
Marine Natural Product Antimycin A Suppresses Wheat Blast Disease Caused by Magnaporthe oryzae Triticum


 ,
,  ,
,  ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
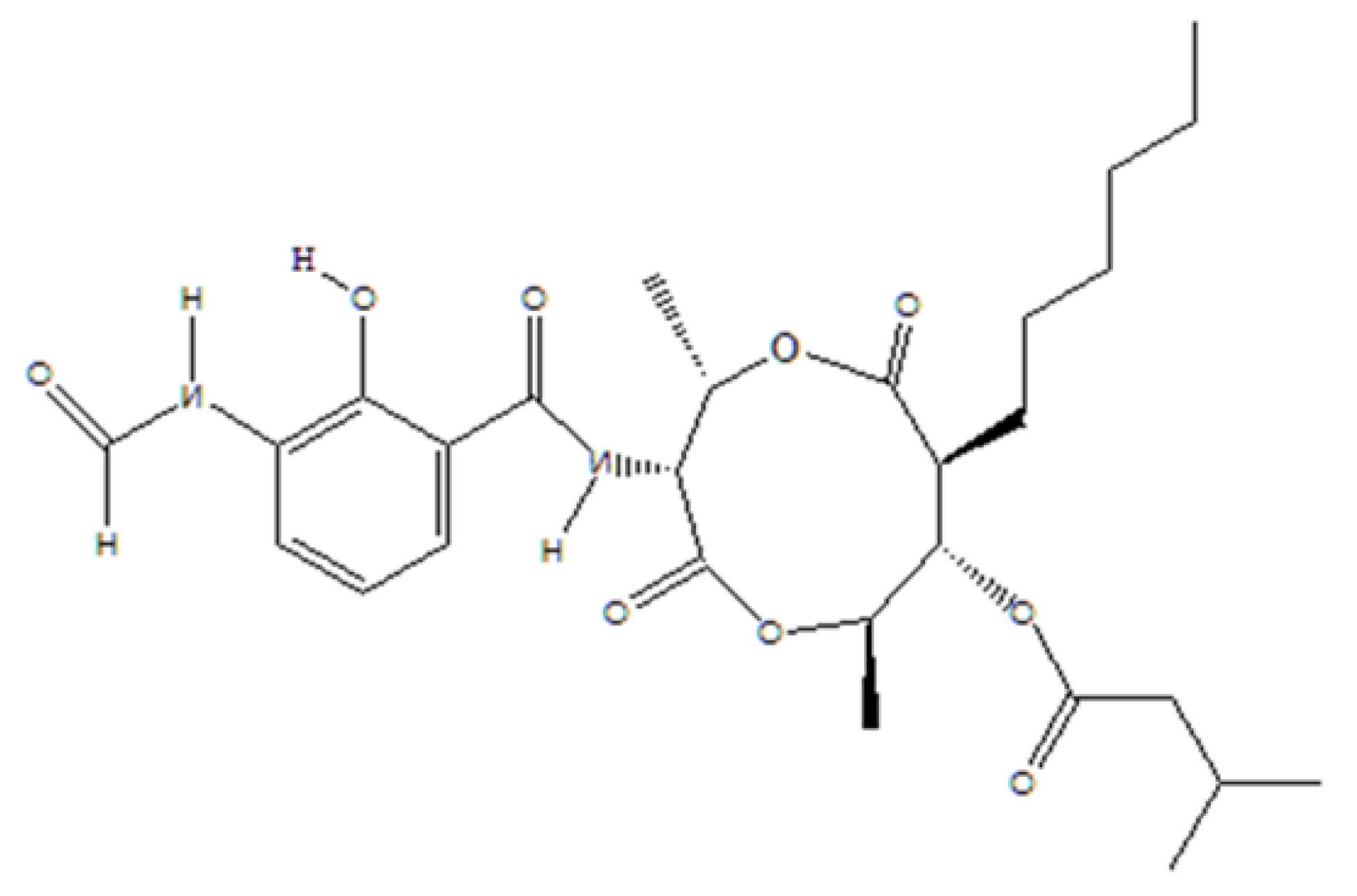
2.1. Chemicals
2.2. Culture of Wheat Blast Isolate
2.3. Preparation of Chemical Solution and Conidial Suspension
2.4. Fungal Growth Inhibition and Morphological Changes of Hyphae
2.5. Inhibition of Conidiogenesis
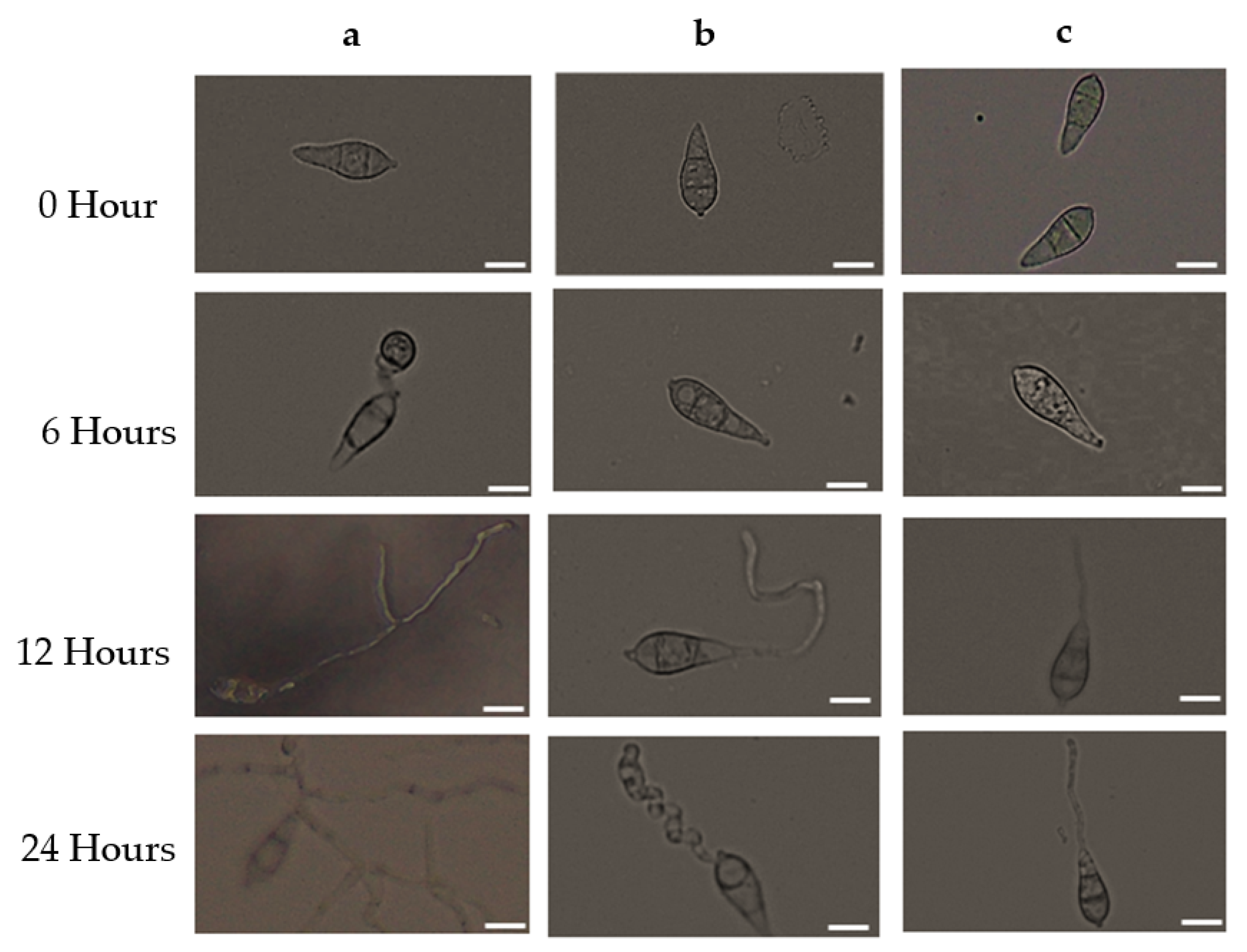
2.6. Germination Inhibition and Morphological Modifications of Germinated Conidia
2.7. Growing of Seedlings
2.8. Field Evaluation of Antimycin A against Wheat Blast
2.8.1. Preparation of Land and Fertilization
2.8.2. Seed Sowing and Management of the Plot
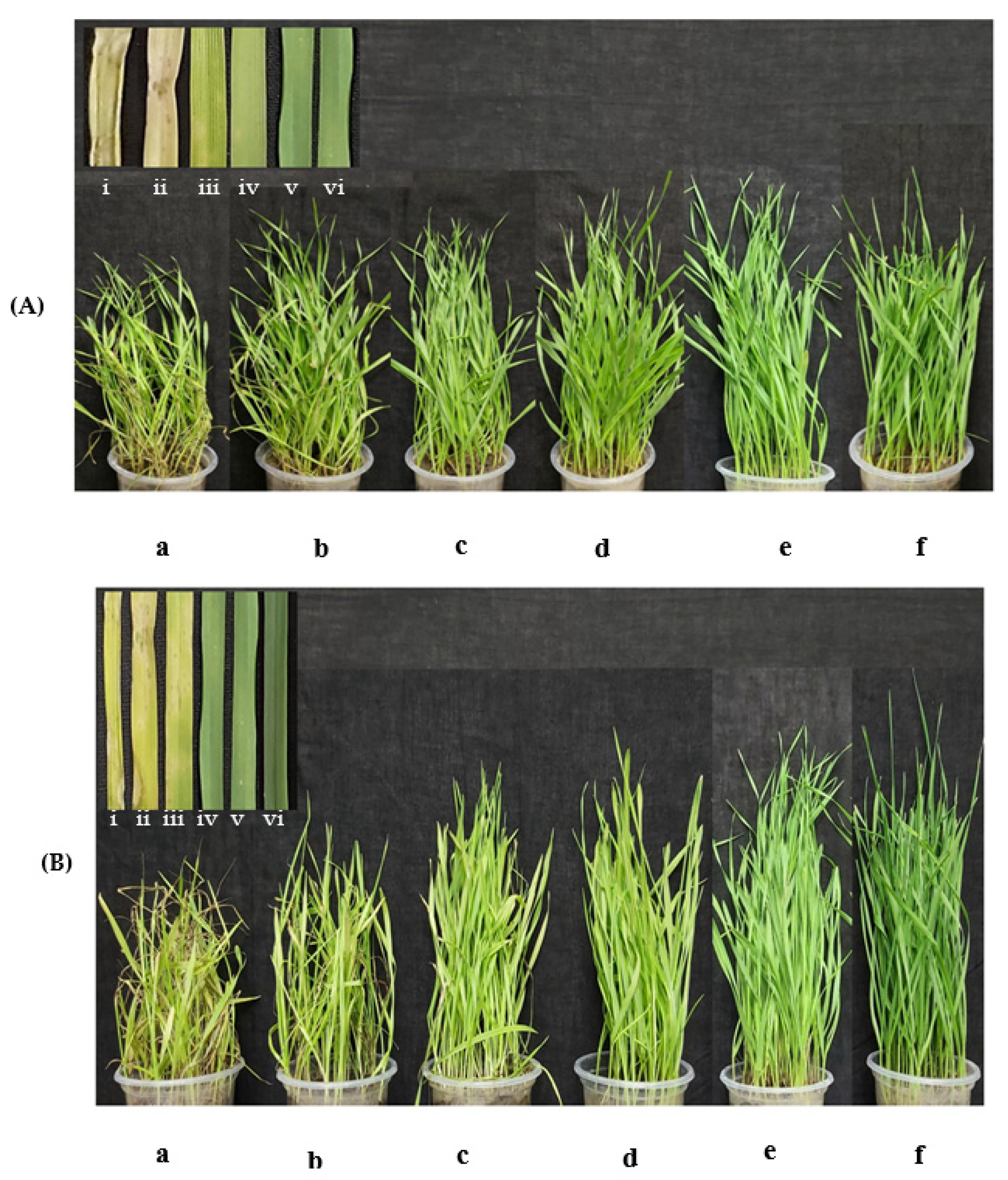
2.9. Plant Infection Assay at Seedling Stage
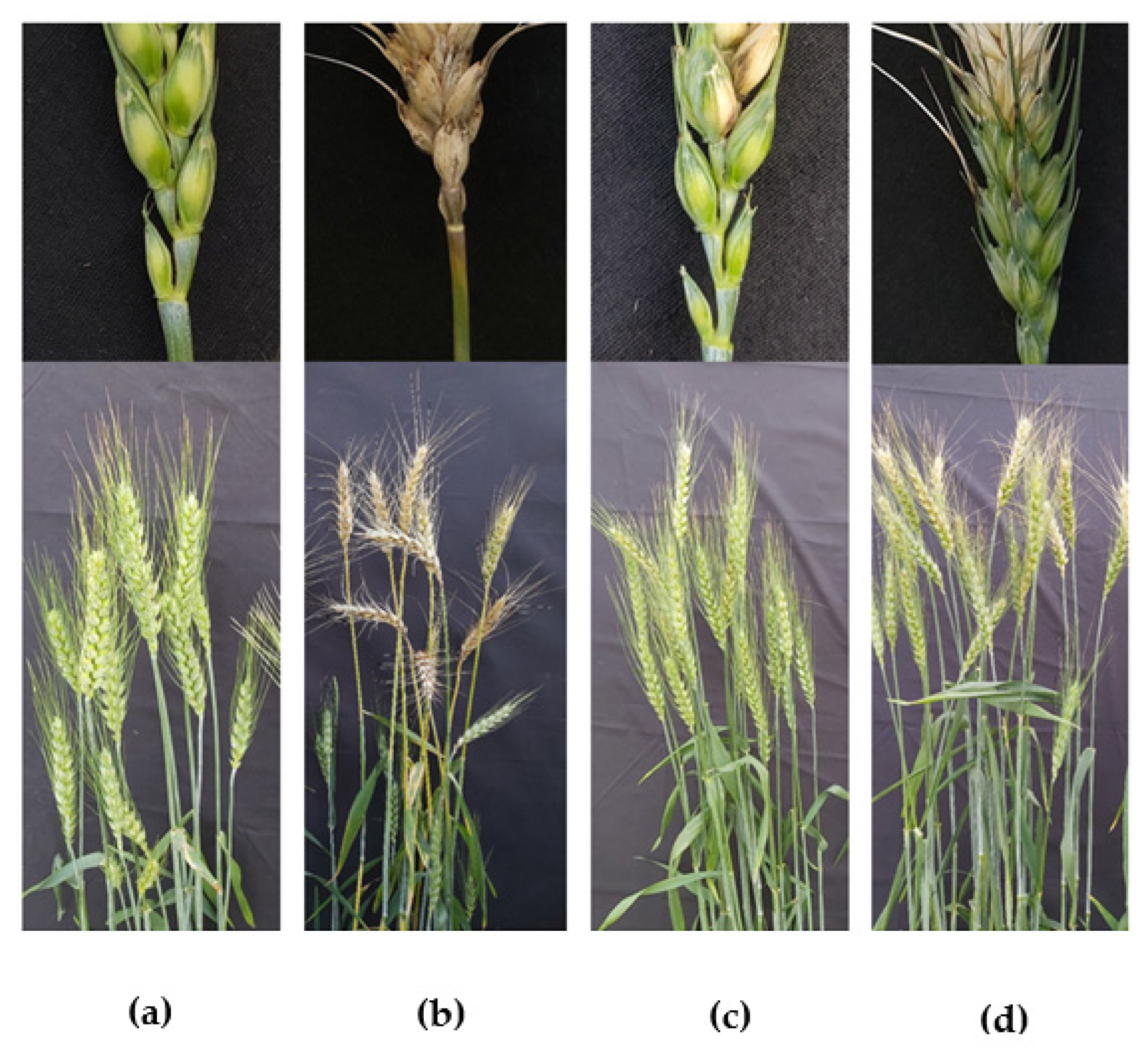
2.10. Infection Assay in Wheat Field at Reproductive Phase
2.11. Recording of Data, Measurement of Disease Intensity and Severity
2.12. Design of Experiment and Statistical Analysis
3. Results
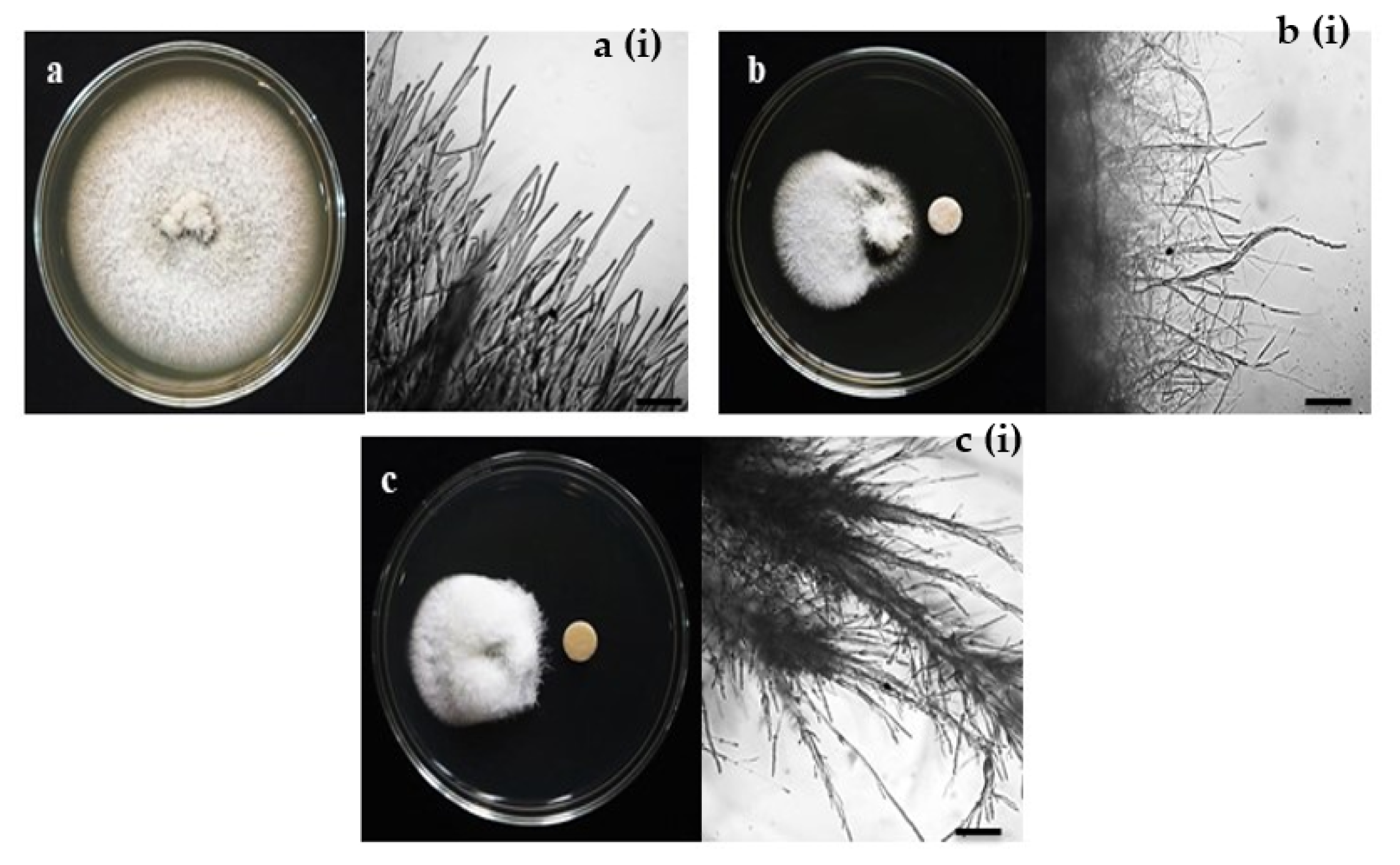
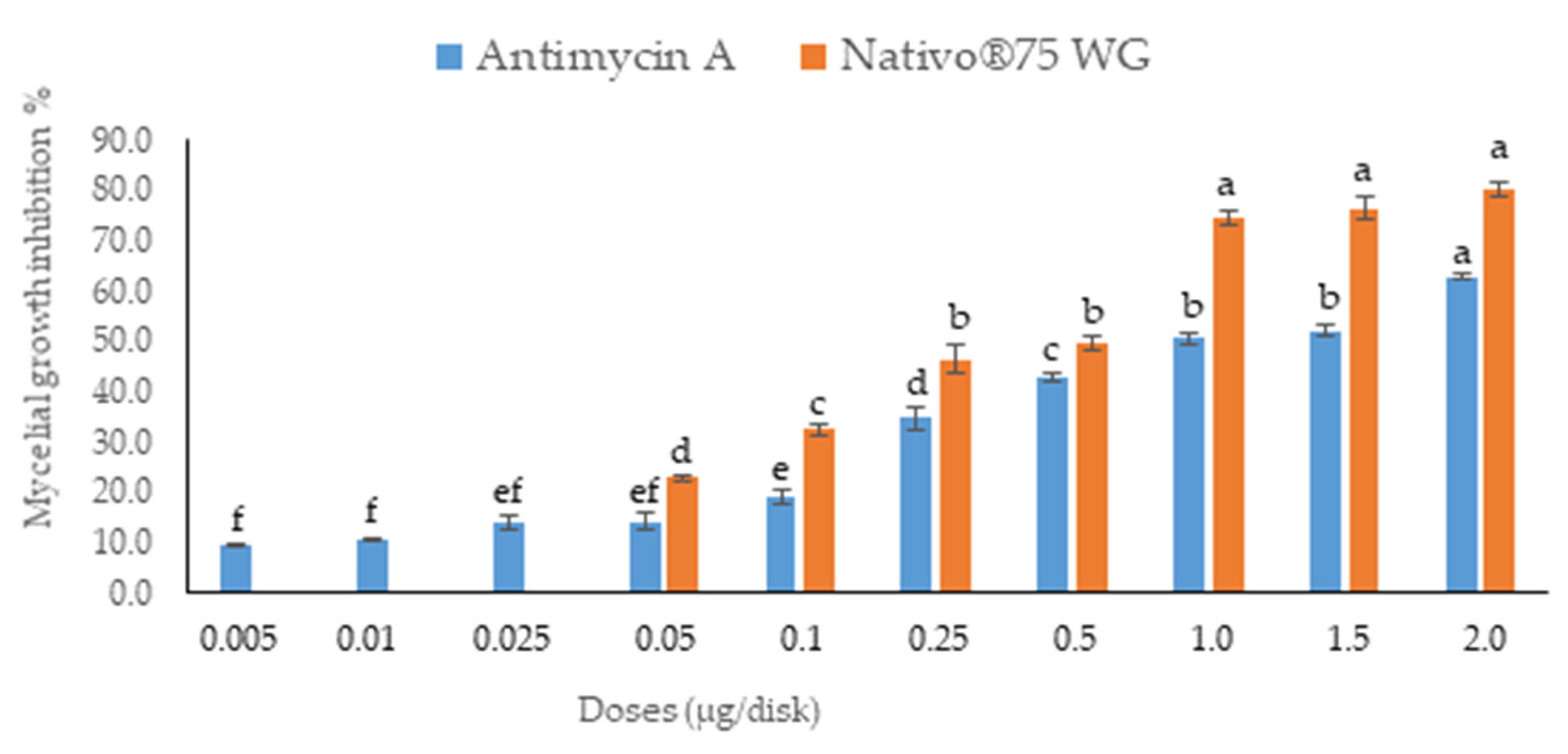
3.1. In Vitro Assays of Fungal Growth Inhibition
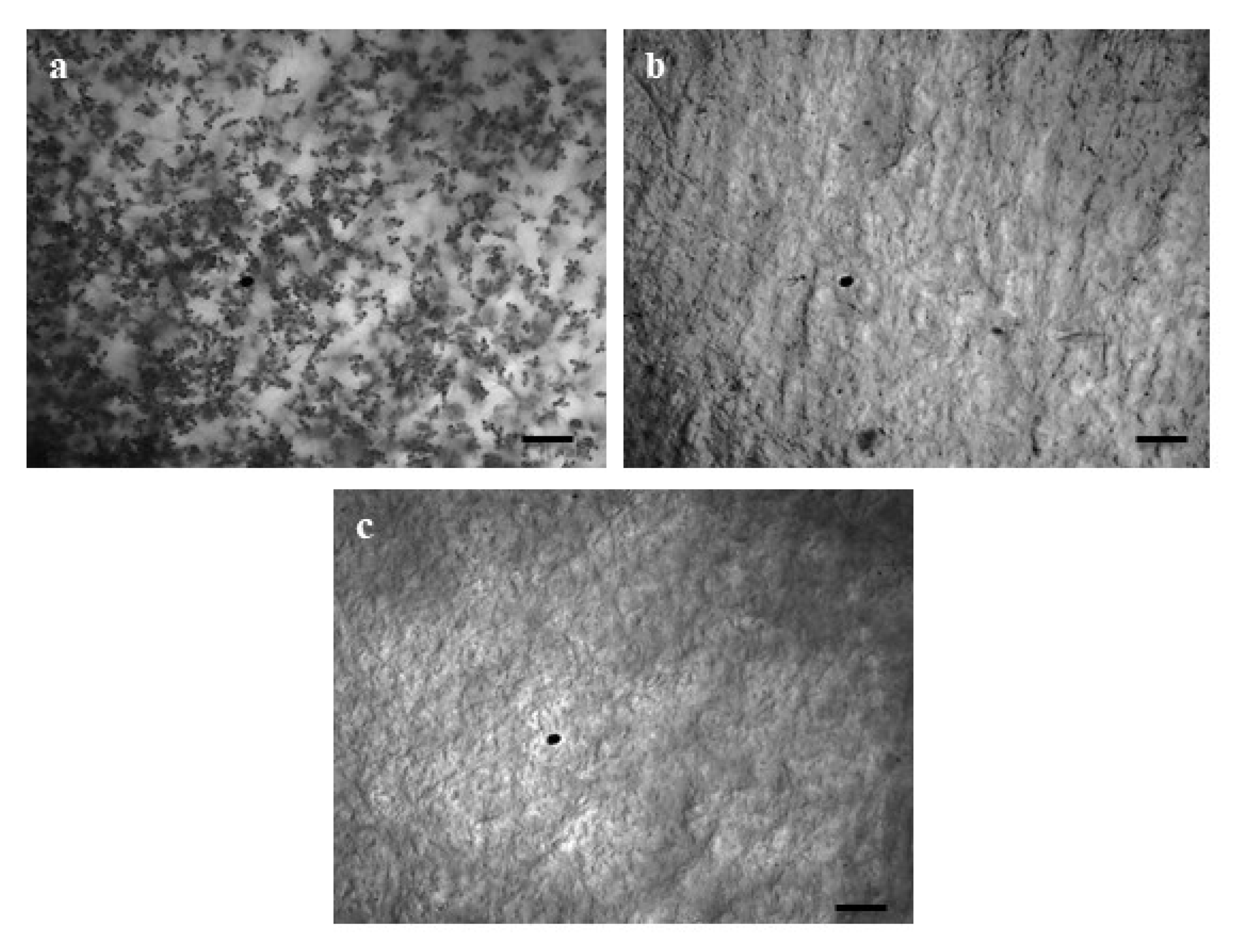
3.2. Antimycin A Block Conidiogenesis in MoT
3.3. Antimycin A Alters Conidia Germination and Developmental Transitions of Germinated Conidia
3.4. Antimycin A Suppresses Wheat Blast Disease at Seedling Stage
3.5. Suppression of Wheat Blast Disease by Antimycin A at Heading Stage of Wheat under Field Conditions
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Igarashi, S.; Utiamada, C.M.; Igarashi, L.C.; Kazuma, A.H.; Lopes, R.S. Pyricularia em trigo. 1. Ocorrencia de Pyricularia sp noestado do Parana. Phytopathol. Bras. 1986, 11, 351–352. [Google Scholar]
- Kohli, M.M.; Mehta, Y.R.; Guzman, E.; Viedma, L.; Cubilla, L.E. Pyricularia blast-a threat to wheat cultivation. Czech J. Genet. Plant Breed. 2011, 47, 130–134. [Google Scholar] [CrossRef] [Green Version]
- Islam, M.T.; Croll, D.; Gladieux, P.; Soanes, D.M.; Persoons, A.; Bhattacharjee, P.; Hossain, M.S.; Gupta, D.R.; Rahman, M.M.; Mahboob, M.G.; et al. Emergence of wheat blast in Bangladesh was caused by a South American lineage of Magnaporthe oryzae. BMC Biol. 2016, 14, 84. [Google Scholar] [CrossRef] [Green Version]
- Islam, M.T.; Kim, K.H.; Choi, J. Wheat blast in Bangladesh: The current situation and future impacts. Plant Pathol. J. 2019, 35, 1. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.T.; Gupta, D.R.; Hossain, A.; Roy, K.K.; He, X.; Kabir, M.R.; Singh, P.K.; Khan, M.; Rahman, A.; Rahman, M.; et al. Wheat blast: A new threat to food security. Phytopathol. Res. 2020, 2, 28. [Google Scholar] [CrossRef]
- Kamoun, S.; Talbot, N.J.; Islam, M.T. Plant health emergencies demand open science: Tackling a cereal killer on the run. PLoS Biol. 2019, 17, e3000302. [Google Scholar] [CrossRef] [PubMed]
- Tembo, B.; Mulenga, R.M.; Sichilima, S.; M’siska, K.K.; Mwale, M.; Chikoti, P.C.; Singh, P.K.; He, X.; Pedley, K.F.; Peterson, G.L.; et al. Detection and characterization of fungus (Magnaporthe oryzae pathotype Triticum) causing wheat blast disease on rain-fed grown wheat (Triticum aestivum L.) in Zambia. PLoS ONE 2020, 15, e0238724. [Google Scholar] [CrossRef] [PubMed]
- Ceresini, P.C.; Castroagudı’n, V.L.; Rodriguez, F.A.; Rios, J.A.; Aucique-Pe´rez, C.E.; Moreira, S.I. Wheat blast: From its origin in South America to its emergence as a global threat. Mol. Plant Pathol. 2019, 20, 155–172. [Google Scholar] [CrossRef]
- Inoue, K.; Suzuki, T.; Ikeda, K.; Jiang, S.; Hosogi, N.; Hyon, G.S. Extracellular matrix of Magnaporthe oryzae may have a role in host adhesion during fungal penetration and is digested by matrix metalloproteinases. J. Gen. Plant Pathol. 2007, 73, 388–398. [Google Scholar] [CrossRef]
- Urashima, A.S.; Hashimoto, Y.; Le Don, D.; Kusaba, M.; Tosa, Y.; Nakayashiki, H.; Mayama, S. Molecular analysis of the wheat blast population in Brazil with a homolog of retrotransposon MGR583. Jpn. J. Phytopathol. 1999, 65, 429–436. [Google Scholar] [CrossRef] [Green Version]
- Singh, P.K.; Gahtyari, N.C.; Roy, C.; Roy, K.K.; He, X.; Tembo, B.; Xu, K.; Juliana, P.; Sonder, K.; Kabir, M.R.; et al. Wheat blast: A disease spreading by intercontinental jumps and its management strategies. Front. Plant Sci. 2021, 12, 710707. [Google Scholar] [CrossRef]
- Hale, A.T.; Graham, R.D.M.; Andreas, M.; Jean-Benoit, M.; Lucie, M.; Lesley, A.B. Wheat blast: Histopathology and transcriptome reprogramming in response to adapted and non-adapted Magnaporthe isolates. New Phytol. 2009, 184, 473–484. [Google Scholar]
- Wilson, R.A.; Talbot, N.J. Under pressure: Investigating the biology of plant infection by Magnaporthe oryzae. Nat. Rev. Microbiol. 2009, 7, 185–195. [Google Scholar] [CrossRef]
- Chakraborty, M.; Mahmud, N.U.; Muzahid, A.N.M.; Rabby, S.M.F.; Islam, T. Oligomycins inhibit Magnaporthe oryzae Triticum and suppress wheat blast disease. PLoS ONE 2020, 15, e0233665. [Google Scholar] [CrossRef]
- Castroagudín, V.L.; Ceresini, P.C.; de Oliveira, S.C.; Reges, J.T.; Maciel, J.L.; Bonato, A.L.; Dorigan, A.F.; McDonald, B.A. Resistance to QoI fungicides is widespread in Brazilian populations of the wheat blast pathogen Magnaporthe oryzae. Phytopathology 2015, 105, 284–294. [Google Scholar] [CrossRef] [Green Version]
- Cruz, C.D.; Valent, B. Wheat blast disease: Danger on the move. Trop. Plant Pathol. 2017, 42, 210–222. [Google Scholar] [CrossRef] [Green Version]
- Poloni, N.M.; Carvalho, G.; Nunes Campos Vicentini, S.; Francis Dorigan, A.; Nunes Maciel, J.L.; McDonald, B.A.; Intra Moreira, S.; Hawkins, N.; Fraaije, B.A.; Kelly, D.E.; et al. Widespread distribution of resistance to triazole fungicides in Brazilian populations of the wheat blast pathogen. Plant Pathol. 2021, 70, 436–448. [Google Scholar] [CrossRef]
- Petriccione, M.; Mastrobuoni, F.; Zampella, L.; Nobis, E.; Capriolo, G.; Scortichini, M. Effect of chitosan treatment on strawberry allergen-related gene expression during ripening stages. J. Food Sci. Technol. 2017, 54, 1340–1345. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez, M.; Núñez, F. Novel approaches to minimizing mycotoxin contamination. Toxins 2020, 12, 216. [Google Scholar] [CrossRef] [Green Version]
- Figueroa, E.; Lee-Estévez, M.; Valdebenito, I.; Farías, J.G.; Romero, J. Potential biomarkers of DNA quality in cryopreserved fish sperm: Impact on gene expression and embryonic development. Rev. Aquac. 2020, 12, 382–391. [Google Scholar] [CrossRef]
- Wulff, B.B.; Dhugga, K.S. Wheat-the cereal abandoned by GM. Science 2018, 361, 451–452. [Google Scholar] [CrossRef]
- Chakraborty, M.; Mahmud, N.U.; Gupta, D.R.; Tareq, F.S.; Shin, H.J.; Islam, T. Inhibitory effects of linear lipopeptides from a marine Bacillus subtilis on the wheat blast fungus Magnaporthe oryzae Triticum. Front. Microbiol. 2020, 11, 665. [Google Scholar] [CrossRef]
- Ōmura, S.; Ikeda, H.; Ishikawa, J.; Hanamoto, A.; Takahashi, C.; Shinose, M.; Takahashi, Y.; Horikawa, H.; Nakazawa, H.; Osonoe, T.; et al. Genome sequence of an industrial microorganism Streptomyces avermitilis: Deducing the ability of producing secondary metabolites. Proc. Natl. Acad. Sci. USA 2001, 98, 12215–12220. [Google Scholar] [CrossRef] [Green Version]
- Mehdi, R.B.A.; Sioud, S.; Fguira, L.F.B.; Bejar, S.; Mellouli, L. Purification and structure determination of four bioactive molecules from a newly isolated Streptomyces sp. TN97 strain. Process Biochem. 2006, 41, 1506–1513. [Google Scholar] [CrossRef]
- Kaur, T.; Kaur, A.; Sharma, V.; Manhas, R.K. Purification and Characterization of a New Antifungal Compound 10-(2, 2-dimethyl-cyclohexyl)-6, 9-dihydroxy-4, 9-dimethyl-dec-2-enoic Acid Methyl Ester from Streptomyces hydrogenans Strain DH16. Front. Microbiol. 2016, 7, 1004. [Google Scholar] [CrossRef]
- Doumbou, C.L.; Salove, M.K.H.; Crawford, D.L.; Beaulieu, C. Actinomycetes, promising tools to control plant diseases and to promote plant growth. Phytoprotection 2001, 82, 85–102. [Google Scholar] [CrossRef] [Green Version]
- Xiong, Z.Q.; Zhang, Z.P.; Li, J.H.; Wei, S.J.; Tua, G.Q. Characterization of Streptomyces padanus JAU4234, a producer of actinomycin X2, fungichromin, and a new polyene macrolide antibiotic. Appl. Environ. Microbiol. 2012, 78, 589–592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palaniyandi, S.A.; Yang, S.H.; Zhang, L.; Suh, J.W. Effects of actinobacteria on plant disease suppression and growth promotion. Appl. Microbiol. Biotechnol. 2013, 97, 9621–9636. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, X.H.; Naing, K.W.; Lee, Y.S.; Kim, Y.H.; Moon, J.H.; Kim, K.Y. Antagonism of antifungal metabolites from Streptomyces griseus H7602 against Phytophthora capsici. J. Basic Microbiol. 2015, 55, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.T.; Monkhung, S.; Lee, Y.S.; Kim, K.Y. Effects of Lysobacter antibioticus HS124, an effective biocontrol agent against Fusarium graminearum, on crown rot disease and growth promotion of wheat. Can. J. Microbiol. 2019, 65, 904–912. [Google Scholar] [CrossRef] [PubMed]
- Fathi, F.; Saberi-Riseh, R.; Khodaygan, P. Survivability and controlled release of alginate-microencapsulated pseudomonas fluorescens vupf506 and their effects on biocontrol of Rhizoctonia solani on potato. Int. J. Biol. Macromol. 2021, 183, 627–634. [Google Scholar] [CrossRef]
- Moradi-Pour, M.; Saberi-Riseh, R.; Mohammadinejad, R.; Hosseini, A. Investigating the formulation of alginate-gelatin encapsulated Pseudomonas fluorescens (VUPF5 and T17-4 strains) for controlling Fusarium solani on potato. Int. J. Biol. Macromol. 2019, 133, 603–613. [Google Scholar] [CrossRef]
- Taechowisan, T.; Peberdy, J.F.; Lumyong, S. Chitinase production by endophytic Streptomyces aureofaciens CMUAc130 and its antagonism against phytopathogenic fungi. Ann. Microbiol. 2003, 53, 447–461. [Google Scholar]
- Taechowisan, T.; Lu, C.; Shen, Y.; Lumyong, S. Secondary metabolites from endophytic Streptomyces aureofaciens CMUAc130 and their antifungal activity. Microbiology 2005, 151, 1691–1695. [Google Scholar] [CrossRef] [Green Version]
- Quecine, M.C.; Araujo, W.L.; Marcon, J.; Gai, C.S.; Azevedo, J.L.; Pizzirani-Kleiner, A.A. Chitinolytic activity of endophytic Streptomyces and potential for biocontrol. Lett. Appl. Microbiol. 2008, 47, 486–491. [Google Scholar] [CrossRef]
- Yuan, W.M.; Crawford, D.L. Characterization of Streptomyces lydicus WYEC108 as a potential biocontrol agent against fungal root and seed rots. Appl. Environ. Microbiol. 1995, 61, 3119–3128. [Google Scholar] [CrossRef] [Green Version]
- Tahvonen, R.; Avikainen, H. The biological control of seed-borne Alternaria brassicicola of cruciferous plants with a powdery preparation of Streptomyces sp. Agric. Food Sci. 1987, 59, 199–208. [Google Scholar] [CrossRef]
- Law, J.W.F.; Ser, H.L.; Khan, T.M.; Chuah, L.H.; Pusparajah, P.; Chan, K.G. The potential of Streptomyces as biocontrol agents against the rice blast fungus, Magnaporthe oryzae (Pyricularia oryzae). Front. Microbiol. 2017, 8, 3. [Google Scholar] [CrossRef] [Green Version]
- Bubici, G. Streptomyces spp. as biocontrol agents against Fusarium species. CAB Rev. Perspective. Agric. Vet. Sci. Nutr. Nat. Resour. 2018, 13, 1–15. [Google Scholar] [CrossRef]
- Santra, H.K.; Maity, S.; Banerjee, D. Production of bioactive compounds with broad spectrum bactericidal action, bio-film inhibition and antilarval potential by the secondary metabolites of the endophytic fungus Cochliobolus sp. APS1 Isolated from the Indian Medicinal Herb Andrographis paniculata. Molecules 2022, 27, 1459. [Google Scholar]
- Mukku, V.J.; Speitling, M.; Laatsch, H.; Helmke, E. New butenolides from two marine streptomycetes. J. Nat. Prod. 2000, 63, 1570–1572. [Google Scholar] [CrossRef] [Green Version]
- Hosotani, N.; Kumagai, K.; Nakagawa, H.; Shimatani, T.; Saji, I. Antimycins A10 approximately A16, seven new antimycin antibiotics produced by Streptomyces spp. SPA-10191 and SPA-8893. J. Antibiot. 2005, 58, 460–467. [Google Scholar] [CrossRef]
- Rieske, J.S. Antimycin A. In Antibiotics; Springer: Berlin/Heidelberg, Germany, 1967; pp. 542–584. [Google Scholar]
- Nafis, A.; Elhidar, N.; Oubaha, B.; Samri, S.E.; Niedermeyer, T.; Ouhdouch, Y.; Hassani, L.; Barakate, M. Screening for Non-polyenic Antifungal Produced by Actinobacteria from Moroccan Habitats: Assessment of Antimycin A19 Production by Streptomyces albidoflavus AS25. Int. J. Mol. Cell. Med. 2018, 7, 133–145. [Google Scholar]
- Yan, L.L.; Han, N.N.; Zhang, Y.Q.; Yu, L.Y.; Chen, J.; Wei, Y.Z.; Li, Q.P.; Tao, L.; Zheng, G.H.; Yang, S.E.; et al. Antimycin A18 produced by an endophytic Streptomyces albidoflavus isolated from a mangrove plant. J. Antibiot. 2010, 63, 259–261. [Google Scholar] [CrossRef]
- Belakhov, V.V.; Boikova, I.V.; Novikova, I.I.; Kolodyaznaya, V.A. Results of examination of the biological activity of nonmedical antibiotics with a view to finding environmentally friendly pesticides for plant protection. Russ. J. Gen. Chem. 2018, 88, 2982–2989. [Google Scholar] [CrossRef]
- Gupta, D.R.; Surovy, M.Z.; Mahmud, N.U.; Chakraborty, M.; Paul, S.K.; Hossain, M.; Bhattacharjee, P.; Mehebub, M.; Rani, K.; Yeasmin, R.; et al. Suitable methods for isolation, culture, storage and identification of wheat blast fungus Magnaporthe oryzae Triticum pathotype. Phytopathol. Res. 2020, 2, 30. [Google Scholar] [CrossRef]
- Bauer, A.W.; Kirby, W.M.; Sherris, J.C.; Turck, M. Antibiotic susceptibility testing by a standardized single disk method. Am. J. Clin. Pathol. 1966, 45, 493–496. [Google Scholar] [CrossRef]
- Riungu, G.M.; Muthorni, J.W.; Narla, R.D.; Wagacha, J.M.; Gathumbi, J.K. Management of Fusarium head blight of wheat and deoxynivalenol accumulation using antagonistic microorganisms. Plant Pathol. J. 2008, 7, 13–19. [Google Scholar] [CrossRef] [Green Version]
- Urashima, A.S.; Igarashi, S.; Kato, H. Host range, mating type, and fertility of Pyricularia grisea from wheat in Brazil. Plant Dis. 1993, 77, 1211–1216. [Google Scholar] [CrossRef]
- Islam, M.T.; von Tiedemann, A. 2,4-Diacetylphloroglucinol suppresses zoosporogenesis and impairs motility of Peronosporomycete zoospores. World J. Microb. Biotechnol. 2011, 27, 2071–2079. [Google Scholar] [CrossRef] [Green Version]
- Robinson, R.J.; Fraaije, B.A.; Clark, I.M.; Jackson, R.W.; Hirsch, P.R.; Mauchline, T.H. Wheat seed embryo excision enables the creation of axenic seedlings and Koch’s postulates testing of putative bacterial endophytes. Sci. Rep. 2016, 6, 25581. [Google Scholar] [CrossRef] [PubMed]
- Bangladesh Agricultural Research Council (BARC). Fertilizer Recommendation Guide (FRG); Bangladesh Agricultural Research Council (BARC): Dhaka, Bangladesh, 2012; pp. 1–265. [Google Scholar]
- Ha, X.; Koopmann, B.; von Tiedemann, A. Wheat blast and Fusarium head blight display contrasting interaction patterns on ears of wheat genotypes differing in resistance. Phytopathology 2016, 106, 270–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turrens, J.F.; Alexandre, A.; Lehninger, A.L. Ubisemiquinone is the electron donor for superoxide formation by complex III of heart mitochondria. Arch. Biochem. Biophys. 1985, 237, 408–414. [Google Scholar] [CrossRef]
- Turrens, J.F. Mitochondrial formation of reactive oxygen species. Physiol. J. 2003, 552, 335–344. [Google Scholar] [CrossRef]
- Formigli, L.; Papucci, L.; Tani, A.; Schiavone, N.; Tempestini, A.; Orlandini, G.E.; Capaccioli, S.; Zecchi Orlandini, S. Aponecrosis: Morphological and biochemical exploration of a syncretic process of cell death sharing apoptosis and necrosis. J. Cell. Physiol. 2000, 182, 41–49. [Google Scholar] [CrossRef]
- Park, W.H.; Han, Y.W.; Kim, S.H.; Kim, S.Z. An ROS generator, antimycin A, inhibits the growth of HeLa cells via apoptosis. J. Cell. Biochem. 2007, 102, 98–109. [Google Scholar] [CrossRef]
- Han, Y.W.; Kim, S.Z.; Kim, S.H.; Park, W.H. The changes of intracellular H2O2 are an important factor maintaining mitochondria membrane potential of antimycin A-treated As4. 1 juxtaglomerular cells. Biochem. Pharmacol. 2007, 73, 863–872. [Google Scholar] [CrossRef]
- Han, Y.H.; Moon, H.J.; You, B.R.; Kim, S.Z.; Kim, S.H.; Park, W.H. p38 inhibitor intensified cell death in antimycin A-treated As4. 1 juxtaglomerular cells via the enhancement of GSH depletion. Anticancer Res. 2009, 29, 4423–4431. [Google Scholar]
- Park, W.H.; You, B.R. Antimycin A induces death of the human pulmonary fibroblast cells via ROS increase and GSH depletion. Int. J. Oncol. 2016, 48, 813–820. [Google Scholar] [CrossRef] [Green Version]
- Nakayama, K.; Okamoto, F.; Harada, Y. Antimycin A: Isolation from a new Streptomyces and activity against rice plant blast fungi. J. Antibiot. Res. 1956, 9, 63–66. [Google Scholar]
- Pham, N.A.; Robinson, B.H.; Hedley, D.W. Simultaneous detection of mitochondrial respiratory chain activity and reactive oxygen in digitonin-permeabilized cells using flow cytometry. Cytom. J. Int. Soc. Anal. Cytol. 2000, 41, 245–251. [Google Scholar] [CrossRef]
- King, M.A.; Radicchi-Mastroianni, M.A. Antimycin A-induced apoptosis of HL-60 cells. Cytom. J. Int. Soc. Anal. Cytol. 2002, 49, 106–112. [Google Scholar] [CrossRef]
- Cai, J.; Jones, D.P. Superoxide in apoptosis: Mitochondrial generation triggered by cytochromec loss. J. Biol. Chem. 1998, 273, 11401–11404. [Google Scholar] [CrossRef] [Green Version]
- King, M.A. Antimycin A-induced killing of HL-60 cells: Apoptosis initiated from within mitochondria does not necessarily proceed via caspase 9. Cytom. J. Int. Soc. Anal. Cytol. 2005, 63, 69–76. [Google Scholar] [CrossRef]
- Wolvetang, E.J.; Johnson, K.L.; Krauer, K.; Ralph, S.J.; Linnane, A.W. Mitochondrial respiratory chain inhibitors induce apoptosis. FEBS Lett. 1994, 339, 40–44. [Google Scholar] [CrossRef] [Green Version]
- Liao, J.H.; Chen, P.Y.; Yang, Y.L.; Kan, S.C.; Hsieh, F.C.; Liu, Y.C. Clarification of the antagonistic effect of the lipopeptides produced by Bacillus amyloliquefaciens BPD1 against Pyricularia oryzae via in situ MALDI-TOF IMS analysis. Molecules 2016, 21, 1670. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Sun, C. Fengycins, cyclic lipopeptides from marine Bacillus subtilis strains, kill the plant-pathogenic fungus Magnaporthe grisea by inducing reactive oxygen species production and chromatin condensation. Appl. Environ. Microbiol. 2018, 84, e00445-18. [Google Scholar] [CrossRef] [Green Version]
- Isono, K.; Nagatsu, J.; Kawashima, Y.; Suzuki, S. Studies on polyoxins, antifungal antibiotics. Agric. Biol. Chem. 1965, 29, 848–854. [Google Scholar]
- Tang, Q.; Bie, X.; Lu, Z.; Lv, F.; Tao, Y.; Qu, X. Effects of fengycin from Bacillus subtilis fmbJ on apoptosis and necrosis in Rhizopus stolonifer. J. Microbiol. 2014, 52, 675–680. [Google Scholar] [CrossRef]
- Nielsen, T.H.; Thrane, C.; Christophersen, C.; Anthoni, U.; Sørensen, J. Structure, production characteristics and fungal antagonism of tensin–a new antifungal cyclic lipopeptide from Pseudomonas fluorescens strain 96.578. J. Appl. Microbiol. 2000, 89, 992–1001. [Google Scholar] [CrossRef]
- Islam, M.T.; Hashidoko, Y.; Deora, A.; Ito, T.; Tahara, S. Suppression of damping-off disease in host plants by the rhizoplane bacterium Lysobacter sp. strain SB-K88 is linked to plant colonization and antibiosis against soilborne Peronosporomycetes. Appl. Environ. Microbiol. 2005, 71, 3786–3796. [Google Scholar] [CrossRef] [Green Version]
- Islam, M.T. Disruption of ultrastructure and cytoskeletal network is involved with biocontrol of damping off pathogen Aphanomyces cochlioides by Lysobacter sp. strain SB-K88. Biol. Control. 2008, 46, 312–321. [Google Scholar] [CrossRef]
- Islam, M.T. Mode of antagonism of a biocontrol bacterium Lysobacter sp. SB-K88 toward a damping-off pathogen Aphanomyces cochlioides. World J. Microb. Biotechnol. 2010, 26, 629–637. [Google Scholar] [CrossRef]
- Islam, M.T.; Fukushi, Y. Growth inhibition and excessive branching in Aphanomyces cochlioides induced by 2,4-diacetylphloroglucinol is linked to disruption of filamentous actin cytoskeleton in the hyphae. World J. Microb. Biotechnol. 2010, 26, 1163–1170. [Google Scholar] [CrossRef]
- Kopecká, M.; Ilkovics, L.; Ramı´kova´, V.; Yamaguchi, M. Effect of cytoskeleton inhibitors on conidiogenesis and capsule in the long neck yeast Fellomyces examined by scanning electron microscopy. Chemotherapy 2010, 56, 197–202. [Google Scholar] [CrossRef]
- Ohara, T. REN1 is required for development of microconidia and macroconidia, but not of chlamydospores, in the plant pathogenic fungus Fusarium oxysporum. Genetics 2004, 166, 113–124. [Google Scholar] [CrossRef] [Green Version]
- Lyu, A.; Liu, H.; Che, H.; Yang, L.; Zhang, J.; Wu, M.; Chen, W.; Li, G. Reveromycins A and B from Streptomyces sp. 3–10: Antifungal activity against plant pathogenic fungi in vitro and in a strawberry food model system. Front. Microbiol. 2017, 8, 550. [Google Scholar] [CrossRef]
- Zhang, N.; Song, L.; Xu, Y.; Pei, X.; Luisi, B.F.; Liang, W. The decrotonylase FoSir5 facilitates mitochondrial metabolic state switching in conidial germination of Fusarium oxysporum. eLife 2021, 10, e75583. [Google Scholar] [CrossRef]
- Tzung, S.P.; Kim, K.M.; Basañez, G.; Giedt, C.D.; Simon, J.; Zimmerberg, J.; Zhang, K.Y.; Hockenbery, D.M. Antimycin A mimics a cell-death-inducing Bcl-2 homology domain 3. Nat. Cell Biol. 2001, 3, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Barhoom, S.; Sharon, A. Bcl-2 proteins link programmed cell death with growth and morphogenetic adaptations in the fungal plant pathogen Colletotrichum gloeosporioides. Fungal Genet. Biol. 2007, 44, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Pring, R.J. Effects of triadimefon on the ultrastructure of rust fungi infecting leaves of wheat and broad bean (Vicia faba). Pestic. Biochem. Phys. 1984, 21, 127–137. [Google Scholar] [CrossRef]
- Sauter, H.; Steglich, W.; Anke, T. Strobilurins: Evolution of a new class of active substances. Angew. Chem. Int. Ed. Engl. 1999, 38, 1328–1349. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Time (h) | Germination of Conidia, Morphology of Germ Tubes, and Appressorial Formation | |
|---|---|---|---|
| Germinated Conidia (% ± SE a) | Morphological Change/Developmental Transitions in the Treated Conidia | ||
| Water | 0 | 0.0 ± 0.00 b | No germination |
| 6 | 100.0 ± 0.00 a | Germination with normal germ tube and normal appressoria | |
| 12 | 100.0 ± 0.00 a | Normal mycelial growth | |
| 24 | 100.0 ± 0.00 a | Normal mycelial growth | |
| Antimycin A | 0 | 0.0 ± 0.00 c | No germination |
| 6 | 42.1 ± 0.35 a | 26.7 ± 0.41% short germ tube and 15.4 ± 0.44% conidia lysed | |
| 12 | 26.7 ± 0.41 b | 12.6 ± 0.40% normal germ tube, 9.3 ± 0.68% short and 4.8 ± 0.29% Abnormally elongated germ tube | |
| 24 | 0.0 ± 0.00 c | No appressoria, no mycelial growth | |
| Nativo® 75 WG | 0 | 0.0 ± 0.00 b | No germination |
| 6 | 44.3 ± 2.33 a | Germinated with a short germ tube | |
| 12 | 44.3 ± 2.33 a | Normal germ tube | |
| 24 | 0.0 ± 0.00 b | No appressoria; no mycelial growth | |
| Parameter | Untreated Control | Healthy Control | Commercial Fungicide Nativo® 75 WG | Preventive (μg/mL) | Curative (μg/mL) | ||||
|---|---|---|---|---|---|---|---|---|---|
| 1 | 5 | 10 | 1 | 5 | 10 | ||||
| % disease incidence | 100 ± 0.00 a | 0 ± 0.00 d | 0 ± 0.00 d | 16.33 ± 2.19 b | 6.67 ± 0.88 c | 0.00 ± 0.00 d | 19 ± 1.15 b | 8.33 ± 0.67 c | 0.00 ± 0.00 d |
| % disease severity | 82 ± 4.73 a | 0 ± 0.00 c | 0 ± 0.00 c | 10.67 ± 2.96 b | 3.33 ± 0.88 b | 0 ± 0.00 c | 12.33 ± 2.40 b | 5.33 ± 1.20 bc | 0.00 ± 0.00 c |
| Treatment | Grain Yield Per Spike (gm) * | 1000-Grain Weight (gm) * | Disease Incidence (%) * | Disease Severity (%) * |
|---|---|---|---|---|
| Healthy control | 2.05 ± 0.05 a | 53.57 ± 1.37 a | 0.00 ± 0.00 c | 0.00 ± 0.00 c |
| Untreated control | 0.86 ± 0.04 b | 34.43 ± 0.27 c | 87.00 ± 2.91 a | 73.67 ± 2.65 a |
| Antimycin A | 1.95 ± 0.06 a | 42.73 ± 0.71 b | 33.00 ± 1.45 b | 23.67 ± 2.31 b |
| Nativo® 75 WG | 2.05 ± 0.13 a | 44.94 ± 1.55 b | 31.71 ± 2.96 b | 23.33 ± 2.95 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paul, S.K.; Chakraborty, M.; Rahman, M.; Gupta, D.R.; Mahmud, N.U.; Rahat, A.A.M.; Sarker, A.; Hannan, M.A.; Rahman, M.M.; Akanda, A.M.; et al. Marine Natural Product Antimycin A Suppresses Wheat Blast Disease Caused by Magnaporthe oryzae Triticum. J. Fungi 2022, 8, 618. https://doi.org/10.3390/jof8060618
Paul SK, Chakraborty M, Rahman M, Gupta DR, Mahmud NU, Rahat AAM, Sarker A, Hannan MA, Rahman MM, Akanda AM, et al. Marine Natural Product Antimycin A Suppresses Wheat Blast Disease Caused by Magnaporthe oryzae Triticum. Journal of Fungi. 2022; 8(6):618. https://doi.org/10.3390/jof8060618
Chicago/Turabian StylePaul, Sanjoy Kumar, Moutoshi Chakraborty, Mahfuzur Rahman, Dipali Rani Gupta, Nur Uddin Mahmud, Abdullah Al Mahbub Rahat, Aniruddha Sarker, Md. Abdul Hannan, Md. Mahbubur Rahman, Abdul Mannan Akanda, and et al. 2022. "Marine Natural Product Antimycin A Suppresses Wheat Blast Disease Caused by Magnaporthe oryzae Triticum" Journal of Fungi 8, no. 6: 618. https://doi.org/10.3390/jof8060618