
Monitoring Mycoparasitism of Clonostachys rosea against Botrytis cinerea Using GFP
by
 , ,
, ,
Rakibul Hasan
1,2 ,
,
Binna Lv
1,
Md. Jamal Uddin
1, 
Yingying Chen
1,
Lele Fan
1,
Zhanbin Sun
1,
Manhong Sun
1,* and
Shidong Li
1,* 1
Institute of Plant Protection, Chinese Academy of Agricultural Sciences, Beijing 100193, China
2
Department of Plant Pathology and Seed Science, Sylhet Agricultural University, Sylhet 3100, Bangladesh
*
Authors to whom correspondence should be addressed.
J. Fungi 2022, 8(6), 567; https://doi.org/10.3390/jof8060567
Submission received: 30 April 2022
/
Revised: 22 May 2022
/
Accepted: 23 May 2022
/
Published: 26 May 2022
(This article belongs to the Special Issue Interactions between Filamentous Fungal Pathogens and Hosts)
Abstract
:Clonostachys rosea is an important mycoparasite, with great potential for controlling numerous plant fungal diseases. Understanding the mechanisms and modes of action will assist the development and application of this biocontrol fungus. In this study, the highly efficient C. rosea 67-1 strain was marked with the green fluorescent protein (GFP), and the transformant possessed the same biological characteristics as the wild-type strain. Fungal interactions with Botrytis cinerea during co-culture and encounter on tomato leaves were assessed by fluorescence confocal and electron microscopy. The results indicated that once the two fungi met, the hyphae of C. rosea grew alongside those of B. cinerea, then attached tightly to the host and developed special structures, via which the biocontrol fungus penetrated the host and absorbed nutrients, eventually disintegrating the cells of the pathogen. Mycoparasitism to B. cinerea was also observed on tomato leaves, suggesting that C. rosea can colonize on plants and act following the invasion of the pathogenic fungus.
1. Introduction
Mycoparasitism is a common interaction in nature, in which a living fungus is parasitized by another fungus [1,2]. During the interaction, a mycoparasite perceives the presence of a potential host, grows towards and attaches to it, then often coils around it and forms infection structures that assist host penetration [3]. During the mycoparasitic process, various cell-wall-degrading enzymes (CWDEs) and antifungal metabolites are secreted to digest and kill host cells and uptake nutrients [4,5]. Mycoparasitism is an essential mechanism in the biocontrol fungi to fight against plant fungal diseases. Several mycoparasites have been studied for decades, among which Trichoderma spp. [6,7,8], Gliocladium spp. [9,10,11] and Coniothyrium minitans [12,13] are the most widely used biocontrol agents in the greenhouse and the field. However, unpredictable efficacy in the field sometimes hinders their application [14]. Understanding the interactions and the modes of action of the mycoparasites will promote their control effects to plant diseases and facilitate their application in the field.
To explore mycoparasitism, interactions between different fungal species are observed by microscopy [15,16,17]. Furthermore, molecular techniques involving constitutive expression of fluorescent proteins can be employed to visualize the associations of mycoparasites and hosts, and their colonization in soil and other non-sterile environments. [18]. In recent decades, the gene-encoding green fluorescent protein (GFP), originally isolated from the jellyfish Aequorea victoria [19], has been widely used as an effective molecular marker in many prokaryotes and eukaryotes without damaging cell activities. For example, GFP was expressed in Trichoderma species, which elucidated their interactions with Pythium ultimum, invasion of the hyphae and sclerotia of Rhizoctonia solani [4,20], and penetration of the plant parasitic nematode Globodera pallida [21]. Using a GFP marker, Németh [22] first visualized Ampelomyces quisqualis, a mycoparasite of powdery mildew, and deciphered its lifestyle in the environment, before and after acting as a mycoparasite.
Clonostachys rosea (syn. Gliocladium roseum) is an attractive mycoparasite capable of invading various phytopathogenic fungi, including R. solani, Sclerotinia sclerotiorum, Botrytis cinerea and Fusarium spp. [23,24,25]. Up to now, C. rosea preparations have been used in vegetables, ornamentals, and herbs to resist fungal diseases, and good results have been achieved in the greenhouse and the field [26,27,28]. Multiple biocontrol mechanisms have been reported for C. rosea, including antagonism, mycoparasitism, competition and induction of plant resistance [25,29], among which mycoparasitism plays an essential role during the biocontrol process. In previous studies, C. rosea isolates were successfully labelled with GFP, and their association with barley roots [30] and infection of the saprophytic nematode Panagrellus redivivus [31] were investigated. However, mycoparasitic interactions between C. rosea and pathogenic fungi remain unclear.
C. rosea 67-1 strain (namely C. chloroleuca 67-1) [32] is a highly efficient biocontrol fungus, targeting many plant pathogenic fungi [33,34]. In this work, a GFP-tagged 67-1 transformant was constructed, and its mycoparasitic activity against B. cinerea was investigated under in-vitro conditions and on tomato leaves. The research provides new insight into the mechanisms of mycoparasitism of C. rosea and will facilitate the development of the biocontrol fungus against plant fungal diseases.
2. Materials and Methods
2.1. Fungal Isolates and Plasmids
C. rosea 67-1 strain (ACCC 39160) was isolated from the soil of a vegetable yard in Hainan Province, China, by the sclerotia baiting method [35]. B. cinerea TC-B1 and S. sclerotium Ss-H were isolated from tomato plants with grey mould and Sclerotinia-infected soybean stems, respectively [36]. All isolates were cultured on potato dextrose agar (PDA) and preserved in 20% glycerol at −80 °C in the Biocontrol of Soil-borne Diseases Lab, Institute of Plant Protection (IPP), Chinese Academy of Agricultural Sciences (CAAS).
The recombinant plasmid pSC003 containing the GFP gene, glyceraldehyde-3-phosphate (gpd) promoter and trpC terminator was provided by Hao Zhang of the IPP and used to construct the GFP-labelled strain.
2.2. Plant Cultivars
The soybean cultivar Zigongdongdou No. 6 was provided by the Institute of Crop Sciences, CAAS, and the tomato cultivar Jiafen No. 201 was obtained from the Institute of Vegetables and Flowers, CAAS.
2.3. Construction of GFP-Labelled Strains of C. rosea
2.3.1. Protoplast Preparation and GFP Transformation
The 67-1 strain was cultured on PDA for 10 days. The spores were eluted, transferred into potato dextrose broth (PDB) and incubated in a fermentation shaker at 180 rpm at 28 °C overnight. The tine hyphae were treated with 40 mg/mL snail enzyme (XJK Biotech, Beijing, China) and incubated at 28 °C and 100 rpm for 3 h. The released protoplasts were collected with a sterilized double-layer microfiber cloth, suspended in STC buffer (200 g sucrose, 50 mL of 1 M Tris-HCl pH 8.0, 5.55 g CaCl2 in 1 L distilled water), and stored on ice [37].
PEG-CaCl2-mediated transformation of GFP was conducted [38]. The pSC003 plasmid was propagated in Escherichia coli DH5α (Transgen Biotech, Beijing, China) in Luria-Bertani (LB) medium with 100 μg/mL ampicillin, and the plasmid DNA was extracted using a TIANprep Rapid Mini Plasmid Kit (Transgen Biotech, Beijing, China). The protoplast suspension was mixed with the linearized plasmid and incubated on ice for 20 min, then 1.25 mL of 40% PTC solution (400 g polyethylene glycol 4000, 10 mL of 1 M Tris-HCl pH 8.0, 20 mg CaCl2 in 1 L distilled water) was added. After 20 min, the mixture was transferred into TB3 medium (3 g yeast extract, 3 g casein acid hydrolysate, 200 g sucrose in 1 L distilled water) and cultured at 28 °C and 100 rpm for 16 h.
2.3.2. Verification of GFP Transformants
The suspension of C. rosea was mixed with 10 mL TB3 medium containing 200 μg/mL G418 and 0.7% agarose and incubated at 26 °C for 2−3 days. The emerged colonies were picked and transferred onto the resistant PDA plates. After four generations, stable transformants were checked under an LSM 980 fluorescence confocal microscope (Carl Zeiss Microscopy GmbH, Jena, Germany). DNA was extracted from the transformants using a Bio Spin Fungus Genomic DNA Extraction Kit (Takara, Dalian, China). The primers GFPF 5′-GTGACCACCTTCACCTACGG-3′ and GFPR 5′-TGTACAGCTCGTCCATGCC-3′ were designed using Primer3Plus Software (Premier, Boston, MA, USA), and PCR amplification was performed in a 25 µL reaction system containing 23 µL PCR mix, 0.5 µL of each primer, and 1 µL DNA template. Thermal cycling was performed with an initial denaturation step at 98 °C for 3 min, followed by 35 cycles at 98 °C for 10 s, 56 °C for 10 s and 72 °C for 1 min, and a final elongation at 72 °C for 8 min.
2.3.3. Growth and Sporulation of the Transformants
The transformants exhibiting strong fluorescence were selected for bioassay. The isolates were incubated on PDA for 10 days, and the spore suspension of 1 × 107 spores/mL was prepared. Five microliters of the suspension were inoculated on a 5 mm sterilized filter paper on the centre of a 9 cm PDA plate. After incubation at 26 °C for 7 days, the diameter of each colony was measured. The spores were eluted with 5 mL of 0.05% Tween-80, and the spore yield was counted under a BX41 microscope (Olympus, Tokyo, Japan). Three replicates were conducted for each isolate.
2.3.4. Efficacy of GFP-Tagged Isolates on Soybean Sclerotinia Stem Rot
The control efficacies of the wild-type strain and GFP-tagged transformants against soybean Sclerotinia stem rot were tested in the greenhouse. Soil collected from a vegetable yard was mixed with 20% nursery substrate and filled in plastic pots (dia. 11 cm), in which three soybean seeds were sown. When six compound leaves had grown, 100 mL of C. rosea suspension (1 × 107 spores/mL) was sprayed onto them. After drying, an equal volume of S. sclerotiorum fermentation liquor was smeared with a brush. Seedlings treated with sterilized water and S. sclerotiorum broth served as the control. Twelve pots were planted for each isolate, and all pots were arranged randomly and maintained at 26−28 °C in the greenhouse. After 7 days, disease indices on all unfolded leaves were determined using a 0−4 grade scale according to lesions on the leaves [36]. Three replicates were conducted for each treatment.
2.4. Mycoparasitism of C. rosea to B. cinerea under In Vitro Conditions
A transformant with the same biological characteristics as the 67-1 strain was selected, named G67-1, and its mycoparasitic actions to B. cinerea were investigated on slides with and without nutrients. The G67-1 and TC-B1 strains were incubated on PDA at 26 °C for 7 days, and the agar strips (2 × 0.5 cm) of both isolates were cut from the edges of the fungal colonies with a sterilized scalpel. A sterilized glass slide (8 × 3 cm) was placed on the centre of a PDA plate, and a strip of C. rosea agar was laid on one side adjacent to the slide. The fungus was cultured at 26 °C for 2 days, and a strip of S. sclerotiorum was laid on the opposite side. In another run, 5 mL of melted PDA was poured into a 9 cm Petri dish, and a sterilized slide was gently submerged into the medium. After solidification, a thin layer of nutrients formed on the surface of the glass, and the two fungi were inoculated successively on both sides. The plates were sealed with Parafilm, and the fungi were co-cultured at 26 °C in an incubator. After 3−4 days, when the two fungi were observed to contact each other (0 h), the slides were taken out, and a droplet of sterile distilled water was added. Interactions between the fungi on the slides were observed under a fluorescence confocal microscope (Carl Zeiss Microscopy GmbH, Göttingen, Germany). A set of fluorescent filters were employed, including a dichroic mirror (495 nm), an excitation filter (450−490 nm) and a barrier filter (500−550 nm). Photographs were captured by an AxioCam ICc5 camera (Carl Zeiss Microscopy, LLC, Thornwood, NY, USA) and processed using Zen 2011 Software (Carl Zeiss Pte. Ltd., Singapore, Singapore). Minor photo editing was performed using Adobe Photoshop (Adobe Systems Incorporated, San Jose, CA, USA) without any changes to the content. The mycoparasitic process was monitored every 24 h, and 10 slides were assessed until the degradation of the host was observed.
2.5. Histological Observation by Scanning Electron Microscopy (SEM)
The confrontation of C. rosea and B. cinerea was also monitored by SEM. The agar strips (2 × 0.5 cm) were cut from the edges of C. rosea and B. cinerea colonies and placed 3 cm from each other on the surface of 5 mL PDA in a 9 cm Petri dish. The plates were sealed with Parafilm, and the fungi were co-cultured at 26 °C for 6 days until G67-1 strain overgrew B. cinerea colonies. The conjoint regions containing both fungi were cut into ~5 mm blocks, fixed in 3% glutaraldehyde at 25 °C in the dark for 48 h, and stored at 4 °C in a refrigerator before observation. Three replicates were included for each sample. The specimens were gently dried using a Leica EM CPD030 instrument (Leica Microsystems, Australia) and coated with gold powder. Interactions between the mycoparasite and its host were detected under a Hitachi SU8010 scanning electron microscope (Hitachi High-Technologies Co., Tokyo, Japan) with an accelerating voltage of 10 kV.
2.6. Interaction of C. rosea and B. cinerea in Tomato Plants
Infection of B. cinerea by C. rosea isolates was investigated on tomato leaves in the greenhouse. Soil collected from an experimental field of the Institute of Plant Protection, CAAS, Langfang, China, was mixed thoroughly with nursery substrate (4:1, v/v) and filled in 11 cm plastic pots. The tomato seeds were surface sterilized with 2.5% NaClO for 10 min, rinsed three times with distilled water, and sown in a nursery tray (54 × 28 cm, 10 × 5 holes). After 20 days, the seedlings were transplanted into the pots, two seedlings per pot. After growing for 7 days, the tomato leaves were inoculated with 100 mL of GFP-labelled C. rosea suspension at a concentration of 1 × 107 spores/mL and B. cinerea fermentation liquor successively. Relative humidity was maintained at 90%, and the temperature in the greenhouse was maintained at 26−28 °C.
Five days after inoculation, the leaves were picked and cut into 1 cm discs with a puncher. Mycoparasitism of C. rosea against B. cinerea on the surface of the leaflets was observed under a confocal scanning microscope (Carl Zeiss Microscopy GmbH, Jena, Germany). Simultaneously, the leaflets were surface sterilized with 2.5% NaClO for 3 min, rinsed with sterile distilled water, and transferred onto 1/4 PDA plates to examine infection of B. cinerea and colonization of C. rosea in tomato leaves. After 7 days, the fungal colonies were morphologically identified.
2.7. Statistical Analysis
Analysis of variance (ANOVA) was assessed using the Package agricolae v4.1.1 (R foundation for statistical computing, Vienna, Austria). The growth and sporulation of the fungal strains were compared by t-test, and their biocontrol activities were evaluated by Fisher’s least significant difference (LSD) test at the 5% significance level.
3. Results
3.1. Stability of GFP-Labelled Strains
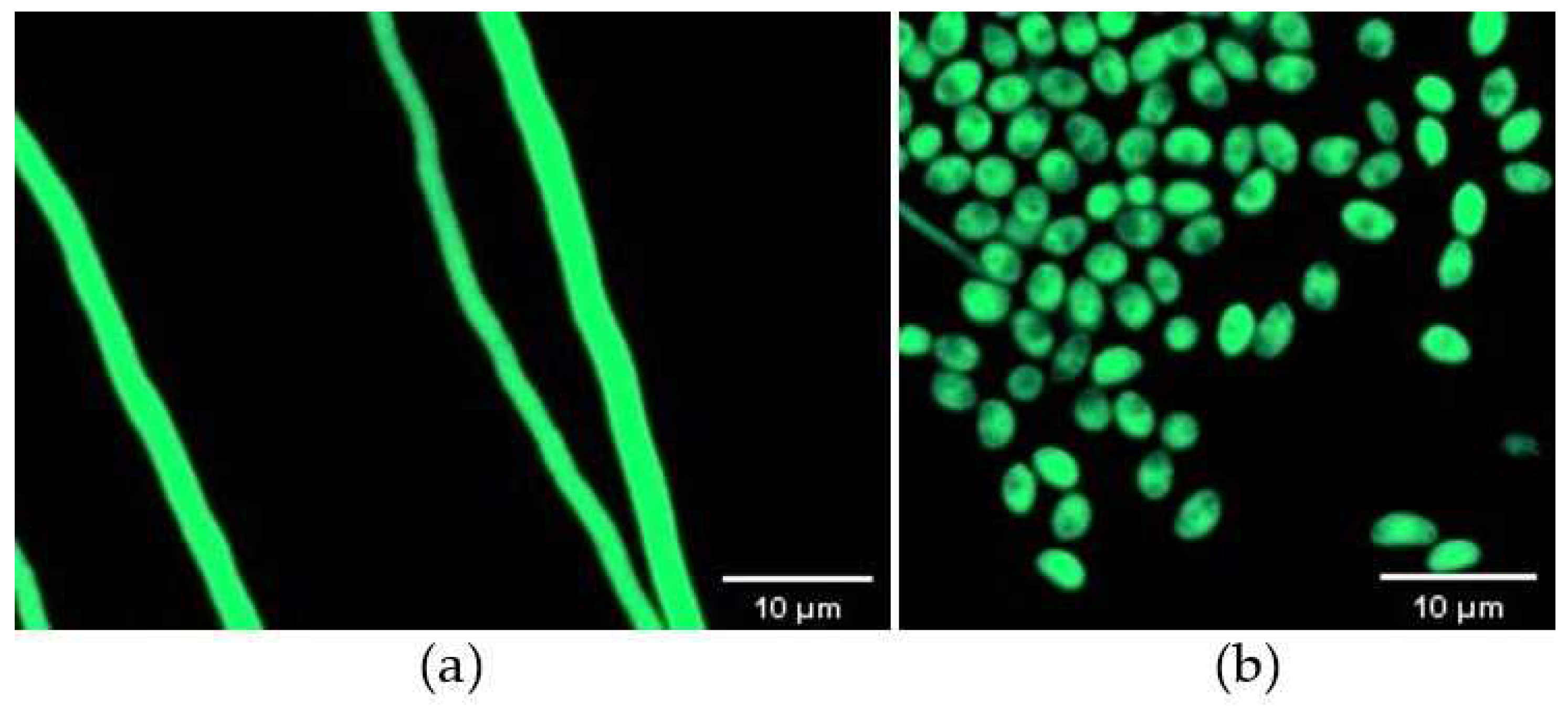
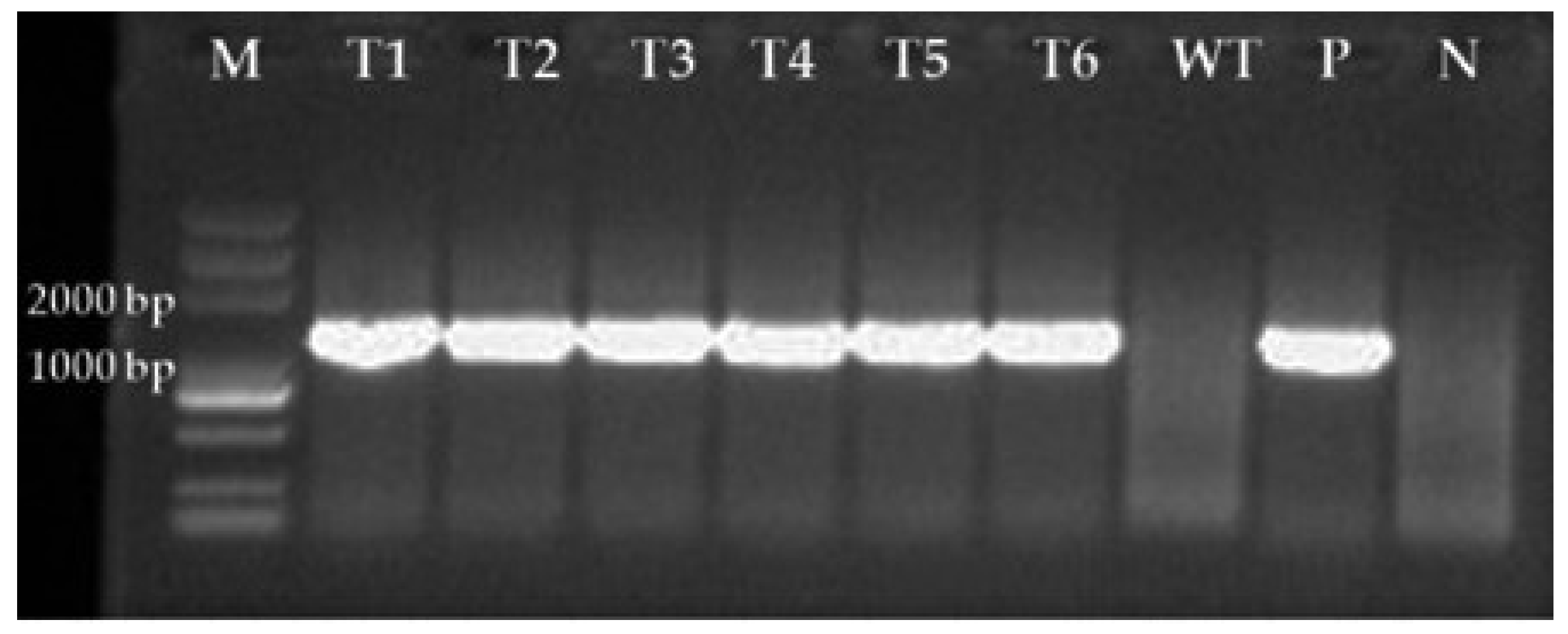
A total of 206 mutants grew on TB3 medium containing G418, and all 20 isolates tested emitted green fluorescence under a fluorescence microscope, among which 6 displayed strong excitation (Figure 1a,b). PCR verification showed that all tested mutants yielded a single band of approximately 1297 bp, indicating GFP was successfully inserted into the genome of the C. rosea 67-1 strain (Figure 2). The morphology of the transformants exhibiting the strongest fluorescence was quite similar to that of the wild-type strain (Figure 3), and their growth diameters and spore yields were 4.5 cm and 5.4 × 107 spores/plate after 7 days, compared with 4.7 cm and 5.7 × 107 spores/plate for the wild-type strain, which showed no statistical difference (p < 0.05). In the pot experiment, the G67-1 and 67-1 strains also showed consistent control effects against soybean Sclerotinia stem rot (p < 0.05, Table 1). The above results indicate that the GFP-labelled G67-1 strain can function as the 67-1 strain.
3.2. Interactions between C. rosea and B. cinerea under In Vitro Conditions
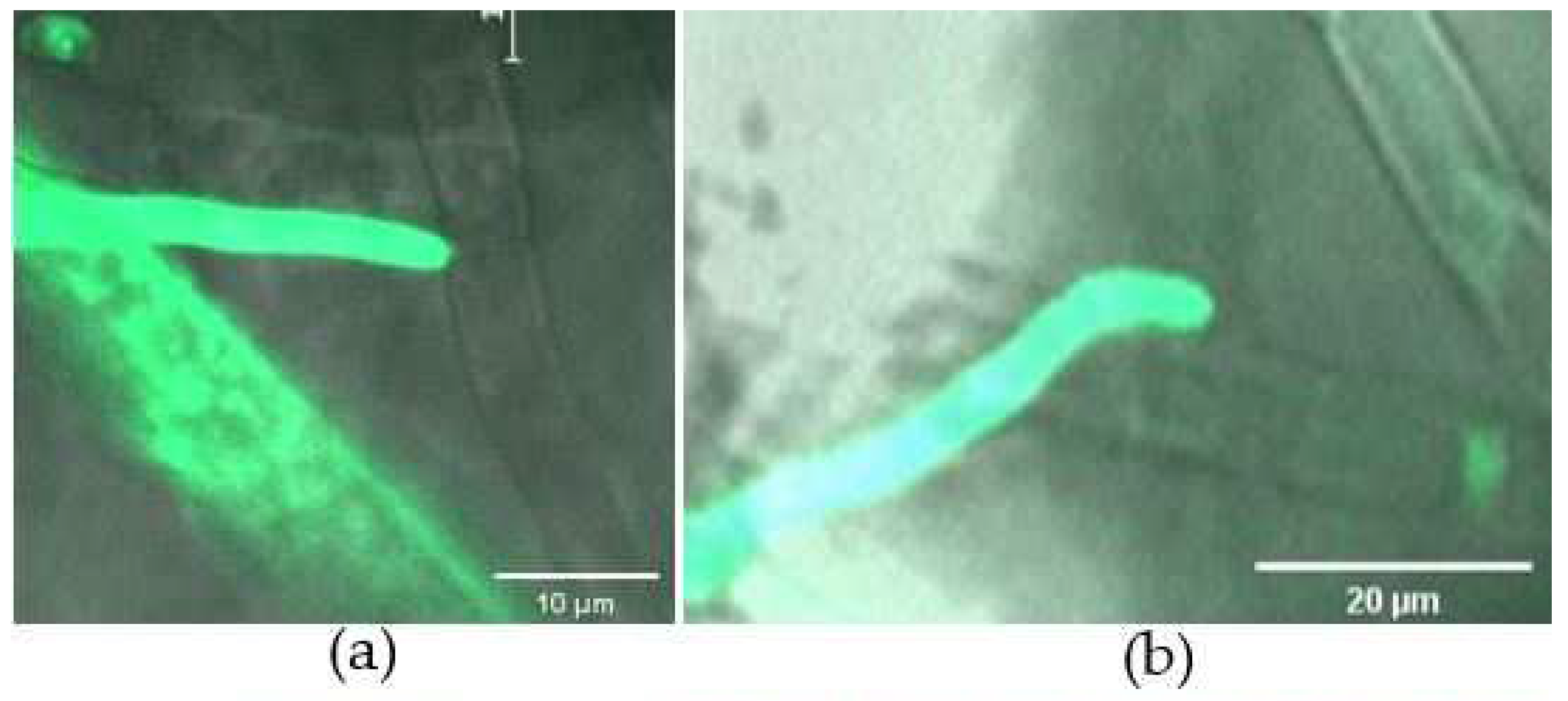
When co-cultured on the opposite sides of the plate, the hyphae of C. rosea and B. cinerea extended across the slide. Under a microscope, we could see that C. rosea grew towards its host over time (Figure 4a). After ~24 h, the hyphae of C. rosea attached to the host (Figure 4b), during which a swollen papilla-like structure formed on the tip of the hyphae (Figure 4c). The mycoparasite hyphae were tight against those of its host and grew alongside them (Figure 4d). On glass slides covering PDA medium, both fungi grew faster than those on slides without nutrients. However, an extra 12 h was needed to produce infection structures for mycoparasitizing the host, implying C. rosea might more easily utilize available nutrients. During the mycoparasitic process, the biocontrol fungus branched profusely, and it was clearly seen that C. rosea branches grew on and around the hyphae of its host (Figure 4e).
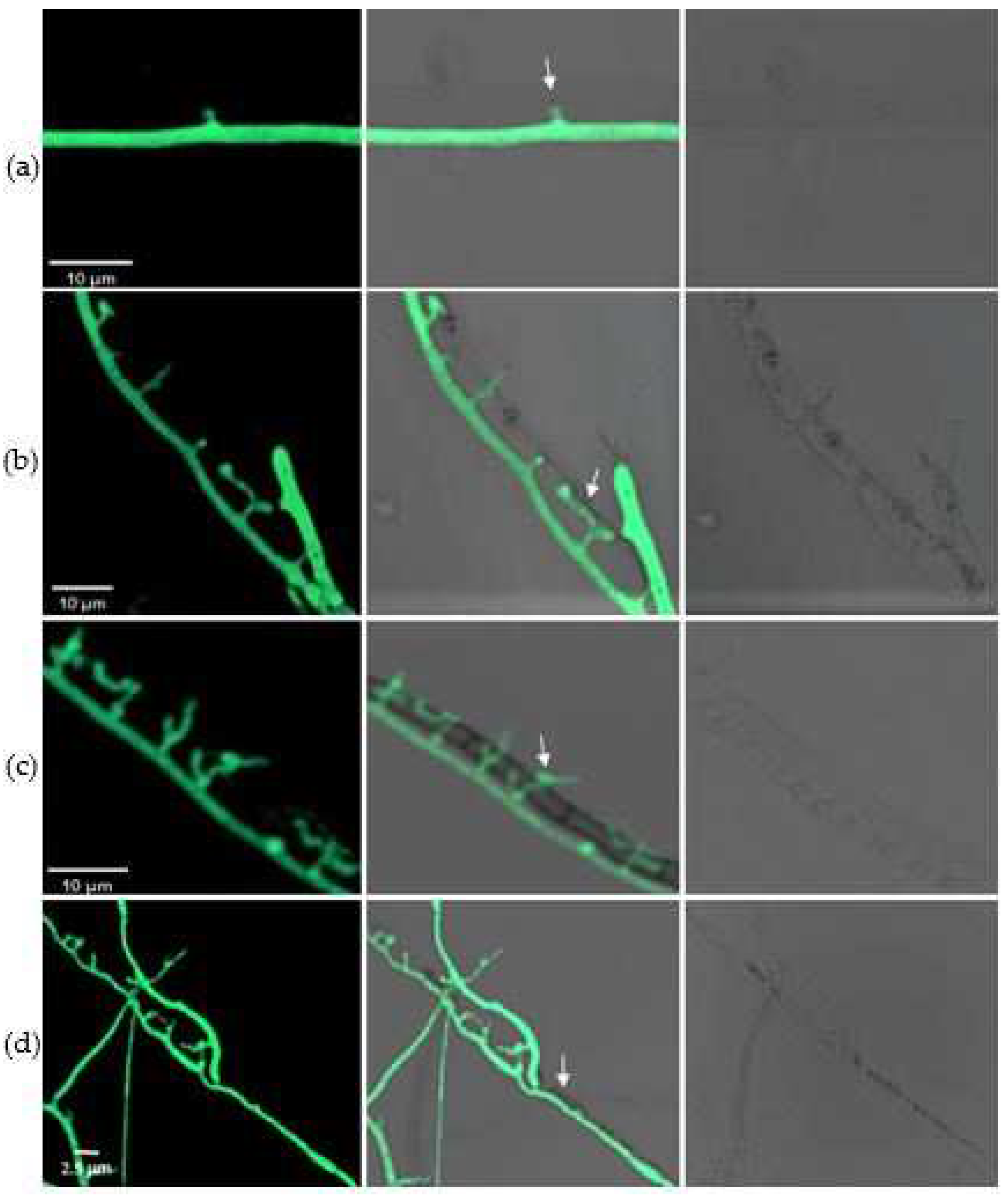
During the mycoparasitic interaction, some specific structures formed that were essential for penetrating the host. At 72 h after encounter, a unique hook-like structure was evident in the mycelia of C. rosea (Figure 5a). Appressoria then developed rapidly, and the invasion of the pathogen was achieved via these structures (Figure 5b). Once penetrating into the cells, the mycoparasite branched inside the host and used the pathogen as a nutrient source (Figure 5c). After 6 days, the hyphae of the pathogen gradually became damaged, and were eventually completely destroyed due to infection with the biocontrol fungus (Figure 5d). The penetration structures also developed in C. rosea on the nutrient-supplied slips, but this was dependent on the richness of the medium.
Under an SEM, it could be clearly seen that the hyphae of B. cinerea were tightly wrapped by those of the biocontrol fungus (Figure 6a), and an amount of appressoria generated from the hyphae of C. rosea, by which the mycoparasite penetrated the host cells (Figure 6b). At this point, the cell walls of the pathogen were partially degraded (Figure 6c). In addition to attacking fungal mycelia, C. rosea could also attack B. cinerea spores, and microscopy showed that the newly produced host conidia were penetrated by C. rosea appressoria (Figure 6d).
3.3. Colonization of C. rosea and Its Mycoparasitic Interaction with B. cinerea on Tomato Leaves
Three days after inoculation, the hyphae of B. cinerea and GFP-labelled C. rosea spread on the surface of the leaflets. When encountering the host, the hyphae of the biocontrol fungus grew alongside the mycelia of B. cinerea, and developed new branches extending towards the pathogen (Figure 7a,b). B. cinerea and C. rosea were also isolated from surface-sterilized tomato leaves, and identified by colony morphology and fluorescence microscopy (Figure 7c,d). The results showed that C. rosea could colonize on the surface and inside tomato leaves and attack the pathogen.
4. Discussion
Using GFP as a marker, interactions between different fungal species can be investigated [4,39,40]. Our current work showed that almost all the GFP transformants of C. rosea were stable and fluorescence could be detected in the hyphae and spores of the mutants continuously cultured in plates and in different infection stages, indicating that GFP had been successfully inserted into the genome of the C. rosea 67-1 strain. Six mutants with strong fluorescence were assayed, and all GFP-labelled transformants were quite similar to the wild-type strain in terms of hyphal extension, sporulation, and biocontrol activities, ensuring no major biological changes had occurred that might affect the interactions of the mycoparasite and its host.
In a mycoparasitic process, a host is penetrated by a predator and typically utilized as a food source. Understanding the interactions and modes of action of mycoparasites will promote their control effects to plant fungal diseases and facilitate their application in the field. In the present study, the mycoparasitism of C. rosea against B. cinerea was investigated, during which four stages were identified. Firstly, the biocontrol fungus grew towards its host. A mycoparasite may perceive compounds secreted by a host and initiate chemotactic responses to these signals [41,42]. When confronted with each other, C. rosea mycelia extended towards those of B. cinerea and produced many papilla-like outgrowths at the tips of the hyphae, where were active developing regions and sensitive to various stimuli. This kind of infection mechanism was also observed in T. atroviride when parasitizing R. solani and P. ultimum [4], and the exudates of R. solani could induce the formation of papilla in T. virens [43].
Next, the mycoparasite attached to its host. Under a fluorescence microscope, it was noticed that C. rosea hyphae attached to those of B. cinerea after 3 days of face-to-face cultivation, grew clinging to the pathogen, and stretched many branches to be ready to attack the host. In a previous study of T. harzianum, the hyphae were also found to specifically evolve and profusely branch to a powdery mildew species Oidium longipes [44]. During this time, the pathogens may secrete glutinous substances that are essential for the parasites to recognize their hosts. It has been evidenced that the interactions of lectin carbohydrate may mediate the recognition and attachment of Trichoderma species to soil-borne plant pathogenic fungi [45].
After attachment, C. rosea isolates generated specialized structures that penetrate host cells, but such structures are not always present in mycoparasites. No penetration structures were detected in Sphaerodes quadrangularis, a facultative biotrophic fungus, when co-cultured with F. oxysporum and F. graminearum [46]. During the mycoparasitic process, C. rosea generated a hook-like structure and appressoria to attack and penetrate the hyphae and spores of B. cinerea. Finally, the mycoparasite absorbed nutrients inside the host, and the cells of the pathogen were gradually disintegrated.
It is also essential for biocontrol fungi to secrete chemical compounds to accomplish host invasion. During the interaction, the mycoparasites may produce multiple cell wall degrading enzymes (CWDEs) and secondary metabolites, such as antibiotics and toxins, that accelerate the degradation of the hosts [9,47]. Several enzymes, including chitinase, cellulase and glucanase, were identified in Trichoderma isolates when colonizing their hosts [5,48]. In the present study, B. cinerea cell walls were damaged when invaded, implying that some hydrolases might be involved in the mycoparasitism of C. rosea.
Many mycoparasite species are facultative fungi that can acquire nutrition from various ambient environments, including soil, plant residues, culture media and fungal hosts. Competition for limited nutrients is essential for the survival of microorganisms, and it is considered as one of the important mechanisms for the biological control of plant fungal pathogens [49]. When placed on the opposite sides of slides without any nutrients, the biocontrol fungus attacked its host and developed dedicated infection structures (papilla, coil, hook and appressoria) within 72 h, while on slides containing PDA medium, it was only seen that C. rosea hyphae attached and grew parallel to host hyphae after 72 h, suggesting that C. rosea might preferentially utilize available nutrients, such as glucose, which delays colonization of the host. Similar results were reported in previous studies, which indicates that the mycoparasites are more likely to form appressoria to take up nutrients if external conditions are poor [50,51] and their biocontrol activities become more effective [52,53].
Biotrophic mycoparasites obtain nutrition from living fungi without killing them. With the penetration structures or absorptive cells, the parasites build close relationships with their hosts, maintaining their survival and propagation [54]. However, for necrotrophic mycoparasites, when encountering potential preys, they may produce toxic compounds and degrading enzymes to kill the hosts and facilitate their invasion. Once colonization is established, the parasites take up nutrients from the dead cells and proliferate in the hosts. In general, most mycoparasitic species are necrotrophs, but they may have short biotrophic phases [55,56,57]. In our current study, it was seen that the C. rosea hyphae grew close to those of the host, produced appressorium-like structures by which they penetrated the mycelia and spores of B. cinerea, and decomposed the hyphae of the pathogen when the necrotrophic phase was initiated. In the area in which T. atroviride and R. solani interacted, the swelling of the hyphae of the biocontrol fungus was observed [4]. Further, when some Trichoderma species attach to prey, it is often followed by coiling around host hyphae and forming appressorium-like structures that assist penetration [58]. However, obligate biotrophic mycoparasites, such as Verticillium biguttatum, Gliocephalis hyaline and Olpitrichum tenellum, can only survive on living organisms and cannot grow on culture media [54,59,60]. Our results show that C. rosea can produce penetration structures and propagate in pathogenic fungi, and can also colonize on plants and within plant tissues, suggesting that C. rosea is a promising facultative mycoparasite with great advantages for targeting plant fungal diseases whenever pathogens invade.
5. Conclusions
In this study, a highly efficient C. rosea 67-1 strain was tagged with GFP, and its interaction with B. cinerea was investigated by fluorescence confocal microscopy and SEM. When encountering the host, C. rosea produces dedicated infection structures, such as papilla, coil, and appressoria, to attack and penetrate the pathogen, absorbs nutrients and disintegrates the pathogen cells. C. rosea can also colonize on tomato leaves and thereby, protect the plants from infection. To the best of our knowledge, this is the first report of infection structures formed in C. rosea during the mycoparasitic process. The findings are of great significance to reveal the mechanisms of the biocontrol fungus and promote the development and application of C. rosea biocontrol agents.
Author Contributions
Conceived and designed the study, M.S. and S.L.; performed experiments, R.H., Y.C. and L.F.; data analysis, R.H. and Z.S.; manuscript preparation, R.H., B.L. and M.J.U.; reviewed the manuscript, M.S.; funding acquisition, S.L. and M.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the National Key Research and Development Program of China (2019YFD1002000), the Science and Technology Program of Qinghai (2019-NK-116), and China Agriculture Research System of MOF and MARA (CARS-23-C04).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Acknowledgments
The authors are grateful to H. Zhang at the Institute of Plant Protection, CAAS, for kindly providing the plasmid.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Viterbo, A.; Horwitz, B. Mycoparasitism. In Cellular and Molecular Biology of Filamentous Fungi; Borkovich, K., Ebbole, D.J., Eds.; ASM: Washington, DC, USA, 2010; pp. 676–693. [Google Scholar] [CrossRef]
- Boddy, L. Interactions between Fungi and Other Microbes. In The Fungi, 3rd ed.; Watkinson, S.C., Boddy, L., Money, N., Eds.; Elsevier: Amsterdam, The Netherlands, 2016; pp. 337–360. [Google Scholar] [CrossRef]
- Karlsson, M.; Atanasova, L.; Jensen, D.F.; Zeilinger, S. Necrotrophic mycoparasites and their genomes. In The Fungal Kingdom; Heitman, J., Howlett, J.B., Crous, P.W., Stukenbrock, E.H., James, T.Y., Gow, N.A.R., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 1005–1026. [Google Scholar] [CrossRef]
- Lu, Z.; Tombolini, R.; Woo, S.; Zeilinger, S.; Lorito, M.; Jansson, J.K. In vivo study of Trichoderma pathogen plant interactions using constitutive and inducible green fluorescent protein reporter systems. Appl. Environ. Microb. 2004, 70, 3073–3081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almeida, F.B.; Cerqueira, F.M.; Silva Rdo, N.; Ulhoa, C.J.; Lima, A.L. Mycoparasitism studies of Trichoderma harzianum strains against Rhizoctonia solani: Evaluation of coiling and hydrolytic enzyme production. Biotechnol. Lett. 2007, 29, 1189–1193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, M.; Ashraf, S. Role of Trichoderma spp. as a biocontrol agent of fungal plant pathogens. In Probiotics and Plant Health; Kumar, V., Kumar, M., Sharma, S., Prasad, R., Eds.; Springer: Singapore, 2017; pp. 497–506. [Google Scholar] [CrossRef]
- Guzmán-Guzmán, P.; Porras-Troncoso, M.D.; Olmedo-Monfil, V.; Herrera-Estrella, A. Trichoderma species: Versatile plant symbionts. Phytopathology 2019, 109, 6–16. [Google Scholar] [CrossRef] [Green Version]
- Woo, S.L.; Scala, F.; Ruocco, M.; Lorito, M. The molecular biology of the interactions between Trichoderma spp., phytopathogenic fungi, and plants. Phytopathology 2006, 96, 181–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karlsson, M.; Durling, M.B.; Choi, J.; Kosawang, C.; Lackner, G.; Tzelepis, G.D.; Nygren, K.; Dubey, M.K.; Kamou, N.; Levasseur, A.; et al. Insights on the evolution of mycoparasitism from the genome of Clonostachys rosea. Genome Biol. Evol. 2015, 7, 465–480. [Google Scholar] [CrossRef]
- Lysøe, E.; Dees, M.W.; Brurberg, M.B. A three-way transcriptomic interaction study of a biocontrol agent (Clonostachys rosea), a fungal pathogen (Helminthosporium solani), and a potato host (Solanum tuberosum). Mol. Plant Microbe Interact. 2017, 30, 645–655. [Google Scholar] [CrossRef] [Green Version]
- Meng, F.; Lv, R.; Cheng, M.; Mo, F.; Zhang, N.; Qi, H.; Liu, J.; Chen, X.; Liu, Y.; Ghanizadeh, H.; et al. Insights into the molecular basis of biocontrol of Botrytis cinerea by Clonostachys rosea in tomato. Sci. Hortic. 2022, 291, 110547. [Google Scholar] [CrossRef]
- Zeng, W.; Wang, D.; Kirk, W.; Hao, J. Use of Coniothyrium minitans and other microorganisms for reducing Sclerotinia sclerotiorum. Biol. Control 2012, 60, 225–232. [Google Scholar] [CrossRef]
- Zhao, H.; Zhou, T.; Xie, J.; Cheng, J.; Jiang, D.; Fu, Y. Host transcriptional response of Sclerotinia sclerotiorum induced by the mycoparasite Coniothyrium minitans. Front. Microbiol. 2020, 11, 183. [Google Scholar] [CrossRef]
- Abd-Elgawad, M.M.M.; Askary, T.H. Factors affecting the success of biological agents used in controlling the plant-parasitic nematodes. Egypt. J. Biol. Pest Control. 2020, 30, 17. [Google Scholar] [CrossRef]
- Smith, S.N.; Prince, M.; Whipps, J.M. Characterization of Sclerotinia and mycoparasite Coniothyrium minitans interaction by microscale co-culture. Lett. Appl. Microbiol. 2008, 47, 128–133. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Vujanovic, V. Changes in mycoparasite Fusarium host’s interfaces in response to the hostile environment as revealed by water contact angle and atomic force microscopy. Biol. Control 2018, 121, 247–255. [Google Scholar] [CrossRef]
- Vujanovic, V.; Kim, S.H.; Lahlali, R.; Karunakaran, C. Spectroscopy and SEM imaging reveal endosymbiont dependent components changes in germinating kernel through direct and indirect coleorhiza fungus interactions under stress. Sci. Rep. 2019, 9, 1665. [Google Scholar] [CrossRef]
- Errampalli, D.; Leung, K.; Cassidy, M.B.; Kostrzynska, M.; Blears, M.; Lee, H.; Trevors, J.T. Applications of the green fluorescent protein as a molecular marker in environmental microorganisms. J. Microbiol. Meth. 1999, 35, 187–199. [Google Scholar] [CrossRef]
- Chalfie, M.; Tu, Y.; Euskirchen, G.; Ward, W.W.; Prasher, D.C. Green fluorescent protein as a marker for gene expression. Science 1994, 263, 802–805. [Google Scholar] [CrossRef] [Green Version]
- Sarrocco, S.; Mikkelsen, L.; Vergara, M.; Jensen, D.F.; Lübeck, M.; Vannacci, G. Histopathological studies of sclerotia of phytopathogenic fungi parasitized by a GFP transformed Trichoderma virens antagonistic strain. Mycol. Res. 2006, 110, 179–187. [Google Scholar] [CrossRef]
- Contina, J.B.; Dandurand, L.M.; Knudsen, G.R. Use of GFP-tagged Trichoderma harzianum as a tool to study the biological control of the potato cyst nematode Globodera pallida. Appl. Soil Ecol. 2017, 115, 31–37. [Google Scholar] [CrossRef] [Green Version]
- Németh, M.Z.; Pintye, A.; Horváth, A.N.; Vági, P.; Kovács, G.M.; Gorfer, M.; Kiss, L. GFP transformation sheds more light on a widespread mycoparasitic interaction. bioRxiv 2019, 516781. [Google Scholar] [CrossRef] [Green Version]
- Barnett, H.L.; Lilly, V.G. A destructive mycoparasite, Gliocladium roseum. Mycologia 2018, 54, 72–77. [Google Scholar] [CrossRef]
- Luongo, L.; Galli, M.; Corazza, L.; Meekes, E.; De Haas, L.; Van Der Plas, C.L.; Köhl, J. Potential of fungal antagonists for biocontrol of Fusarium spp. in wheat and maize through competition in crop debris. Biocontrol Sci. Technol. 2005, 15, 229–242. [Google Scholar] [CrossRef]
- Rodriguez, M.A.; Cabrera, G.; Gozzo, F.C.; Eberlin, M.N.; Godeas, A. Clonostachys rosea BAFC3874 as a Sclerotinia sclerotiorum antagonist: Mechanisms involved and potential as a biocontrol agent. J. Appl. Microbiol. 2011, 110, 1177–1186. [Google Scholar] [CrossRef] [PubMed]
- Jensen, B.; Knudsen, I.M.B.; Jensen, D.F. Biological seed treatment of cereals with fresh and long-term stored formulations of Gliocladium roseum: Biocontrol efficacy against Fusarium culmorum. Eur. J. Plant Pathol. 2000, 106, 233–242. [Google Scholar] [CrossRef]
- Wu, H.Q.; Sun, L.L.; Liu, F.; Wang, Z.Y.; Cao, C.W. Preparation of dry flowable formulations of Clonostachys rosea by spray drying and application for Sclerotinia sclerotiorum control. J. Integr. Agric. 2018, 17, 613–620. [Google Scholar] [CrossRef] [Green Version]
- Tian, T.; Li, S.D.; Sun, M.H. Synergistic effect of dazomet soil fumigation and Clonostachys rosea against cucumber Fusarium wilt. Phytopathology 2014, 104, 1314–1321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Köhl, J.; Kolnaar, R.; Ravensberg, W.J. Mode of action of microbial biological control agents against plant diseases: Relevance beyond efficacy. Front. Plant Sci. 2019, 10, 845. [Google Scholar] [CrossRef] [Green Version]
- Lübeck, M.; Knudsen, I.M.B.; Jensen, B.; Thrane, U.; Janvier, C.; Jensen, D.F. GUS and GFP transformation of the biocontrol strain Clonostachys rosea IK726 and the use of these marker genes in ecological studies. Mycol. Res. 2002, 106, 815–826. [Google Scholar] [CrossRef]
- Zhang, L.; Yang, J.; Niu, Q.; Zhao, X.; Ye, F.; Liang, L.; Zhang, K.Q. Investigation on the infection mechanism of the fungus Clonostachys rosea against nematodes using the green fluorescent protein. Appl. Microbiol. Biot. 2008, 78, 983–990. [Google Scholar] [CrossRef]
- Moreira, G.M.; Abreu, L.M.; Carvalho, V.G.; Schroers, H.J.; Pfenning, L.H. Multilocus phylogeny of Clonostachys subgenus Bionectria from Brazil and description of Clonostachys chloroleuca sp. nov. Mycol. Prog. 2016, 15, 1031–1039. [Google Scholar] [CrossRef]
- Zhang, Y.H.; Li, S.D.; Wang, G.Q.; Otsu, Y.; Wang, Z.Y. Biological control of soybean Sclerotinia rot with wettable powder of Gliocladium. Plant Prot. 2007, 33, 141–142. [Google Scholar]
- Ma, G.Z.; Wang, S.F.; Bao, Z.H.; Li, S.D.; Sun, M.H.; Fu, H.R.; Ge, P.H. Inhibition and protective activities of Gliocladium roseum 67-1 to Rhizoctonia solani. Crops 2011, 6, 77–80. [Google Scholar]
- Zhang, Y.H.; Gao, H.L.; Ma, G.Z.; Li, S.D. Mycoparasitism of Gliocladium roseum 67-1 on Sclerotinia sclerotiorum. Acta Phytopathol. Sin. 2004, 34, 211–214. [Google Scholar]
- Sun, Z.B.; Sun, M.H.; Li, S.D. Identification of mycoparasitism related genes in Clonostachys rosea 67-1 active against Sclerotinia sclerotiorum. Sci. Rep. 2015, 5, 18169. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.B.; Sun, M.H.; Zhou, M.; Li, S.D. Transformation of the endochitinase gene Chi67-1 in Clonostachys rosea 67-1 increases its biocontrol activity against Sclerotinia sclerotiorum. AMB Express 2017, 7, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, M.Q.; Jiang, J.; Sun, M.H.; Xie, X.M.; Li, S.D. Construction of Clonostachys rosea 67-1 genetic transformation system by restriction enzyme-mediated integration (REMI). Chin. J. Biol. Control 2013, 29, 263–269. [Google Scholar]
- Lorito, M.; Woo, S.L.; Garcia, I.; Colucci, G.; Harman, G.E.; Pintor-Toro, J.A.; Filippone, E.; Muccifora, S.; Lawrence, C.B.; Zoina, A.; et al. Genes from mycoparasitic fungi as a source for improving plant resistance to fungal pathogens. Proc. Natl. Acad. Sci. USA 1998, 95, 7860–7865. [Google Scholar] [CrossRef] [Green Version]
- Kowsari, M.; Motallebi, M.; Zamani, M. Construction of new GFP-tagged fusants for Trichoderma harzianum with enhanced biocontrol activity. J. Plant Prot. Res. 2014, 54, 122–131. [Google Scholar] [CrossRef] [Green Version]
- Moreno-Ruiz, D.; Lichius, A.; Turrà, D.; Di Pietro, A.; Zeilinger, S. Chemotropism assays for plant symbiosis and mycoparasitism related compound screening in Trichoderma atroviride. Front. Microbiol. 2020, 11, 601251. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Ge, H.; Zhang, F.; Guo, N.; Wang, Y.; Chen, L.; Ji, X.; Li, C. Biocontrol potential of Trichoderma harzianum isolate T-aloe against Sclerotinia sclerotiorum in soybean. Plant Physiol. Bioch. 2016, 100, 64–74. [Google Scholar] [CrossRef]
- Inayati, A.; Sulistyowati, L.; Aini, L.Q.; Yusnawan, E. Mycoparasitic activity of indigenous Trichoderma virens strains against mungbean soil-borne pathogen Rhizoctonia solani: Hyperparasite and hydrolytic enzyme production. J. Agric. Sci. 2020, 42, 229–242. [Google Scholar] [CrossRef]
- Kiss, L.; Pintye, A.; Zseli, G.; Jankovics, T.; Szentiványi, O.; Hafez, Y.M.; Cook, R.T.A. Microcyclic conidiogenesis in powdery mildews and its association with intracellular parasitism by Ampelomyces. Eur. J. Plant Pathol. 2010, 126, 445–451. [Google Scholar] [CrossRef]
- Inbar, J.; Chet, I. The role of lectins in recognition and adhesion of the mycoparasitic fungus Trichoderma spp. to its host. In Toward Anti-Adhesion Therapy for Microbial Diseases, Advances in Experimental Medicine and Biology; Kahane, I., Ofek, I., Eds.; Springer: Boston, MA, USA, 1996; Volume 408, pp. 229–231. [Google Scholar] [CrossRef]
- Goh, Y.K.; Vujanovic, V. Sphaerodes quadrangularis biotrophic mycoparasitism on Fusarium avenaceum. Mycologia 2010, 102, 757–762. [Google Scholar] [CrossRef] [PubMed]
- Sharma, K.; Mishra, A.K.; Misra, R.S. Morphological, biochemical and molecular characterization of Trichoderma harzianum isolates for their efficacy as biocontrol agents. J. Phytopathol. 2009, 157, 51–56. [Google Scholar] [CrossRef]
- de Lima, F.B.; Félix, C.; Osório, N.; Alves, A.; Vitorino, R.; Domingues, P.; da Silva Ribeiro, R.T.; Esteves, A.C. Trichoderma harzianum T1A constitutively secretes proteins involved in the biological control of Guignardia citricarpa. Biol. Control 2017, 106, 99–109. [Google Scholar] [CrossRef]
- Chet, I.; Inbar, J.; Hadar, I. Fungal antagonists and mycoparasites. In The Mycota IV: Environmental and Microbial Relationships, 2nd ed.; Wicklow, D., Söderström, B., Eds.; Springer: Berlin/Heidelberg, Germany, 1997; pp. 165–184. [Google Scholar] [CrossRef]
- Johnson, J.M.; Alex, T.; Oelmüller, R. Piriformospora indica: The versatile and multifunctional root endophytic fungus for enhanced yield and tolerance to biotic and abiotic stress in crop plants. J. Trop. Agric. 2014, 52, 103–122. [Google Scholar]
- Rajani, P.; Rajasekaran, C.; Vasanthakumari, M.M.; Olsson, S.B.; Ravikanth, G.; Shaanker, R.U. Inhibition of plant pathogenic fungi by endophytic Trichoderma spp. through mycoparasitism and volatile organic compounds. Microbiol. Res. 2020, 242, 126595. [Google Scholar] [CrossRef] [PubMed]
- Tjamos, E.C.; Papavizas, G.C.; Cook, R.J. Biological Control of Plant Diseases: Progress and Challenges for the Future; Springer: Berlin/Heidelberg, Germany, 2013; pp. 417–427. [Google Scholar]
- Benítez, T.; Rincón, A.M.; Limón, M.C.; Codón, A.C. Biocontrol mechanisms of Trichoderma strains. Int. Microbiol. 2004, 7, 249–260. [Google Scholar] [PubMed]
- van den Boogert, P.H.J.F.; Deacon, J.W. Biotrophic mycoparasitism by Verticillium biguttatum on Rhizoctonia solani. Eur. J. Plant Pathol. 1994, 100, 137–156. [Google Scholar] [CrossRef]
- Kosawang, C.; Karlsson, M.; Vélëz, H.; Rasmussen, P.H.; Collinge, D.B.; Jensen, B.; Jensen, D.F. Zearalenone detoxification by zearalenone hydrolase is important for the antagonistic ability of Clonostachys rosea against mycotoxigenic Fusarium graminearum. Fungal Biol. 2014, 118, 364–373. [Google Scholar] [CrossRef]
- Schöneberg, A.; Musa, T.; Voegele, R.T.; Vogelgsang, S. The potential of antagonistic fungi for control of Fusarium graminearum and Fusarium crookwellense varies depending on the experimental approach. J. Appl. Microbiol. 2015, 118, 1165–1179. [Google Scholar] [CrossRef]
- Jensen, B.; Lübeck, P.S.; Jørgensen, H.J.L. Clonostachys rosea reduces spot blotch in barley by inhibiting pre-penetration growth and sporulation of Bipolaris sorokiniana without inducing resistance. Pest Manag. Sci. 2016, 72, 2231–2239. [Google Scholar] [CrossRef]
- Chet, I.; Harman, G.E.; Baker, R. Trichoderma hamatum: Its hyphal interactions with Rhizoctonia solani and Pythium spp. Microb. Ecol. 1981, 7, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, K.; Holtzman, K.; Seifert, K.A. Morphology, phylogeny and biology of Gliocephalis hyalina, a biotrophic contact mycoparasite of Fusarium species. Mycologia 2005, 97, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Li, D.C.; Shen, C.Y. Olpitrichum tenellum as a biotrophic contact mycoparasite. Can. J. Bot. 1996, 74, 2014–2016. [Google Scholar] [CrossRef]
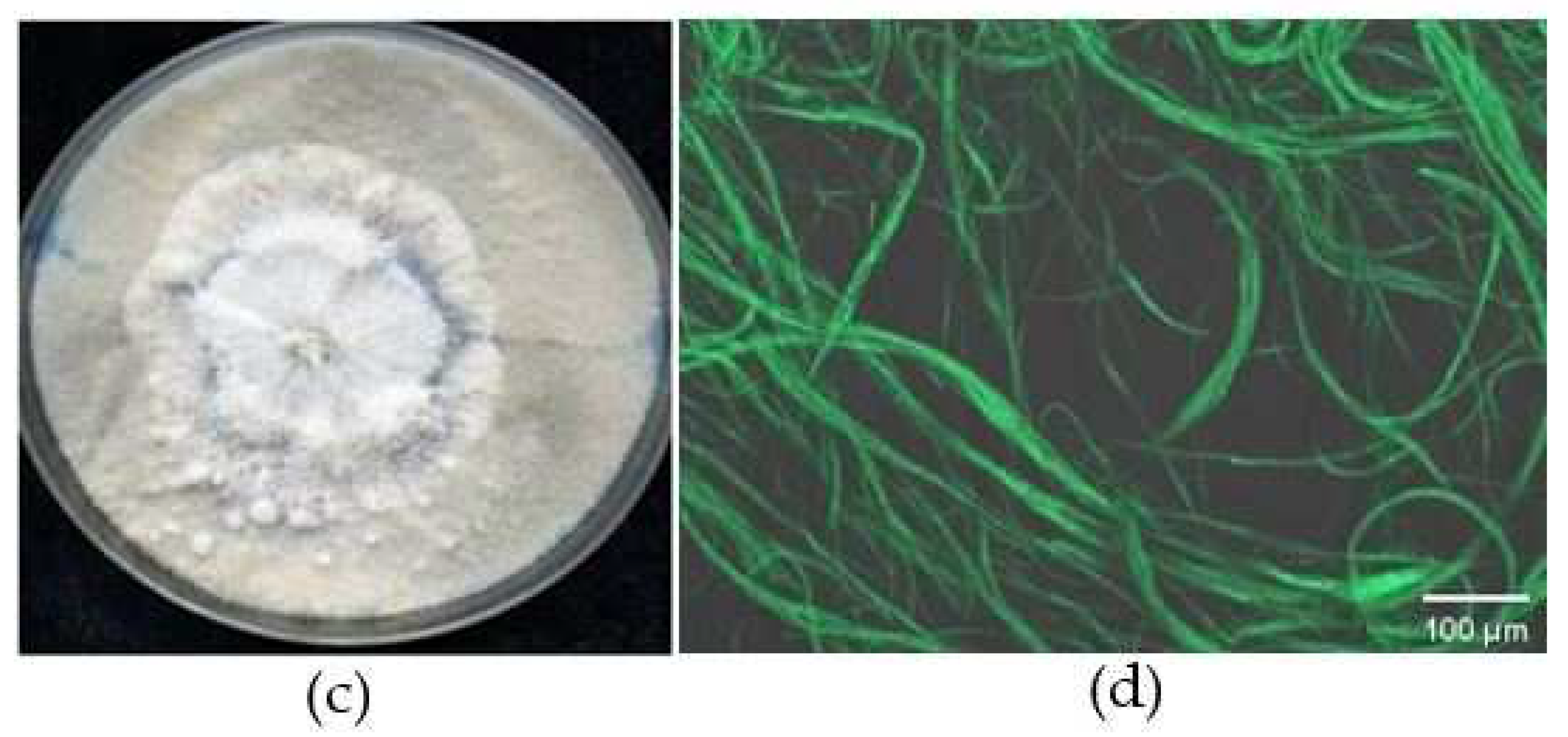
Figure 1.
Confocal microscopy images of GFP-labelled C. rosea 67-1 transformants observed under phase-contrast light. (a) Vegetative hyphae. (b) Conidia.
Figure 1.
Confocal microscopy images of GFP-labelled C. rosea 67-1 transformants observed under phase-contrast light. (a) Vegetative hyphae. (b) Conidia.

Figure 2.
PCR verification of GFP-labelled C. rosea transformants using primers GFPF and GFPR. M, DNA markers; T1−T6, Transformants; WT, Wild-type strain; P, Plasmid; N, Negative control.
Figure 2.
PCR verification of GFP-labelled C. rosea transformants using primers GFPF and GFPR. M, DNA markers; T1−T6, Transformants; WT, Wild-type strain; P, Plasmid; N, Negative control.

Figure 3.
Colony morphology of GFP–labelled C. rosea 67-1 transformant on PDA plate after 7 days. WT, Wild-type strain.
Figure 3.
Colony morphology of GFP–labelled C. rosea 67-1 transformant on PDA plate after 7 days. WT, Wild-type strain.

Figure 4.
Interaction between C. rosea G67-1 and B. cinerea TC-B1 strains confrontation cultured on slides within 0−48 h. (a) Hyphal extension of G67-1 towards the mycelia of TC-B1. (b) Attachment of G67-1 hyphae to those of its host. (c) Papilla formed at the tip of G67-1 hyphae. (d) G67-1 hyphae growing alongside its host. (e) Hyphal branches of G67-1 growing on and around its host.
Figure 4.
Interaction between C. rosea G67-1 and B. cinerea TC-B1 strains confrontation cultured on slides within 0−48 h. (a) Hyphal extension of G67-1 towards the mycelia of TC-B1. (b) Attachment of G67-1 hyphae to those of its host. (c) Papilla formed at the tip of G67-1 hyphae. (d) G67-1 hyphae growing alongside its host. (e) Hyphal branches of G67-1 growing on and around its host.

Figure 5.
Infection structures formed in C. rosea G67-1 strain against B. cinerea. (a) Hook-like structures of G67-1 formed at 72 h after encountering B. cinerea. (b) Production of appressoria and penetration into host hyphae. (c) Appressorium branches produced inside host cells. (d) Disintegration of B. cinerea after 6 days. Mycoparasitic interaction between the two isolates was observed using a fluorescence microscope under phase-contrast, differential interference contrast (DIC), and normal fields.
Figure 5.
Infection structures formed in C. rosea G67-1 strain against B. cinerea. (a) Hook-like structures of G67-1 formed at 72 h after encountering B. cinerea. (b) Production of appressoria and penetration into host hyphae. (c) Appressorium branches produced inside host cells. (d) Disintegration of B. cinerea after 6 days. Mycoparasitic interaction between the two isolates was observed using a fluorescence microscope under phase-contrast, differential interference contrast (DIC), and normal fields.

Figure 6.
Observation of mycoparasitism of C. rosea G67-1 strain to B. cinerea under a scanning electron microscope. (a) G67-1 hyphae coil around those of B. cinerea. (b) Attachment to and penetration into B. cinerea hyphae by C. rosea appressoria. (c) Partial degradation of B. cinerea cell walls. (d) Attachment to and penetration into B. cinerea spores. B, B. cinerea; C, C. rosea.
Figure 6.
Observation of mycoparasitism of C. rosea G67-1 strain to B. cinerea under a scanning electron microscope. (a) G67-1 hyphae coil around those of B. cinerea. (b) Attachment to and penetration into B. cinerea hyphae by C. rosea appressoria. (c) Partial degradation of B. cinerea cell walls. (d) Attachment to and penetration into B. cinerea spores. B, B. cinerea; C, C. rosea.

Figure 7.
Colonization of C. rosea G67-1 strain and its mycoparasitism against B. cinerea TC-B1 on tomato leaves observed under a fluorescence microscope. (a) G67-1 hyphae growing towards B. cinerea. (b) G67-1 hyphae growing alongside the mycelia of TC-B1. (c) Fungal colonies derived from surface-sterilized tomato leaflets on PDA plates. (d) Mycelia of GFP-labelled C. rosea and B. cinerea isolates overlapping regions of PDA plates.
Figure 7.
Colonization of C. rosea G67-1 strain and its mycoparasitism against B. cinerea TC-B1 on tomato leaves observed under a fluorescence microscope. (a) G67-1 hyphae growing towards B. cinerea. (b) G67-1 hyphae growing alongside the mycelia of TC-B1. (c) Fungal colonies derived from surface-sterilized tomato leaflets on PDA plates. (d) Mycelia of GFP-labelled C. rosea and B. cinerea isolates overlapping regions of PDA plates.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Control efficacy of GFP-labelled C. rosea 67-1 transformants against soybean Sclerotinia stem rot.
Table 1.
Control efficacy of GFP-labelled C. rosea 67-1 transformants against soybean Sclerotinia stem rot.
| Strain | Disease Index | Control Efficacy (%) |
|---|---|---|
| G67-1 | 28.4 ± 1.7 b | 55.9 ± 1.9 a |
| 67-1 | 26.9 ± 1.9 b | 58.2 ± 2.2 a |
| CK | 64.3 ± 1.1 a | - |
Data are means ± standard deviation (SD) of three replicates. Different letters in a column indicate significant differences at p < 0.05.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Hasan, R.; Lv, B.; Uddin, M.J.; Chen, Y.; Fan, L.; Sun, Z.; Sun, M.; Li, S. Monitoring Mycoparasitism of Clonostachys rosea against Botrytis cinerea Using GFP. J. Fungi 2022, 8, 567. https://doi.org/10.3390/jof8060567
AMA Style
Hasan R, Lv B, Uddin MJ, Chen Y, Fan L, Sun Z, Sun M, Li S. Monitoring Mycoparasitism of Clonostachys rosea against Botrytis cinerea Using GFP. Journal of Fungi. 2022; 8(6):567. https://doi.org/10.3390/jof8060567
Chicago/Turabian StyleHasan, Rakibul, Binna Lv, Md. Jamal Uddin, Yingying Chen, Lele Fan, Zhanbin Sun, Manhong Sun, and Shidong Li. 2022. "Monitoring Mycoparasitism of Clonostachys rosea against Botrytis cinerea Using GFP" Journal of Fungi 8, no. 6: 567. https://doi.org/10.3390/jof8060567
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.