Fertimetro, a Principle and Device to Measure Soil Nutrient Availability for Plants by Microbial Degradation Rates on Differently-Spiked Buried Threads

 ,
,  ,
, 

Abstract
:1. Introduction
2. Materials and Methods
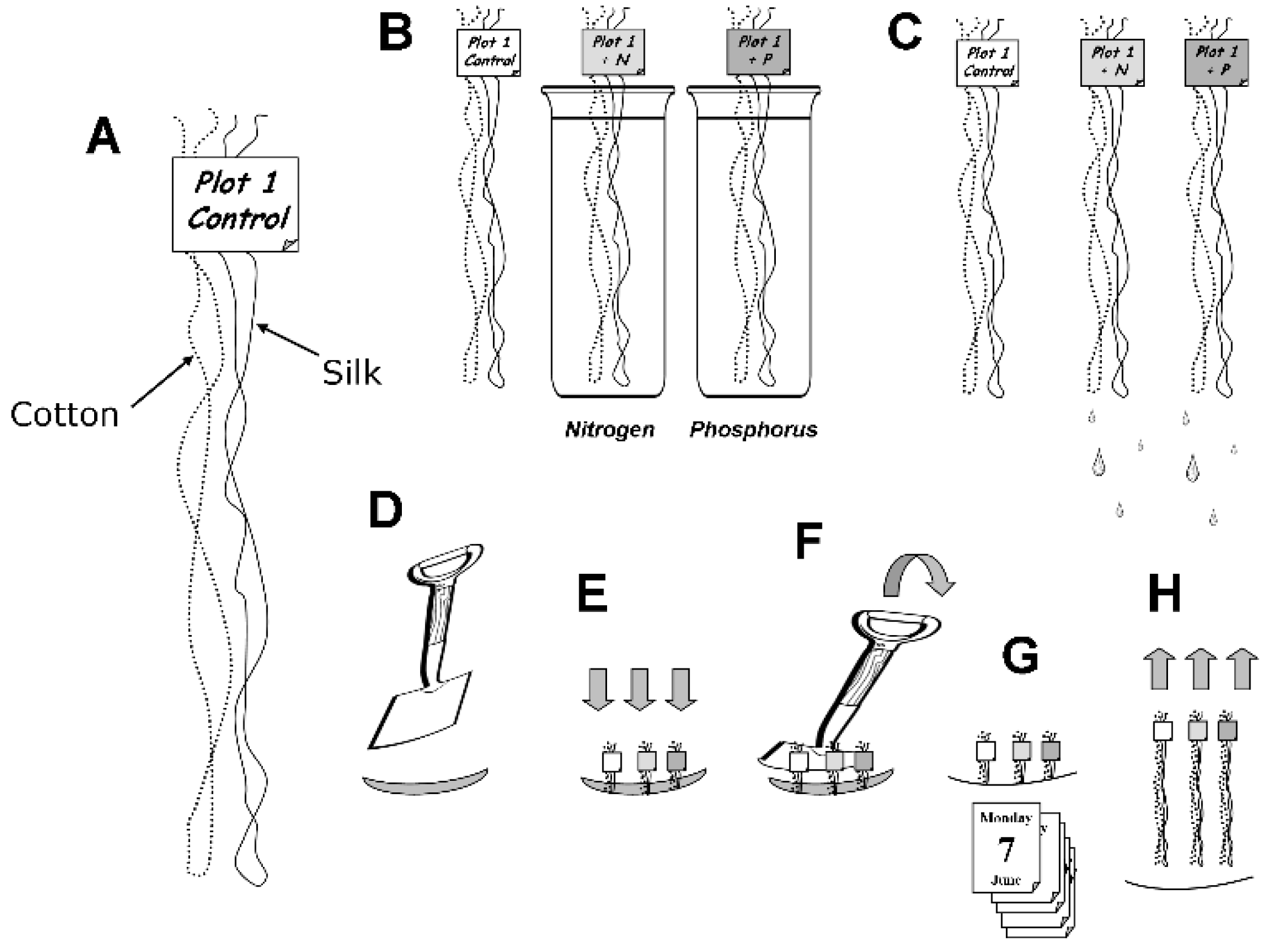
2.1. The Experimental Site
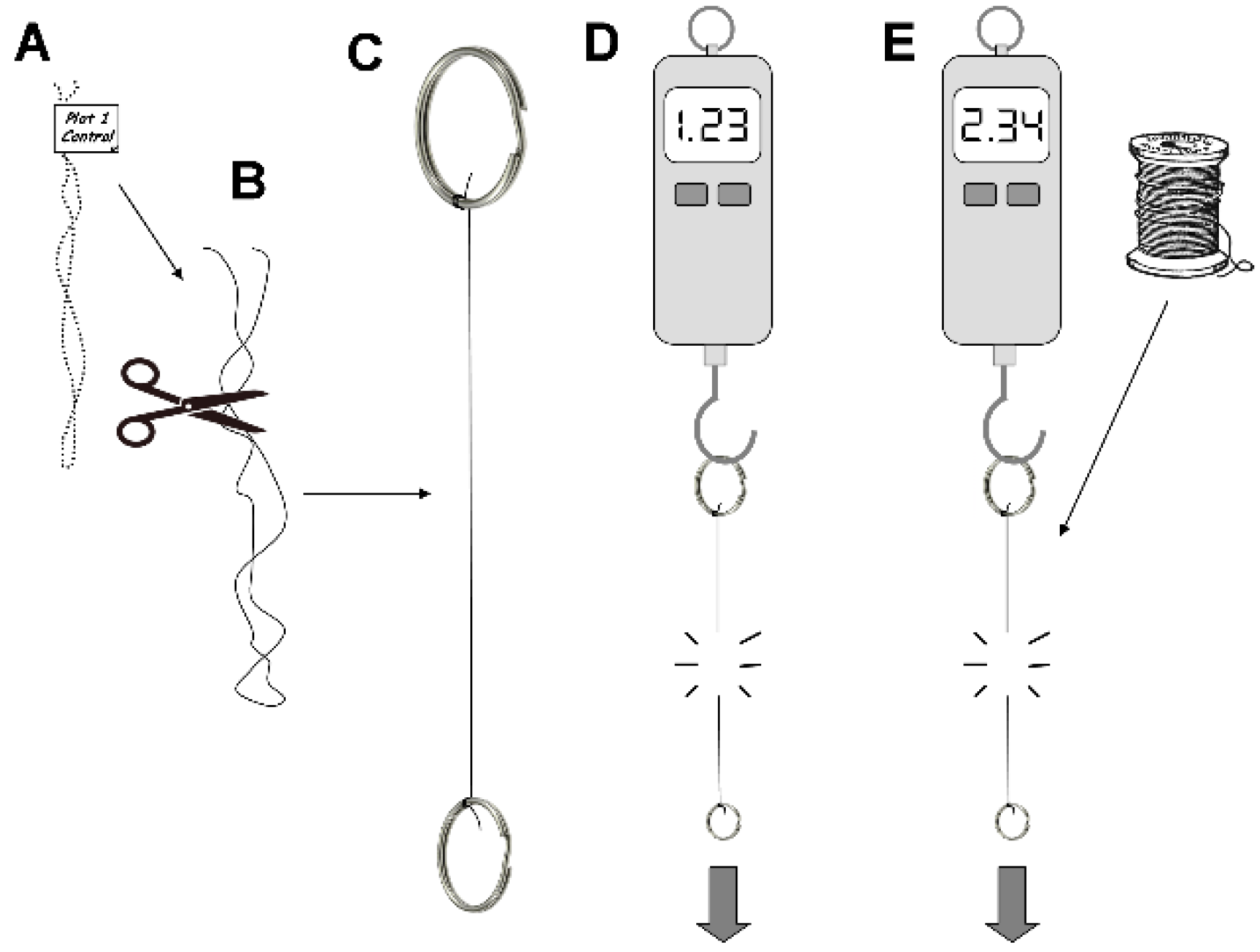
2.2. Analysis of Soil Microbial Activities on Threads
3. Results and Discussion
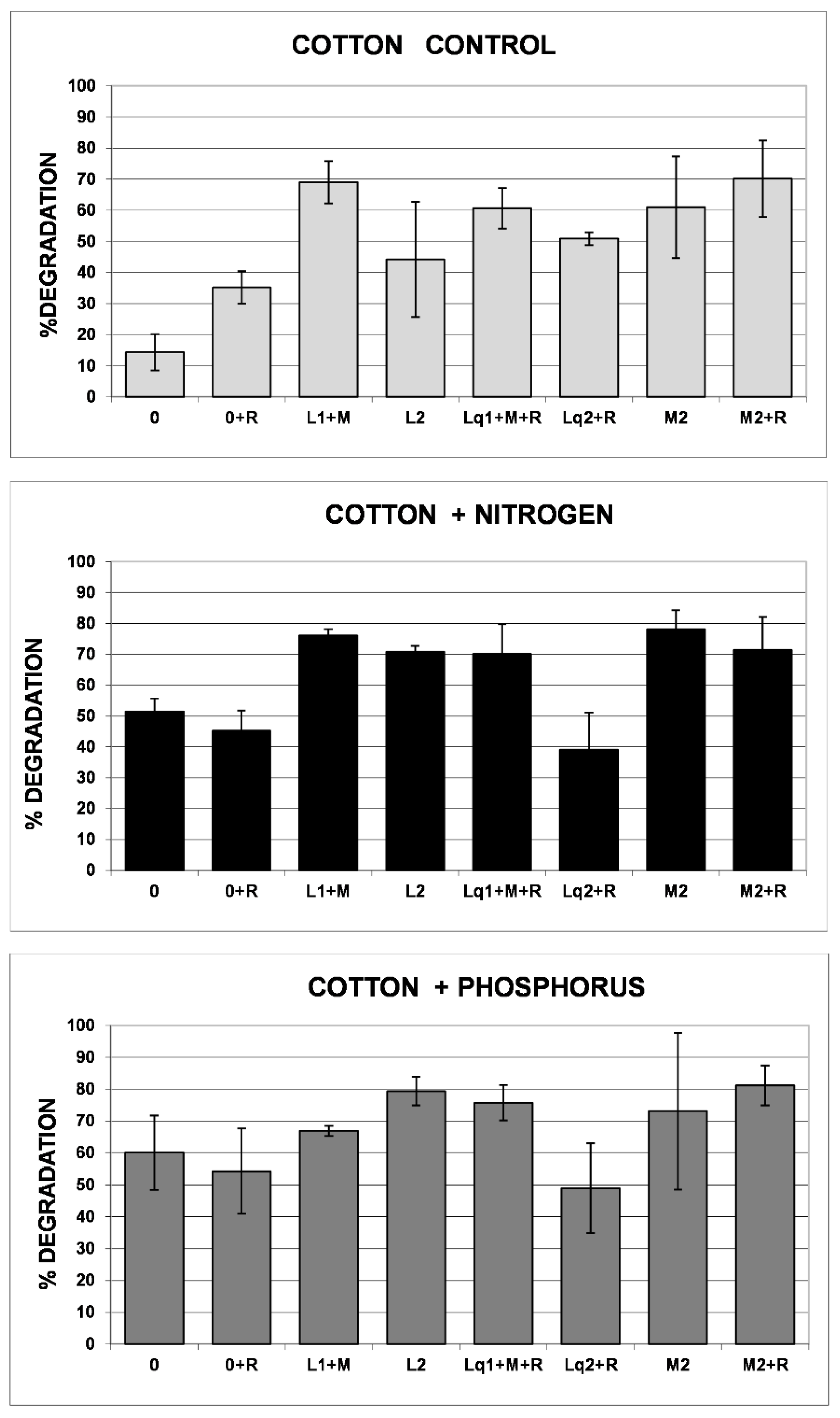
3.1. Cotton Threads’ Reporting Performance
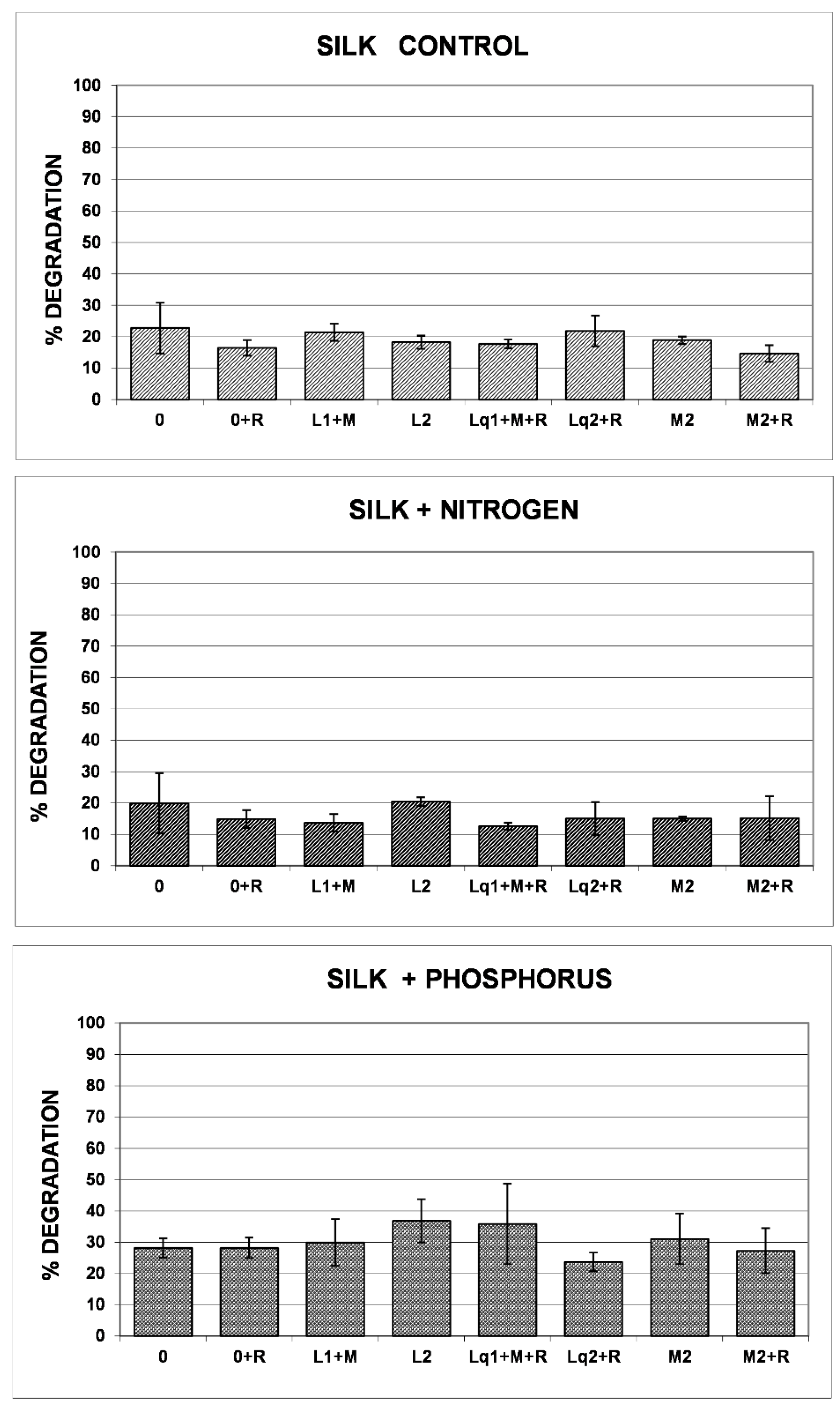
3.2. Silk Threads’ Reporting Performance
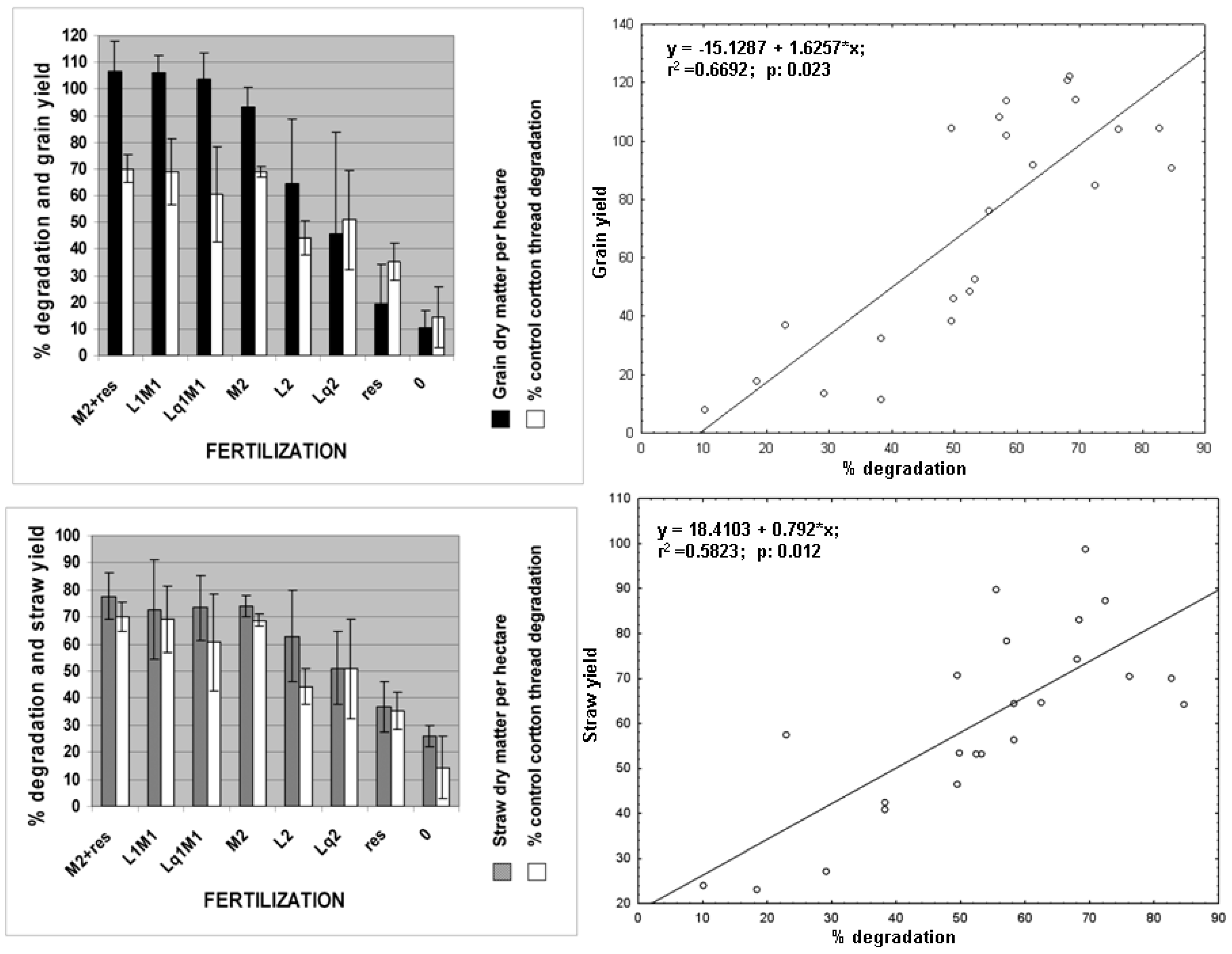
3.3. Productivity Prediction by the Fertimetro
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Anderson, J.P.E.; Domsch, K.H. Quantification of bacterial and fungal contributions to soil respiration. Arch. Microbiol. 1973, 93, 113–127. [Google Scholar] [CrossRef]
- Anderson, J.P.E.; Domsch, K.H. A physiological method for the quantitative measurement of microbial biomass in soils. Soil. Biol. Biochem. 1978, 10, 215–221. [Google Scholar] [CrossRef]
- Anderson, T.H.; Domsch, K.H. The metabolic quotient for CO2 (qCO2) as a specific activity parameter to assess the effects of environmental conditions, such as pH, on the microbial biomass of forest soils. Soil. Biol. Biochem. 1993, 25, 393–395. [Google Scholar] [CrossRef]
- Axelrood, P.E.; Chow, M.L.; Radomski, C.C.; McDermott, J.M.; Davies, J. Molecular characterization of bacterial diversity from British Columbia forest soils subjected to disturbance. Can. J. Microbiol. 2002, 48, 655–674. [Google Scholar] [CrossRef] [PubMed]
- Nannipieri, P.; Pietramellara, G.; Renella, G. Omics in Soil Science; Caster Academic Press: Norfolk, UK, 2014. [Google Scholar]
- Roesch, L.F.; Fulthorpe, R.R.; Casella, G.; Hadwin, A.K.; Kent, A.D.; Daroub, S.H.; Camargo, F.A.; Farmerie, W.G.; Triplett, E.W. Pyrosequencing enumerates and contrasts soil microbial diversity. ISME J. 2007, 1, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Heal, O.W.; Howson, G.; French, D.D.; Jeffers, J.N.R. Decomposition of cotton strips in tundra. In Soil Organisms and Decomposition in Tundra; Holding, A.J., Heal, O.W., MacLean, S.F., Flanagan, P.W., Eds.; Tundra Biome Steering Committee: Stockholm, Sweden, 1974; pp. 341–362. [Google Scholar]
- Anonymous. Methods of Test for Textiles—Woven Fabrics—Determination of Breaking Strength and Elongation Strip Method; BS 2576; British Standards Institution: London, UK, 1986. [Google Scholar]
- Harrison, A.F.; Latter, P.M.; Walton, D.W.H. (Eds.) Cotton Strip Assay an Index of Decomposition in Soils; NERC/ITE Symposium No. 24; Institute of Terrestrial Ecology, Grange-over-Sands: Cumbria, UK, 1988. [Google Scholar]
- Latter, P.M.; Walton, D.W.H. The cotton strip assay for cellulose decomposition studies in soil: History of the assay and development. In Cotton Strip Assay: An Index of Decomposition in Soils; NERC/ITE Symposium No. 24; Harrison, A.F., Latter, P.M., Walton, D.W.H., Eds.; Grange-over-Sands: Cumbria, UK, 1988; pp. 7–10. [Google Scholar]
- Semenov, A.M.; Batomunkueva, B.P.; Nizovtseva, D.V.; Panikov, N.S. Method of determination of cellulase activity in soils and in microbial cultures, and its calibration. J. Microbiol. Meth. 1996, 24, 259–267. [Google Scholar] [CrossRef]
- Correll, R.L.; Harch, B.D.; Kirkby, C.A.; O’Brien, K.; Pankhurst, C.E. Statistical analysis of reduction in tensile strength of cotton strips as a measure of soil microbial activity. J. Microbiol. Meth. 1997, 31, 9–17. [Google Scholar] [CrossRef]
- Chew, I.; Obbard, J.P.; Stanforth, R.R. Microbial cellulose decomposition in soils from a rifle range contaminated with heavy metals. Environ. Pollut. 2001, 111, 367–375. [Google Scholar] [CrossRef]
- Mendelssohn, I.A.; Slocum, M.G. Relationship between soil cellulose decomposition and oil contamination after an oil spill at Swanson Creek, Maryland. Mar. Pollut. Bull. 2004, 48, 359–370. [Google Scholar] [CrossRef]
- Nachimuthu, G.; King, K.; Kristiansen, P.; Lockwood, P.; Guppy, C. Comparison of methods for measuring soil microbial activity using cotton strips and a respirometer. J. Microbiol. Meth. 2007, 69, 322–329. [Google Scholar] [CrossRef] [PubMed]
- Concheri, G.; Tiozzo, S.; Stevanato, P.; Squartini, A. The nutrient-primed incremented substrate degradation principle. A novel method and an automated tool to assess and correct agricultural soil deficiencies to optimize its fertility and crop productivity. Appl. Soil Ecol. 2018, 123, 686–692. [Google Scholar] [CrossRef]
- Giardini, L.; Parrini, P. Concimazione e irrigazione nel Veneto. Nota I.—Rilievi produttivi del quadriennio, 1964–1967. Riv. Agron. 1969, 3, 113–119. [Google Scholar]
- Giardini, L.; Borin, M.; Berti, A. Effetti del letame e del liquame bovino in avvicendamenti colturali a diverso livello di concimazione. Riv. Agron. 1999, 33, 118–129. [Google Scholar]
- Nardi, S.; Morari, F.; Berti, A.; Tosoni, M.; Giardini, L. Soil organic matter properties after 40 years of different use of organic and mineral fertilizers. Eur. J. Agron. 2004, 21, 357–367. [Google Scholar] [CrossRef]
- Giardini, L. Potenzialità Produttività e Sostenibilità dei Sistemi Colturali, 40 Anni di Ricerche in Veneto; Patron Publisher: Bologna, Italy, 2004; pp. 81–87. ISBN 8855527487. [Google Scholar]
- Egli, T. On multiple-nutrient-limited growth of microorganisms, with special reference to dual limitation by carbon and nitrogen substrates. Ant. Leeuw. 1991, 60, 225–234. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Designation | Treatment |
|---|---|
| 0 | No fertilization |
| 0 + r | No fertilization + crop residues |
| L1 + M | Manure (30 t ha−1 year−1) + mineral fertilizer (150 N + 75 P2O5 + 210 K2O kg ha−1 year−1) |
| L2 | Manure (60 t ha−1 year−1) |
| Lq1 + M + r | Slurry (60 t ha−1 year−1) + mineral fertilizer (150 N + 75 P2O5 + 210 K2O kg ha−1 year−1) + crop residues |
| Lq2 + r | Slurry (120 t ha−1 year−1) + crop residues |
| M2 | Mineral fertilizer (300 N + 150 P2O5 + 420 K2O kg ha−1 year−1) |
| M2 + r | Mineral fertilizer (300 N + 150 P2O5 + 420 K2O kg ha−1 year−1) + crop residues |
| Corn Yield (October) | Fertimetro Data (May) | r Coefficient | p-Value |
|---|---|---|---|
| Yield of corn grain (tons per hectare) | % degradation cotton control | 0.78 | 0.0001 *** |
| Yield of corn grain (tons per hectare) | % degradation cotton + nitrogen | 0.70 | 0.0001 *** |
| Yield of corn grain (tons per hectare) | % degradation cotton + phosphorus | 0.55 | 0.0049 ** |
| Yield of corn straw (tons per hectare) | % degradation cotton control | 0.73 | 0.0001 *** |
| Fat content % in corn grain | % degradation cotton control | −0.63 | 0.0009 *** |
| Starch content % in corn grain | % degradation cotton control | 0.47 | 0.0200 * |
| % humidity in corn grain | % degradation cotton control | −0.63 | 0.0009 *** |
| Protein content % in corn grain | % degradation silk + phosphorus | −0.43 | 0.0339 * |
| Starch content % in corn grain | % degradation silk + phosphorus | 0.42 | 0.0395 * |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Concheri, G.; Tiozzo, S.; Stevanato, P.; Morari, F.; Berti, A.; Polese, R.; Borin, M.; Squartini, A. Fertimetro, a Principle and Device to Measure Soil Nutrient Availability for Plants by Microbial Degradation Rates on Differently-Spiked Buried Threads. Soil Syst. 2019, 3, 3. https://doi.org/10.3390/soilsystems3010003
Concheri G, Tiozzo S, Stevanato P, Morari F, Berti A, Polese R, Borin M, Squartini A. Fertimetro, a Principle and Device to Measure Soil Nutrient Availability for Plants by Microbial Degradation Rates on Differently-Spiked Buried Threads. Soil Systems. 2019; 3(1):3. https://doi.org/10.3390/soilsystems3010003
Chicago/Turabian StyleConcheri, Giuseppe, Stefano Tiozzo, Piergiorgio Stevanato, Francesco Morari, Antonio Berti, Riccardo Polese, Maurizio Borin, and Andrea Squartini. 2019. "Fertimetro, a Principle and Device to Measure Soil Nutrient Availability for Plants by Microbial Degradation Rates on Differently-Spiked Buried Threads" Soil Systems 3, no. 1: 3. https://doi.org/10.3390/soilsystems3010003