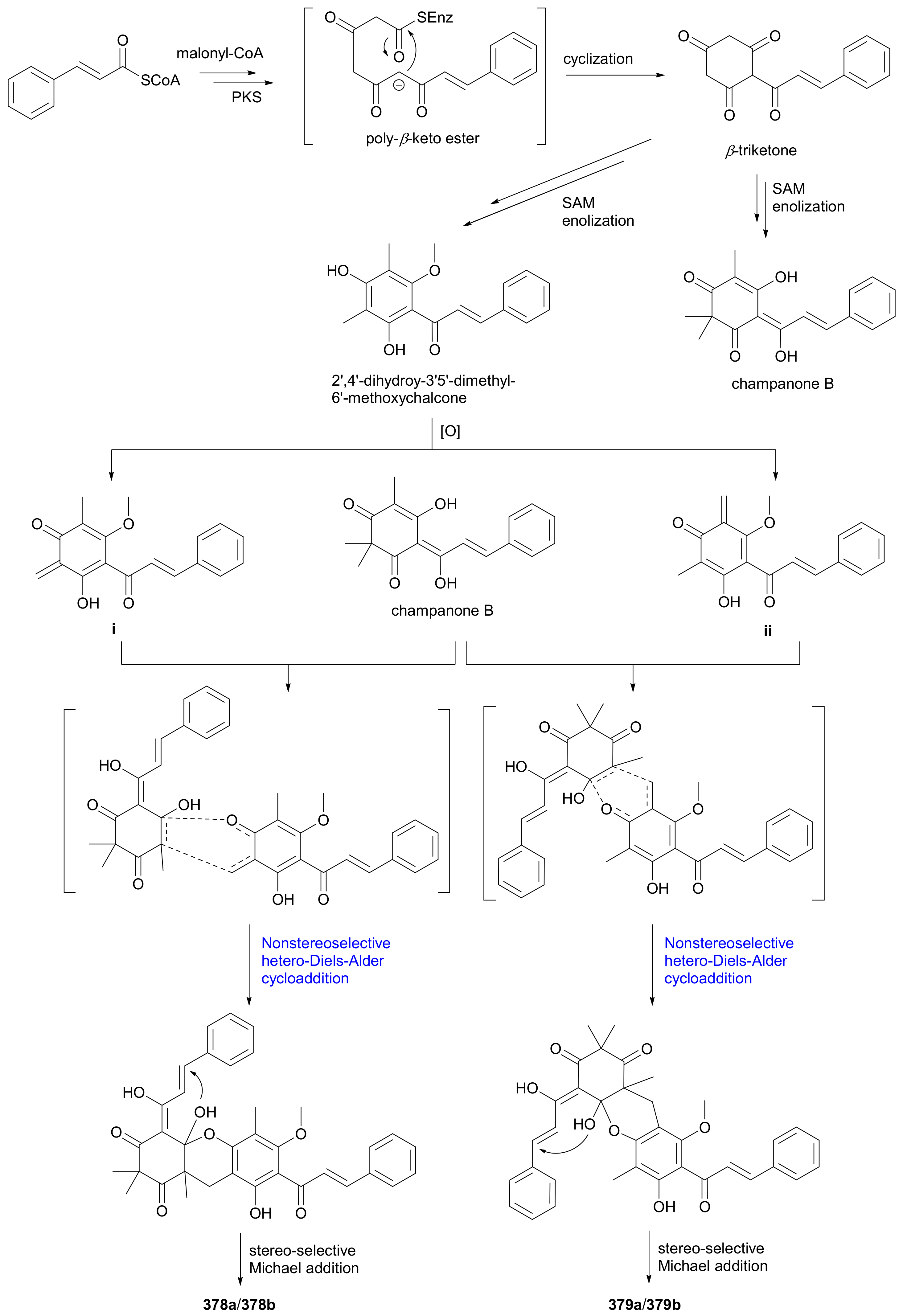{kind=link}
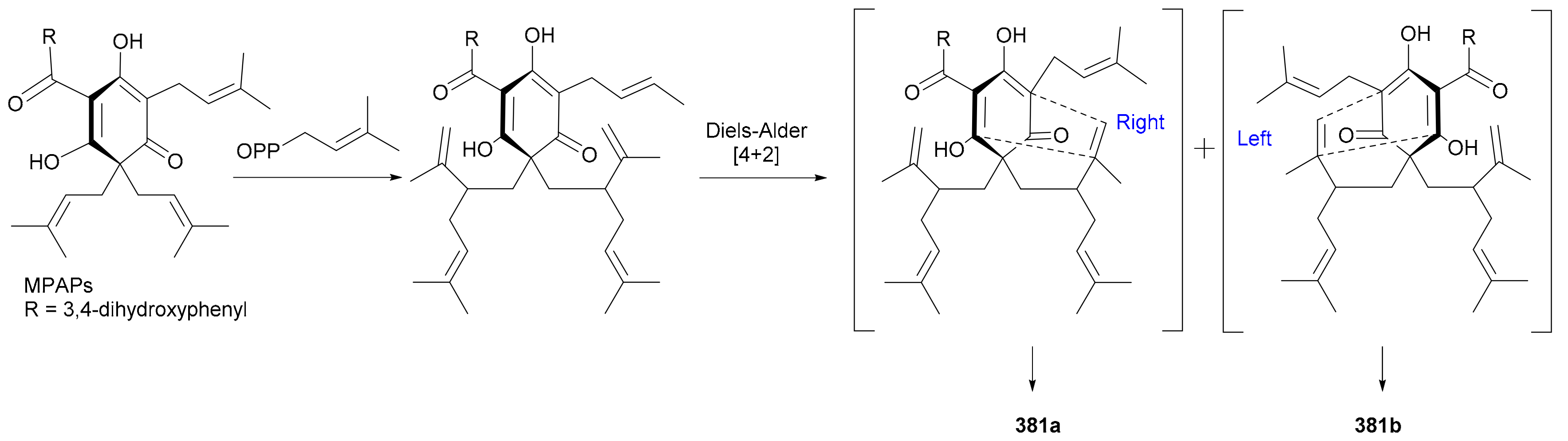{kind=link}
{kind=link}
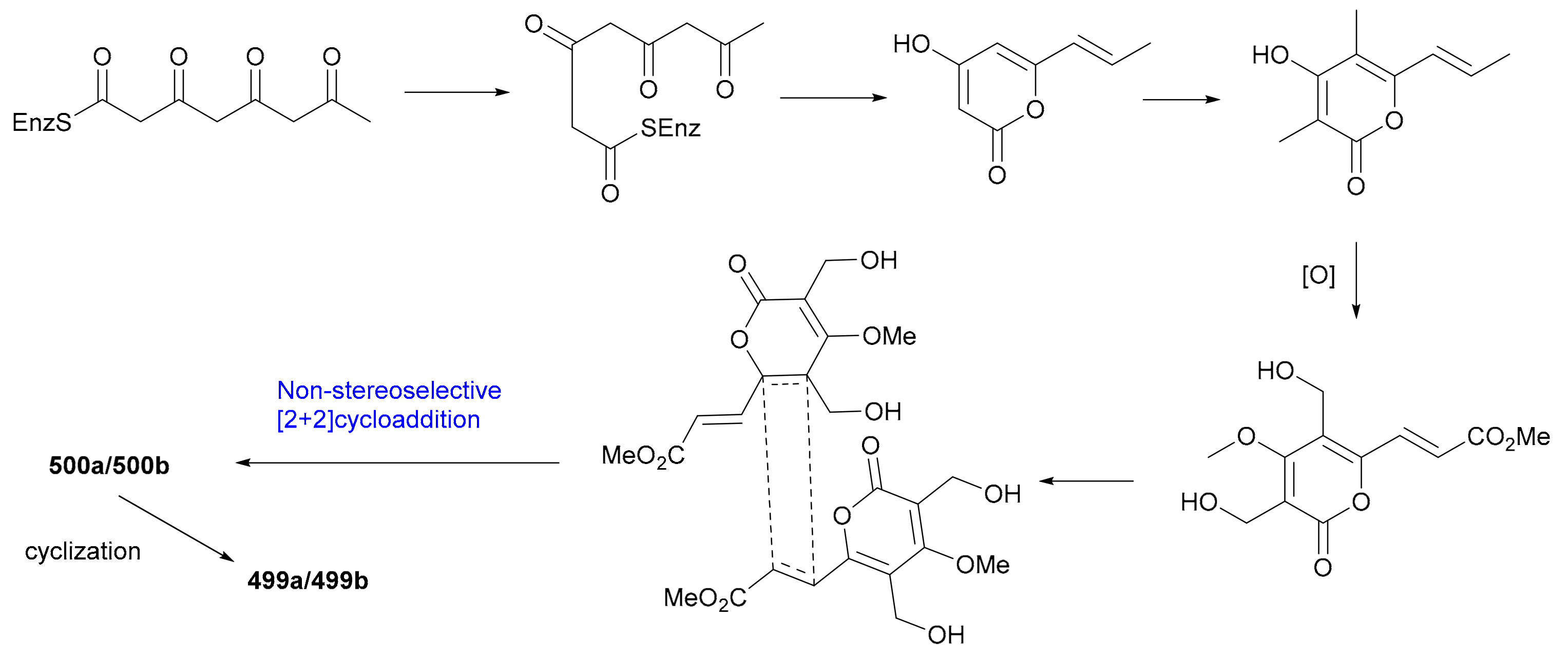{kind=link}
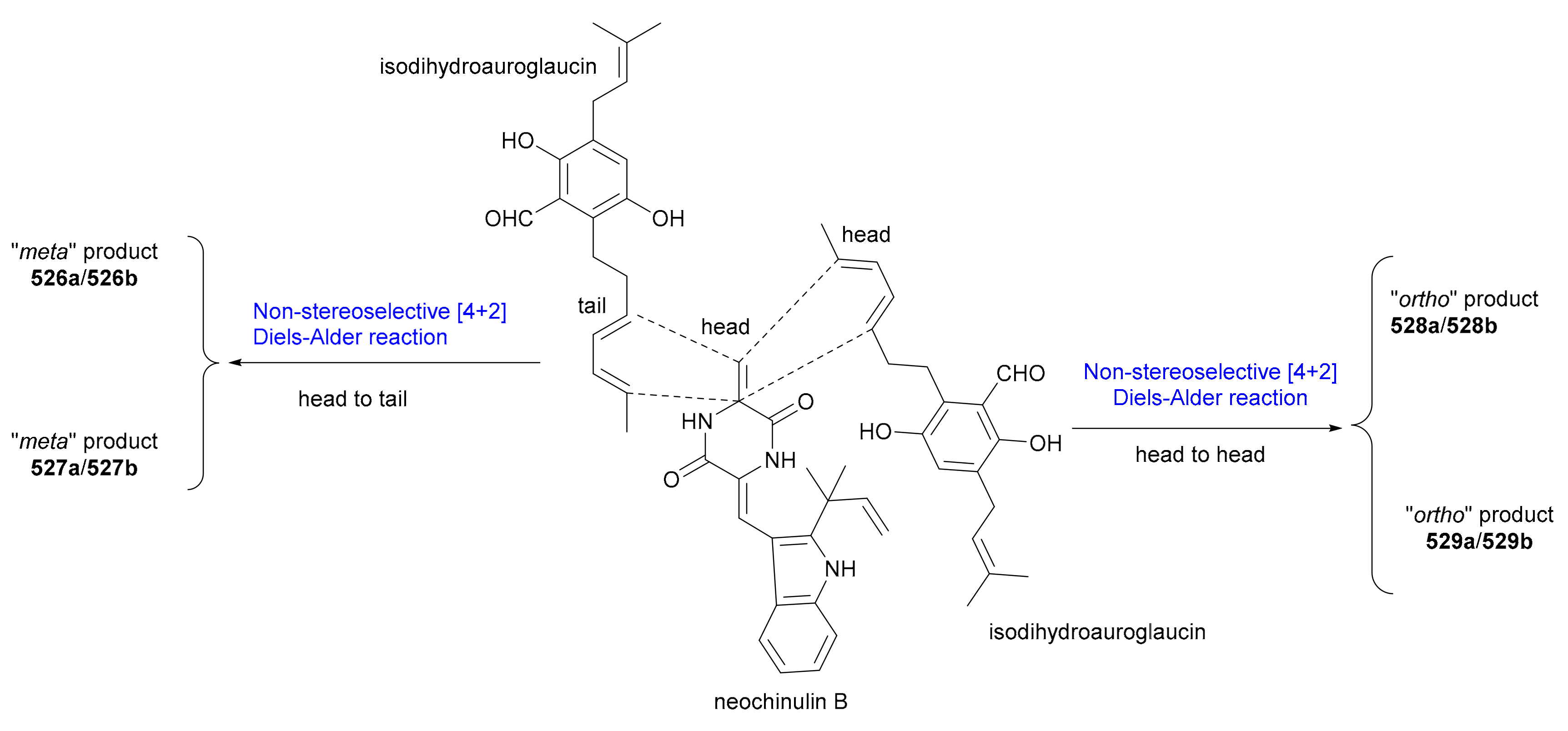{kind=link}
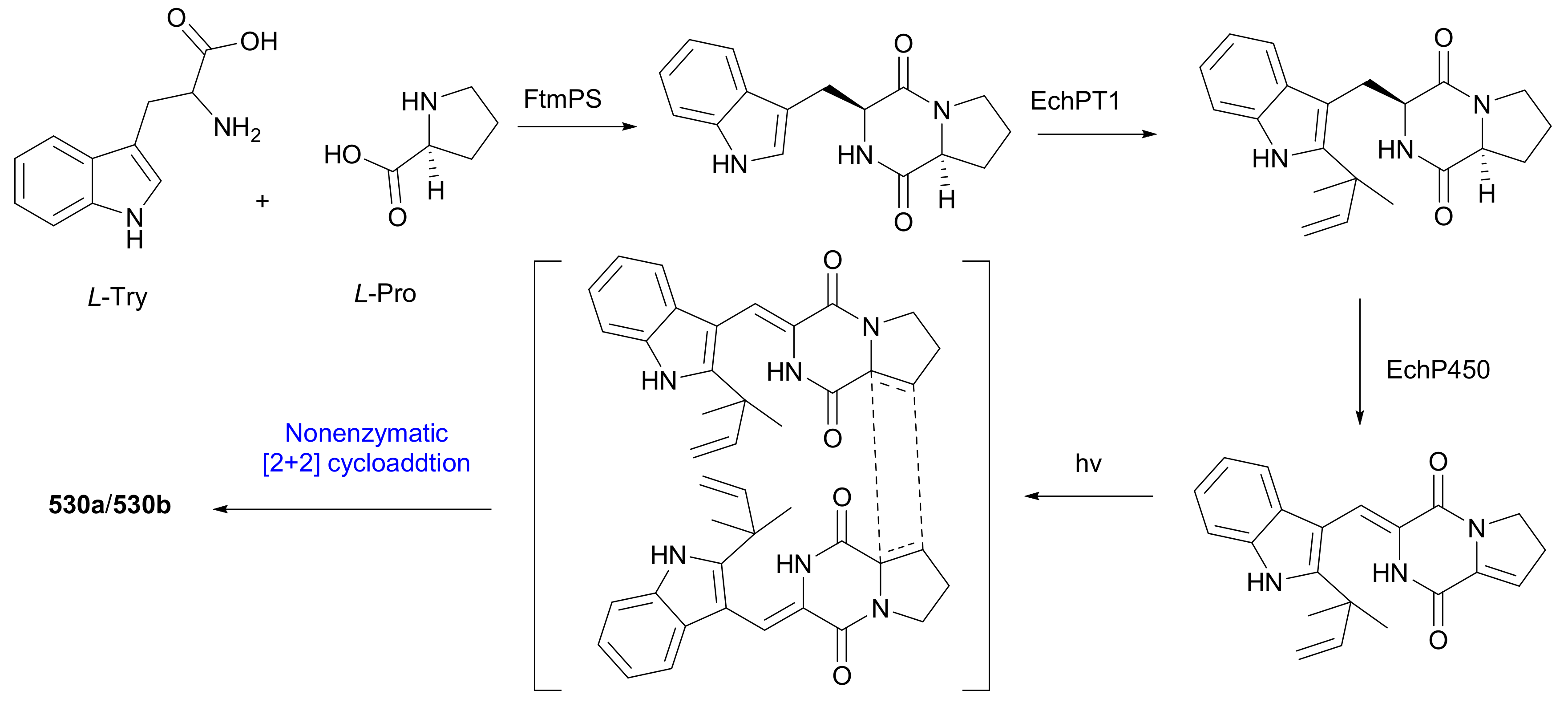{kind=link}
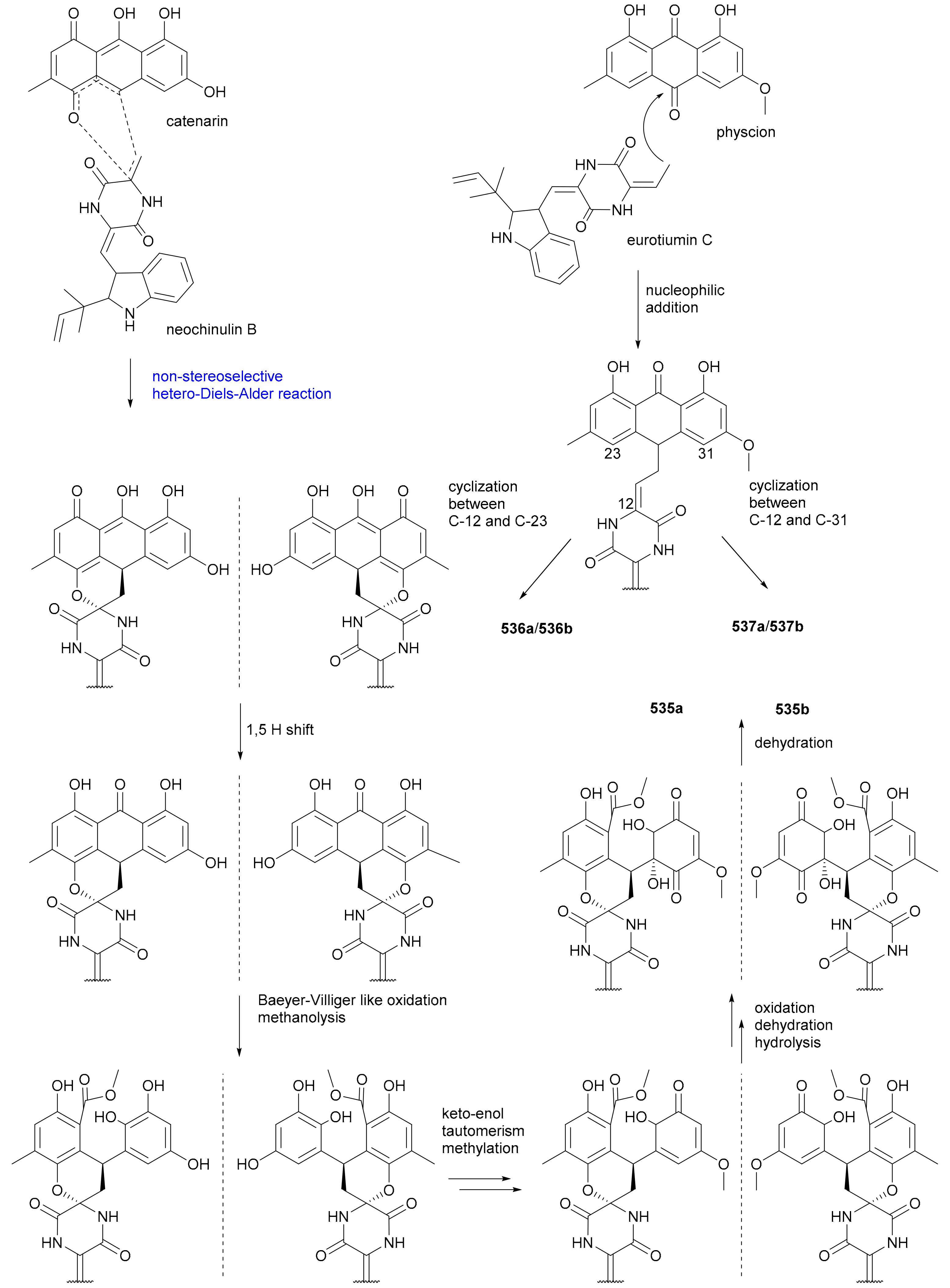{kind=link}
Table 1.
J7,8 Values, specific optical rotations and ECD data of 8,4′-oxyneolignians.
| No. | C-7 & C-8 Configurations | J7,8 Values | Specific Optical Rotations | ECD Data | ||||
|---|---|---|---|---|---|---|---|---|
| J7,8 (Hz) | Solvent | [α]D | Solvent | T (°C) | Δε | λ (nm) | ||
| 1a | (7R,8S)-erythro | 3.5 | CD3OD | +3.3 | MeOH | 25 | +2.25 | 239 |
| 1b | (7S,8R)-erythro | 3.5 | CD3OD | −4.7 | MeOH | 25 | −2.75 | 239 |
| 2a | (7R,8R)-threo | 6.8 | CD3OD | −21.7 | MeOH | 25 | −3.45 | 231 |
| 2b | (7S,8S)-threo | 6.8 | CD3OD | +16.0 | MeOH | 25 | +2.83 | 232 |
| 3a | (7S,8S)-threo | 8.8 | CDCl3 | +18.0 | MeOH | 20 | −1.12 | 239 |
| 3b | (7R,8R)-threo | 8.8 | CDCl3 | −20.0 | MeOH | 20 | +1.40 | 237 |
| 4a | (7R,8S)-erythro | 2.6 | CDCl3 | +20.5 | MeOH | 20 | +2.38 | 245 |
| 4b | (7S,8R)-erythro | 2.6 | CDCl3 | −22.0 | MeOH | 20 | −1.89 | 243 |
| 5a | (7R,8S)-erythro | 3.3 | CDCl3 | +32.0 | MeOH | 20 | +0.25 | 238 |
| 5b | (7S,8R)-erythro | 3.3 | CDCl3 | −28.2 | MeOH | 20 | −0.01 | 238 |
| 6a | (7S,8S)-threo | 6.9 | CDCl3 | +36.7 | MeOH | 20 | +8.92 | 239 |
| 6b | (7R,8R)-threo | 6.9 | CDCl3 | −33.5 | MeOH | 20 | −6.50 | 240 |
| 7a | (7R,8R)-threo | 8.0 | CDCl3 | −28.5 | MeOH | 20 | +9.64 | 230 |
| 7b | (7S,8S)-threo | 8.0 | CDCl3 | +26.9 | MeOH | 20 | −8.74 | 230 |
| 8a | (7R,8S)-erythro | 7.2 | CDCl3 | −31.0 | MeOH | 20 | −3.55 | 240 |
| 8b | (7S,8R)-erythro | 7.2 | CDCl3 | +29.0 | MeOH | 20 | +2.80 | 240 |
| 9a | (7S,8S)-threo | 7.4 | CDCl3 | +34.0 | MeOH | 20 | +3.70 | 244 |
| 9b | (7R,8R)-threo | 7.4 | CDCl3 | −34.0 | MeOH | 20 | −2.81 | 243 |
| 10a | (7R,8R)-threo | 6.5 | CDCl3 | −28.0 | MeOH | 20 | +16.90 | 240 |
| 10b | (7S,8S)-threo | 6.5 | CDCl3 | +32.0 | MeOH | 20 | −17.65 | 238 |
| 11a | (7R,8R)-threo | 7.6 | CDCl3 | −20.0 | MeOH | 20 | −6.57 | 238 |
| 11b | (7S,8S)-threo | 7.6 | CDCl3 | +21.0 | MeOH | 20 | +8.87 | 242 |
| 12a | (7R,8S)-erythro | 4.7 | CDCl3 | −31.0 | MeOH | 20 | −3.55 | 240 |
| 12b | (7S,8R)-erythro | 4.7 | CDCl3 | +29.0 | MeOH | 20 | +2.80 | 240 |
| 13a | (7S,8R)-erythro | 3.0 | CDCl3 | +17.1 | CHCl3 | 20 | −1.03 | 232 |
| 13b | (7R,8S)-erythro | 3.0 | CDCl3 | −16.2 | CHCl3 | 20 | +1.03 | 232 |
| 14a | (7S,8S)-threo | 6.3 | CDCl3 | −35.1 | CHCl3 | 20 | −1.14 | 240 |
| 14b | (7R,8R)-threo | 6.3 | CDCl3 | +32.6 | CHCl3 | 20 | +1.17 | 240 |
| 15a | (7R,8R)-threo | 8.1 | CDCl3 | +30.4 | CHCl3 | 20 | +1.58 | 240 |
| 15b | (7S,8S)-threo | 8.1 | CDCl3 | −29.8 | CHCl3 | 20 | −1.54 | 240 |
| 16a | (7S,8R)-erythro | 3.3 | CDCl3 | +16.6 | CHCl3 | 20 | +3.18 | 234 |
| 16b | (7R,8S)-erythro | 3.3 | CDCl3 | −16.4 | CHCl3 | 20 | −3.20 | 234 |
| 17a | (7R,8R)-threo | 8.0 | CDCl3 | +7.0 | MeOH | 20 | - | - |
| 17b | (7S,8S)-threo | 8.0 | CDCl3 | −7.0 | MeOH | 20 | - | - |
| 18a | (7S,8R)-erythro | 4.7 | CDCl3 | +20.0 | MeOH | 20 | +11.17 | 230 |
| 18b | (7R,8S)-erythro | 4.7 | CDCl3 | −18.0 | MeOH | 20 | −4.00 | 235 |
| 19a | (7S,8S)-threo | 6.1 | CD3OD | +18.0 | MeOH | 20 | +1.70 | 230 |
| 19b | (7R,8R)-threo | 6.1 | CD3OD | −18.0 | MeOH | 20 | −1.20 | 230 |
| 20a | (7R,8S)-erythro | 4.8 | CD3OD | −10.0 | MeOH | 20 | +2.70 | 232 |
| 20b | (7S,8R)-erythro | 4.8 | CD3OD | +8.0 | MeOH | 20 | −2.82 | 233 |
| 21a | (7S,8R)-erythro | 5.4 | CD3OD | +15.0 | MeOH | 20 | +3.15 | 228 |
| 21b | (7R,8S)-erythro | 5.4 | CD3OD | −15.0 | MeOH | 20 | −2.65 | 228 |
| 22a | (7S,8S)-threo | 7.7 | CDCl3 | +17.2 | MeOH | 20 | −0.59 | 233 |
| 22b | (7R,8R)-threo | 7.7 | CDCl3 | −19.0 | MeOH | 20 | +0.51 | 232 |
| 23a | (7R,8R)-threo | 7.6 | CDCl3 | −24.5 | MeOH | 20 | - | - |
| 23b | (7S,8S)-threo | 7.6 | CDCl3 | +26.0 | MeOH | 20 | - | - |
| 24a | (7R,8S)-erythro | 4.6 | CDCl3 | −18.0 | MeOH | 20 | +12.20 | 241 |
| 24b | (7S,8R)-erythro | 4.6 | CDCl3 | +22.0 | MeOH | 20 | −10.81 | 241 |
| 25a | (7S,8S)-threo | 6.8 | CDCl3 | +23.0 | MeOH | 20 | +0.57 | 240 |
| 25b | (7R,8R)-threo | 6.8 | CDCl3 | −25.0 | MeOH | 20 | −1.05 | 237 |
Table 2.
Cytotoxic activities of enantiomers. (a) part 1; (b) part 2.
| Cell lines | Compds. | IC50 (μM) | Reference | Cell Lines | Compds. | IC50 (μM) | Reference |
|---|---|---|---|---|---|---|---|
| (a) | |||||||
| HL-60 | (+)-171a | 5.62 | [105] | HCT-116 | (−)-314a | 1.38 | [162] |
| (−)-171b | 3.51 | [105] | (+)-314b | >10 | [162] | ||
| (±)-171 | 2.65 | [105] | HepG2 | (−)-314a | 3.30 | [162] | |
| (+)-172a | 9.64 | [105] | (+)-314b | >10 | [162] | ||
| (−)-172b | 8.16 | [105] | BGC-823 | (−)-314a | 6.51 | [162] | |
| (±)-172 | 5.58 | [105] | (+)-314b | >10 | [162] | ||
| Hep3B | (−)-268a | >10 | [146] | NIC-H1650 | (−)-314a | 8.19 | [162] |
| (+)-268b | 5.1 | [146] | (+)-314b | >10 | [162] | ||
| HL-60 | (+)-297a | 12.08 | [156] | A2780 | (−)-314a | 2.14 | [162] |
| (−)-297b | 19.24 | [156] | (+)-314b | >10 | [162] | ||
| MDA-MB-231 | (+)-297a | >50 | [156] | SF-268 | (+)-536a | 12.5 | [261] |
| (−)-297b | 18.46 | [156] | (−)-536b | >100 | [261] | ||
| HCT-116 | (+)-312a | inactive | [160] | (+)-537a | 30.1 | [261] | |
| (−)-312b | 14.23 | [160] | (−)-537b | >100 | [261] | ||
| A549 | (−)-390a | 4.64 | [8] | HepG2 | (+)-536a | 15.0 | [261] |
| (+)-390b | 10.54 | [8] | (−)-536b | >100 | [261] | ||
| MCF-7 | (−)-390a | 5.60 | [8] | (+)-537a | 37.3 | [261] | |
| (+)-390b | 15.52 | [8] | (−)-537b | >100 | [261] | ||
| MDA-MB-231 | (−)-390a | 3.86 | [8] | ||||
| (+)-390b | 11.86 | [8] | |||||
| (b) | |||||||
| HL-60 | (+)-381a | 3.42 | [191,192] | MCF-7 | (+)-381a | 4.18 | [191,192] |
| (−)-381b | >20 | [191,192] | (−)-381b | >20 | [191,192] | ||
| (−)-382a | 16.54 | [191,192] | (−)-382a | 14.44 | [191,192] | ||
| (+)-382b | >40 | [191,192] | (+)-382b | >40 | [191,192] | ||
| (±)-382 | 14.44 | [191,192] | (±)-382 | 36.00 | [191,192] | ||
| (−)-383a | 3.15 | [191,192] | (−)-383a | 5.85 | [191,192] | ||
| (+)-383b | 2.35 | [191,192] | (+)-383b | 10.76 | [191,192] | ||
| (±)-383 | 18.08 | [191,192] | (±)-383 | 17.05 | [191,192] | ||
| (−)-384a | 3.45 | [191,192] | (−)-384a | 3.17 | [191,192] | ||
| (+)-384b | 2.36 | [191,192] | (+)-384b | 3.08 | [191,192] | ||
| (±)-384 | 2.93 | [191,192] | (±)-384 | 11.92 | [191,192] | ||
| (−)-385a | 2.63 | [191,192] | (−)-385a | 14.60 | [191,192] | ||
| (+)-385b | 5.41 | [191,192] | (+)-385b | 15.02 | [191,192] | ||
| (±)-385 | 13.90 | [191,192] | (±)-385 | 15.47 | [191,192] | ||
| SMMC-7721 | (+)-381a | 4.19 | [191,192] | SW480 | (+)-381a | 7.22 | [191,192] |
| (−)-381b | >20 | [191,192] | (−)-381b | >20 | [191,192] | ||
| (−)-382a | 16.20 | [191,192] | (−)-382a | 17.43 | [191,192] | ||
| (+)-382b | >40 | [191,192] | (+)-382b | >40 | [191,192] | ||
| (±)-382 | 22.83 | [191,192] | (±)-382 | 27.73 | [191,192] | ||
| (−)-383a | 5.55 | [191,192] | (−)-383a | 3.04 | [191,192] | ||
| (+)-383b | 12.30 | [191,192] | (+)-383b | 5.53 | [191,192] | ||
| (±)-383 | 20.71 | [191,192] | (±)-383 | 13.60 | [191,192] | ||
| (−)-384a | 3.80 | [191,192] | (−)-384a | 1.99 | [191,192] | ||
| (+)-384b | 10.83 | [191,192] | (+)-384b | 1.52 | [191,192] | ||
| (±)-384 | 15.36 | [191,192] | (±)-384 | 7.62 | [191,192] | ||
| (−)-385a | 12.53 | [191,192] | (−)-385a | 3.39 | [191,192] | ||
| (+)-385b | 13.21 | [191,192] | (+)-385b | 4.31 | [191,192] | ||
| (±)-385 | 14.82 | [191,192] | (±)-385 | 8.84 | [191,192] | ||
| A549 | (+)-381a | 4.51 | [191,192] | A549 | (±)-383 | 24.02 | [191,192] |
| (−)-381b | >20 | [191,192] | (−)-384a | 2.96 | [191,192] | ||
| (−)-382a | 17.27 | [191,192] | (+)-384b | 18.78 | [191,192] | ||
| (+)-382b | >40 | [191,192] | (±)-384 | 15.44 | [191,192] | ||
| (±)-382 | 17.30 | [191,192] | (−)-385a | 22.36 | [191,192] | ||
| (−)-383a | 4.40 | [191,192] | (+)-385b | 9.53 | [191,192] | ||
| (+)-383b | 16.33 | [191,192] | (±)-385 | 10.24 | [191,192] | ||
Table 3.
Antiviral activities of enantiomers a.
| Virus/Host | Compds. | EC50 (μM) | IC50 (μM) | SI | Reference |
|---|---|---|---|---|---|
| HSV-1/Vero | (+)-254a | 6.41 | 19.25 | 3.0 | [7] |
| (−)-254b | 3.70 | 16.02 | 4.3 | [7] | |
| (±)-254 | 1.23 | 2.14 | 1.7 | [7] | |
| Acyclovir b | >100 | 0.41 | >243.9 | [7] | |
| Virus A (H3N3)/MDCK | (+)-104a | 9.86 | 19.25 | 1.9 | [67] |
| (−)-104b | 11.11 | 77.61 | 6.9 | [67] | |
| (±)-104 | 3.13 | 6.42 | 2.1 | [67] | |
| (−)-105a | 8.62 | 77.61 | 7.9 | [67] | |
| (+)-105b | 17.46 | 57.74 | 3.3 | [67] | |
| (±)-105 | 2.87 | 25.87 | 9.0 | [67] | |
| Oseltamivir b | 3.38 | 3073 | 910.5 | [67] | |
| Ribavirin b | 6.19 | 4771 | 770.7 | [67] | |
| KSHV/Vero | (+)-368a | 8.75 | 140.6 | 16.06 | [184] |
| (−)-368b | 29.13 | 173.7 | 5.96 | [184] | |
| (+)-369a | 202.9 | >500 | >2.46 | [184] | |
| (−)-369b | 140.9 17.67 | >500 | >2.55 | [184] | |
| (+)-370a | 211.1 | 12.51 | [184] | ||
| (−)-370b | 39.80 | >300 | >7.50 | [184] | |
| (+)-371a | 40.00 | >300 | >7.50 | [184] | |
| (−)-371b | 158.50 | >300 | >1.89 | [184] | |
| Acyclovir b | 0.41 | 99.18 | 241.9 | [184] | |
| EV71/Vero | (−)-558a | 69.1 | 143.7 | 2.1 | [272] |
| (+)-558b | 14.2 | 130.2 | 9.2 | [272] | |
| (±)-558 | 14.2 | 126.6 | 7.9 | [272] | |
| Ribavirin b | >256.1 | 4098 | >16 | [272] |
a EC50 represents concentration required to inhibit virus growth by 50%; IC50 represents concentration required to inhibit host cell growth by 50%; SI (Selectivity index) = IC50/EC50. b Positive controls.
Table 4.
Antibacterial activities of enantiomers.
| Bacterial Strains | Compds. | Activities | Reference |
|---|---|---|---|
| Enterococcus faecalis | (+)-147a | MIC = 21.97 μg/mL | [97] |
| (−)-147b | MIC = 12.54 μg/mL | [97] | |
| Penicilin a | MIC < 2.96 μg/mL | [97] | |
| Staphylococcus aureus | (+)-285a | IC50 = 1.27 μM | [153] |
| (−)-285b | IC50 = 1.79 μM | [153] | |
| (+)-286a | IC50 = 2.27 μM | [153] | |
| (−)-286b | IC50 = 4.3 μM | [153] | |
| (+)-287a | IC50 = 3.6 μM | [153] | |
| (−)-287b | IC50 = 6.0 μM | [153] | |
| (+)-288a | IC50 = 0.61 μM | [153] | |
| (−)-288b | IC50 = 2.27 μM | [153] | |
| Chloramphenicol a | IC50 = 0.43 μM | [153] |
a Positive controls.
Table 5.
Antifungal activities of enantiomers.
| Fungal Strains | Compds. | Activities | Reference |
|---|---|---|---|
| Candida albicans | (−)-464a | MIC = 26.2 μM | [219] |
| (+)-464b | inactive | [219] | |
| Candida albicans | (+)-483a | MIC80 = 19.5 μg/mL | [229] |
| (−)-483b | MIC80 = 48.8 μg/mL | [229] | |
| (+)-484a | MIC80 = 24.0 μg/mL | [229] | |
| (−)-484b | MIC80 > 50.0 μg/mL | [229] | |
| Fusarium solani | (+)-524a | MIC > 64 μg/mL | [256] |
| (−)-524b | MIC = 32 μg/mL | [256] |
Table 6.
Anti-inflammatory activities of enantiomers.
| Assay Model | Compds. | IC50 (μM) | Reference |
|---|---|---|---|
| BV-2/NO | (+)-43a | 26.4 | [39] |
| (−)-43b | 8.9 | [39] | |
| (+)-44a | 5.9 | [39] | |
| (−)-44b | 14.7 | [39] | |
| Quercetin a | 17.0 | [39] | |
| RAW 264.7/NO | (+)-82a | 17.9 | [54] |
| (−)-82b | 5.6 | [54] | |
| (+)-83a | 15.1 | [54] | |
| (−)-83b | 4.3 | [54] | |
| RAW 264.7/NO | (+)-221a | 3.6 | [124] |
| (−)-221b | 22.8 | [124] | |
| (±)-221 | 14.7 | [124] | |
| Indomethacin a | 42.2 | [124] | |
| BV-2/NO | (+)-407a | 6.9 | [130] |
| (−)-407b | 1.4 | [130] | |
| (±)-407 | 1.0 | [130] | |
| Minocyclinebe a | 27.2 | [130] |
a positive controls.
Table 7.
Antioxidative activities of enantiomers.
| Assay Model | Compds. | IC50 (μM) | Reference |
|---|---|---|---|
| DPPH | (+)-527a | 5.8 | [257] |
| (−)-527b | 23.5 | [257] | |
| (+)-528a | 9.8 | [257] | |
| (−)-528b | 24.9 | [257] | |
| (+)-529a | 3.7 | [257] | |
| (−)-529b | 6.1 | [257] | |
| Ascorbic acid a | 23.0 | [257] |
a Positive control.
Table 8.
Cell protective activities of enantiomers.
| Cell/Inducing Agents | Compds. | Cell Viability | Reference |
|---|---|---|---|
| SH-SY5Y/H2O2 | (−)-24a | 54.7% at 50 μM | [21] |
| (+)-24b | 70.5% at 50 μM | [21] | |
| Trolox a | ~69.0% at 50 μM | [21] | |
| SH-SY5Y/H2O2 | (+)-72a | ~69% at 25 μM | [46] |
| (−)-72b | inactive at 25 μM | [46] | |
| Trolox a | ~62% at 25 μM | [46] | |
| SH-SY5Y/H2O2 | (+)-119a | 76.29% at 50 μM | [75] |
| (−)-119b | 56.48% at 50 μM | [75] | |
| PC12/OKA | (+)-129a | 65.4% at 10 μM | [83] |
| (−)-129b | 83.4% at 10 μM | [83] | |
| (+)-130a | 91.2% at 10 μM | [83] | |
| (−)-130b | 69.5% at 10 μM | [83] | |
| H9C2/ischemia-hypoxia | (−)-165a | ~51% at 0.1 μM | [103] |
| (+)-165b | inactive | [103] | |
| (−)-166a | ~55% at 0.1 μM | [103] | |
| (+)-166b | ~45% at 0.1 μM | [103] | |
| (−)-173a | inactive | [103] | |
| (+)-173b | ~52% at 0.1 μM | [103] | |
| Salvianolic acid B a | ~46% at 0.1 μM | [103] | |
| HUVEC/glucose | (+)-423a | 102.6% at 1 μM | [200] |
| (−)-423b | 79.9% at 1 μM | [200] | |
| (+)-426a | 102.6% at 1 μM | [200] | |
| (−)-426b | 79.9% at 1 μM | [200] | |
| Cortical neurons/MPP+ | (+)-433a | ~90% at 16 μM | [206] |
| (−)-433b | inactive | [206] |
a Positive controls.
Table 9.
Enzyme inhibitory activities of enantiomers.
| Enzymes | Compds. | IC50 (μM) | Reference |
|---|---|---|---|
| AchE | (+)-157a | >100 | [100] |
| (−)-157b | 28.3 | [100] | |
| Galanthamine a | 1.9 | [100] | |
| AchE | (+)-523a | 2.3 | [256] |
| (−)-523b | 13.8 | [256] | |
| (±)-523 | 9.5 | [256] | |
| Tacrine a | 0.14 | [256] | |
| α-Glucosidase | (+)-495a | 63.7 | [237] |
| (−)-495b | 27.9 | [237] | |
| (±)-495 | 36.1 | [237] | |
| Acarbose a | 477.0 | [237] | |
| PTP1B | (−)-339a | Inactive | [176] |
| (+)-339b | 43.6 | [176] | |
| (+)-340a | 38.1 | [176] | |
| (−)-340b | Inactive | [176] | |
| (−)-341a | 61.0 | [176] | |
| (+)-341b | Inactive | [176] | |
| (−)-342a | 58.2 | [176] | |
| (+)-342b | Inactive | [176] | |
| Oleanolic acid a | 2.5 | [176] | |
| COX-2 | (+)-641a | 2.52 | [305] |
| (−)-641b | 6.04 | [305] | |
| (+)-644a | 17.8 | [305] | |
| (−)-644b | 9.7 | [305] | |
| Celecoxib a | 0.016 | [305] |
a Positive controls.
Table 10.
Aβ aggregation inhibition of enantiomers.
| Compds. | Inhibition (%) | Reference |
|---|---|---|
| (+)-6a | 31.2 | [19] |
| (−)-6b | 75.3 | [19] |
| Curcumin a | 62.1 | [19] |
| (+)-73a | 62.1 | [51] |
| (−)-73b | 81.6 | [51] |
| Curcumin a | 63.2 | [51] |
| (+)-132a | 85.8 | [85] |
| (−)-132b | 73.6 | [85] |
| Curcumin a | 57.0 | [85] |
| (+)-242a | 33.9 | [131] |
| (−)-242b | 50.6 | [131] |
| Curcumin a | 63.3 | [131] |
a Positive controls.
Table 11.
Miscellaneous activities of enantiomers.
| Models | Compds. | Activities | Reference |
|---|---|---|---|
| MT1 receptor agonistic activity | (−)-145a | agonistic rate = 11.26% | [93] |
| (+)-145b | inactive | [93] | |
| MT2 receptor agonistic activity | (−)-145a | agonistic rate = 52.44% | [93] |
| (+)-145b | inactive | [93] | |
| NF-κB inhibition | (+)-330a | inactive | [169] |
| (−)-330b | IC50 = 7.27 μM | [169] | |
| Antifouling activity | (−)-530a | inactive | [259] |
| (+)-530b | adhesive rate = 48.4% | [259] | |
| Protein-protein interaction inhibition | (+)-647a | inactive | [308] |
| (−)-647b | modestly | [308] | |
| Olfactory responses | (+)-648a | strong | [309] |
| (−)-648b | weak | [309] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Yu, J.-H.; Yu, Z.-P.; Capon, R.J.; Zhang, H. Natural Enantiomers: Occurrence, Biogenesis and Biological Properties. Molecules 2022, 27, 1279. https://doi.org/10.3390/molecules27041279
AMA Style
Yu J-H, Yu Z-P, Capon RJ, Zhang H. Natural Enantiomers: Occurrence, Biogenesis and Biological Properties. Molecules. 2022; 27(4):1279. https://doi.org/10.3390/molecules27041279
Chicago/Turabian StyleYu, Jin-Hai, Zhi-Pu Yu, Robert J. Capon, and Hua Zhang. 2022. "Natural Enantiomers: Occurrence, Biogenesis and Biological Properties" Molecules 27, no. 4: 1279. https://doi.org/10.3390/molecules27041279