
Cell Surface B2m-Free Human Leukocyte Antigen (HLA) Monomers and Dimers: Are They Neo-HLA Class and Proto-HLA?
 ,
,
Abstract
:1. Introduction
2. Early Reports on the Formation of Cell Surface B2m-Free HCs (Face-2)
2.1. Are Cell Surface B2m-Free HCs Ephemeral Due to the Dissociation of B2m?
2.2. Can HLA HCs Emerge on the Cell Surface as B2m-Free HCs?
3. Research on B2m-Free HLA Variants on Tumor Cells
3.1. Do Proinflammatory Cytokines Induce the Expression of Face-2 on Tumor Cells?
3.2. Post-Translational Expression of Face-2 Independent of Face-1 in Cancer Cells
3.3. Does Face-2 Replace the Loss of Face-1 in Human Cancers?
- Type 1:
- Tumor cells can lose all of the six HLA-I alleles present in the normal cells;
- Type 2:
- Tumor cells can lose a single HLA haplotype or one set of HLA-Ia genes localized on chromosome 6 (HLA-A, B, and C);
- Type 3:
- Tumor cells can downregulate an HLA-A, B or C locus, producing a phenotype with only four HLA-Ia alleles,
- Type 4:
- Tumor cells can lose a single HLA-I allele out of the six expressed by somatic cells;
- Type 5:
- Tumor cells can lose HLA-Ia (classical) alleles and upregulate the expression of HLA-Ib (non-classical) haplotypes, namely HLA-E, HLA-F, and HLA-G), primarily based on the correlation between the loss of HLA-Ia, concomitant with the upregulation of HLA-Ib.
4. Is Cell Surface Expression of Face-2 Common on Activated and Virus-Transformed Immune Cells?
5. Normal Tissues Express Low Levels of Face-2
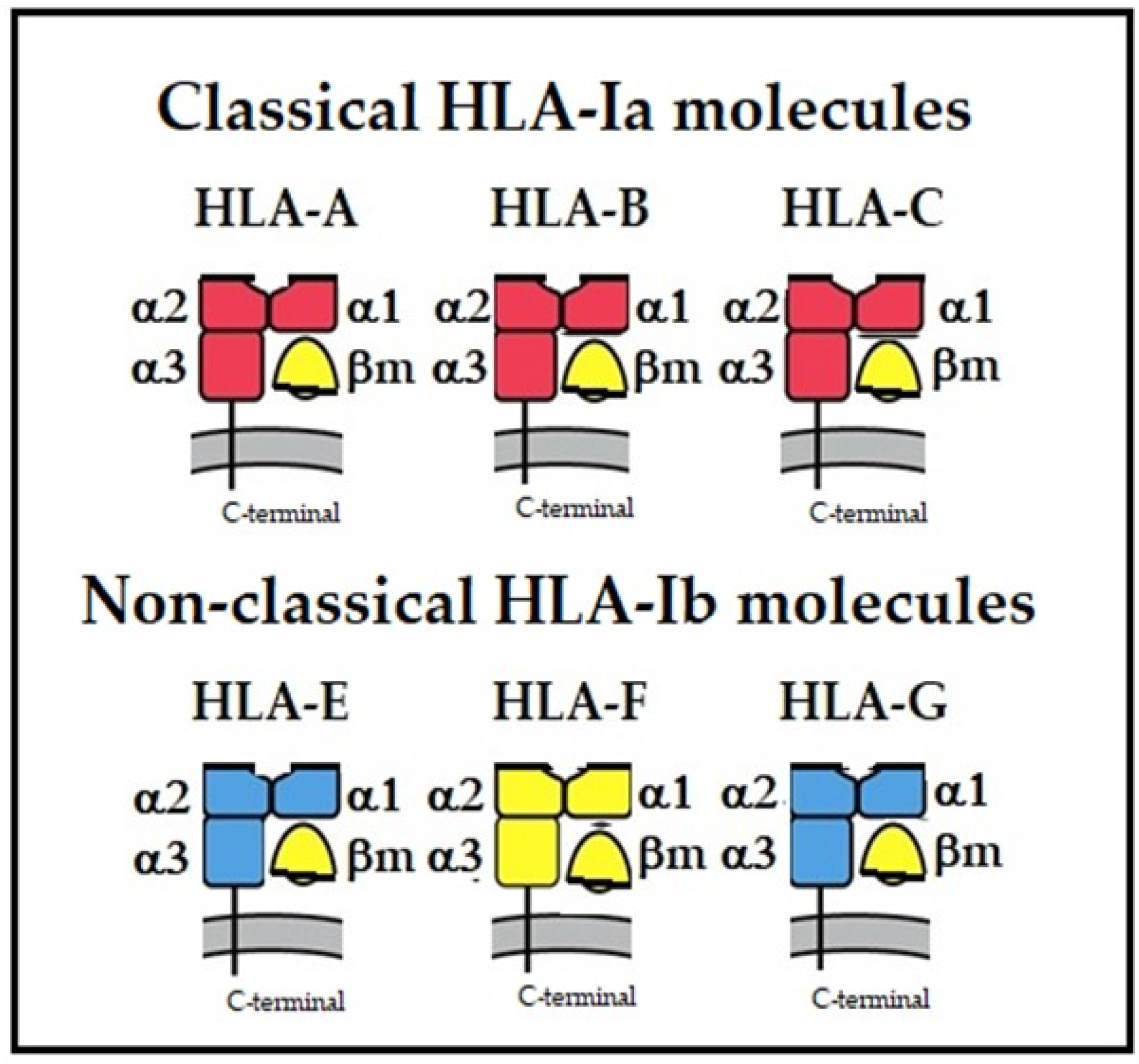
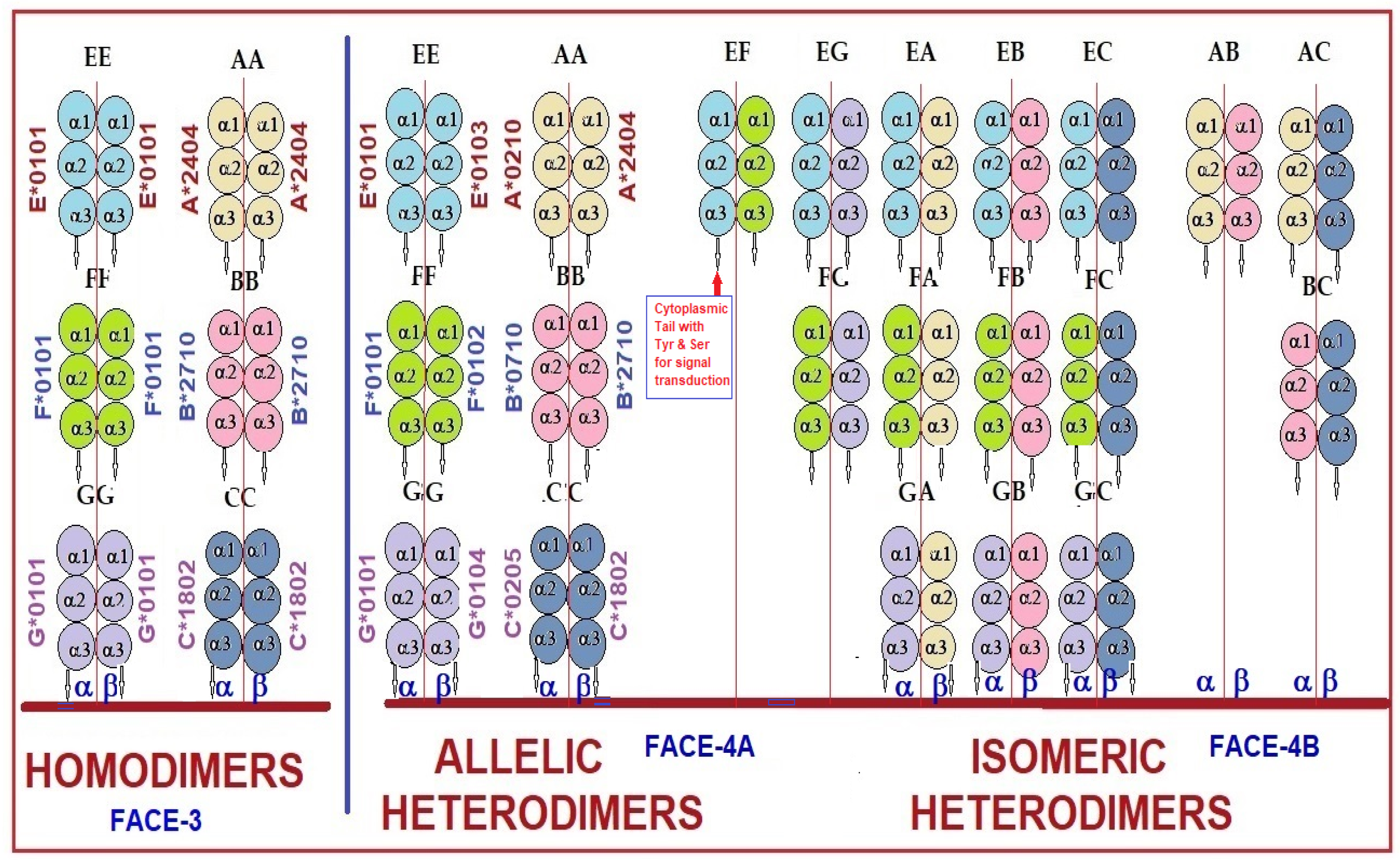
6. Monomeric and Dimeric Variants among Non-Classical HLA Molecules
7. Does Face-2 of HLA B27 Alleles Play a Role in Spondyloarthropathies?
8. Current Concepts Concerning Oligomerization of Face-2
8.1. Is the Cell Surface Clustering of Face-2 a Causal Factor for Dimerization to Generate Face-3 and Face-4 Variants?
8.2. Is Dimerization a Necessary Prerequisite for Face-2 on the Cell Surface?
8.3. Do Cysteine Residues in Face-2 Facilitate and Stabilize Dimerization?
8.4. Documentation of Cysteine-42-Mediated Homodimerization of HLA-G
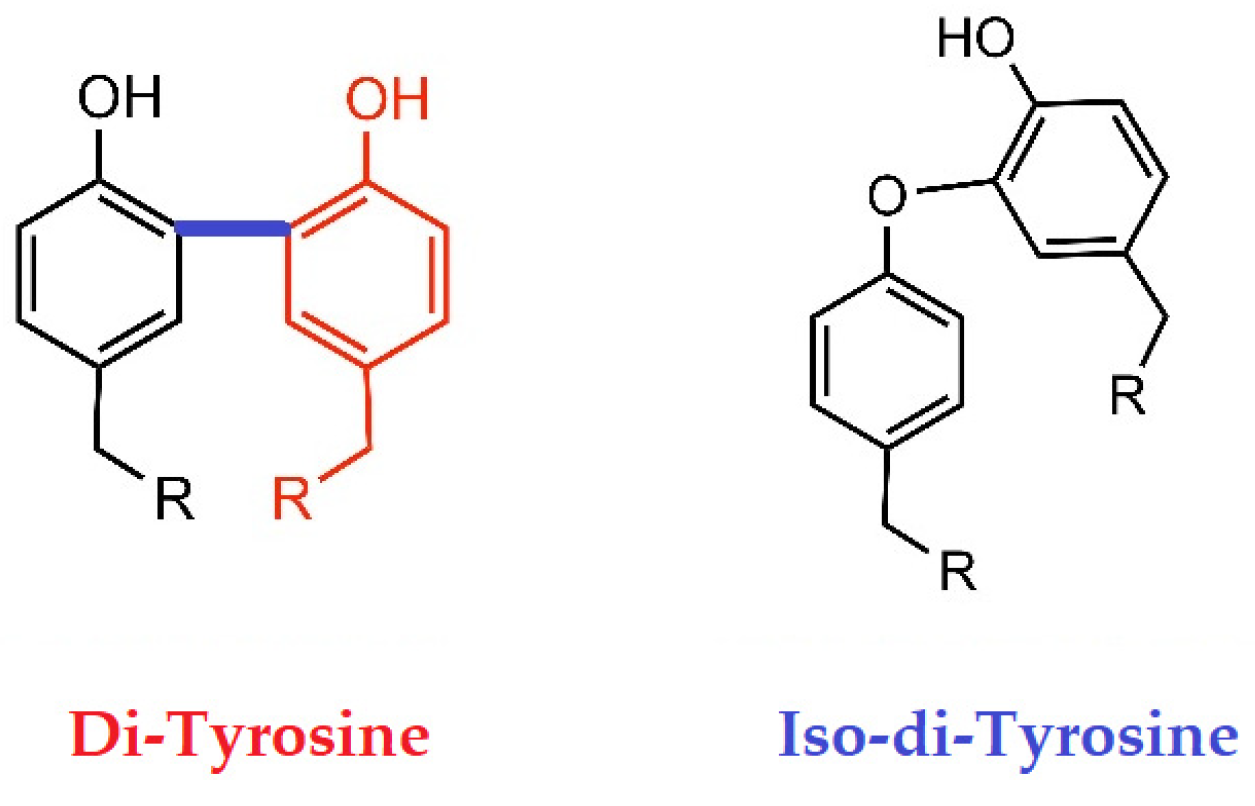
8.5. Does Oxidative Stress Play a Role in Dimerization?
9. Diversified Functional Capabilities of B2m-Free HLA Variants
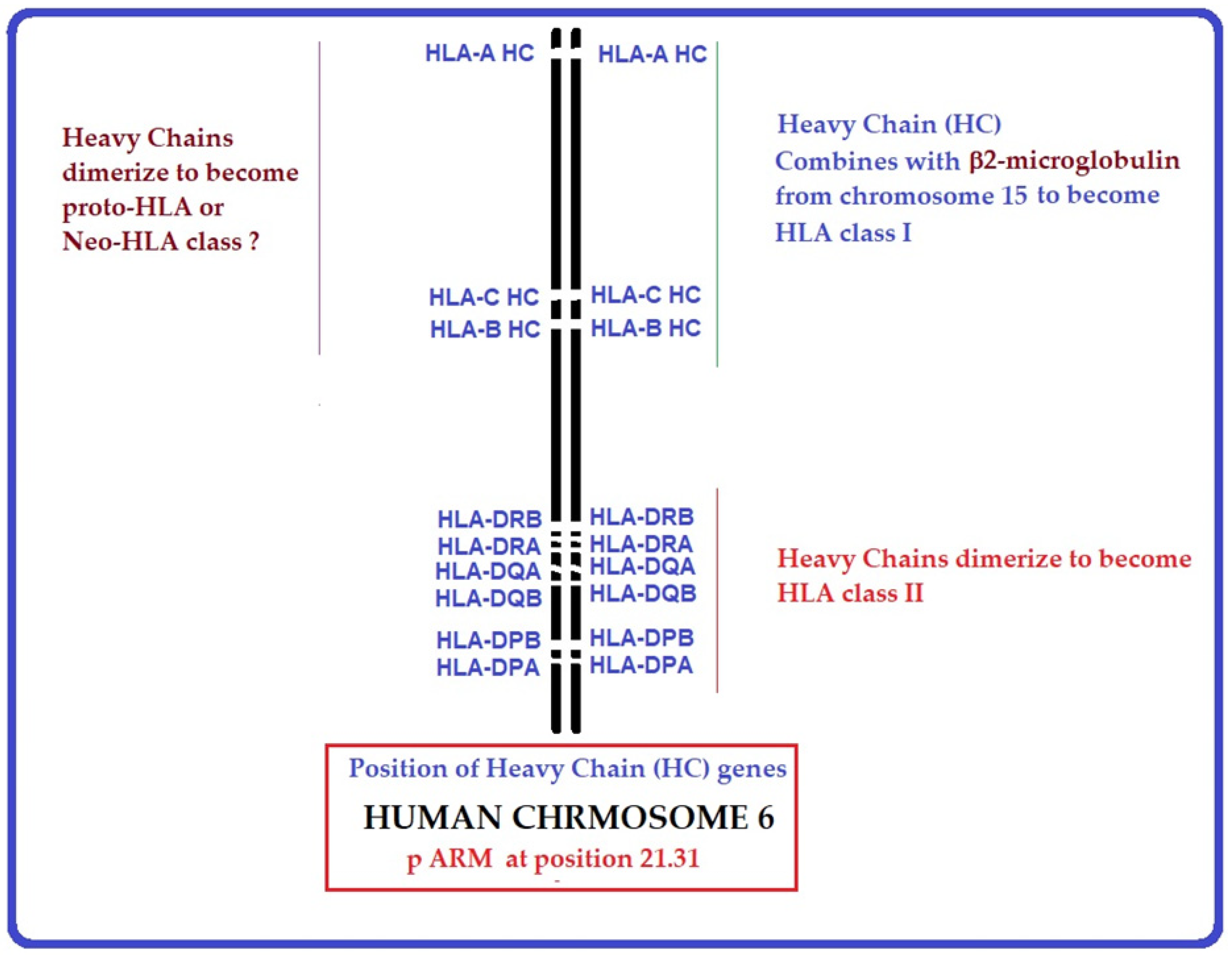
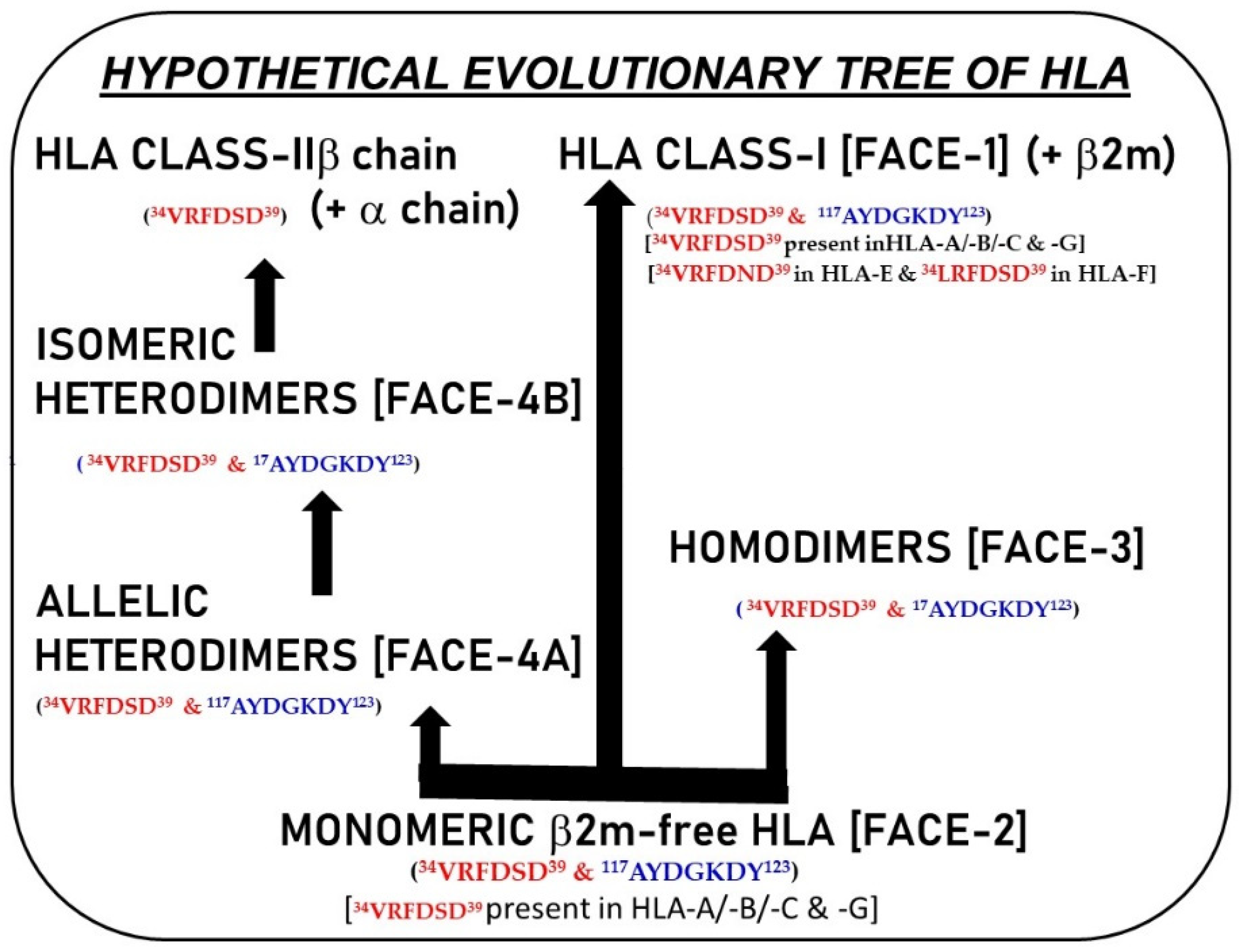
10. Are B2m-Free HC Variants the Evolutionary Progenitors of HLA-I and II and May Represent Proto-HLA?
11. Summary: Genesis, Transformation, and Functions of HLA Variants (Neo-HLA)
- (a)
- Inducing the expression of B2m-free HCs with PMA on activated T cells and blocking their exit with BFA from the ER together confirms that the induction of Face-2 on activated cells requires the direct transport via ER independent of their association with B2m.
- (b)
- In a second experiment, they stripped Face-2 from PMA-activated T cells by brief treatment with trypsin in the cold. The treatment of these cells with trypsin selectively removed surface B2m-free HCs but had no effect on B2m-associated HLA (Face-1) (p. 107, Lines 19–21, [11]). The treated cells were then incubated at 37 °C in the presence or absence of BFA. They noted that the BFA did not block Face-2 reappearance on these trypsin-treated activated T cells, indicating that they are dissociated from B2m-associated HLAs. They immunostained with mAb HC10 to confirm that the emergence of Face-2 is due to dissociation of B2m from Face-1.
- (i)
- HLA–HCs associated with B2m (HLA-I);
- (ii)
- HLA–HC dimers belonging to HLA-II (DP/DQ/DR);
- (iii)
- The monomers, homo-, and hetero-dimers of HLA–HCs (HLA-A/-B/-C/-E/-F/-G).
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Wu, Y.; Zhang, N.; Hashimoto, K.; Xia, C.; Dijkstra, J.M. Structural comparison between MHC Classes I and II; in evolution, a Class-II-like molecule probably came first. Front. Immunol. 2021, 12, 621153. [Google Scholar] [CrossRef] [PubMed]
- Owen, M.J.; Crumpton, M.J. Biochemistry of major human histocompatibility antigens. Immunol. Today 1980, 1, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Lefranc, M.P.; Duprat, E.; Kaas, Q.; Tranne, M.; Thiriot, A.; Lefranc, G. IMGT unique numbering for MHC groove G-DOMAIN and MHC superfamily (MhcSF) G-LIKE-DOMAIN. Dev. Comp. Immunol. 2005, 29, 917–938. [Google Scholar] [CrossRef] [PubMed]
- Lefranc, M.-P. Immunoglobulin and T Cell Receptor Genes: IMGT® and the Birth and Rise of Immunoinformatics. Front. Immunol. 2014, 5, 22. [Google Scholar] [CrossRef] [Green Version]
- Ploegh, H.L.; Orr, H.T.; Strominger, J.L. Major histocompatibility antigens: The human (HLA-A, -B, -C) and murine (H-2K, H-2D) class I molecules. Cell 1981, 24, 287–299. [Google Scholar] [CrossRef]
- Due, C.; Simonsen, M.; Olsson, L. The major histocompatibility complex class I heavy chain as a structural subunit of the human cell membrane insulin receptor: Implications for the range of biological functions of histocompatibility antigens. Proc. Natl. Acad. Sci. USA 1986, 83, 6007–6011. [Google Scholar] [CrossRef] [PubMed]
- Samson, M.; Cousin, J.L.; Fehlman, M. Cross-linking of insulin receptors to MHC antigens in human B lymphocytes: Evidence for selective molecular interactions. J. Immunol. 1986, 137, 2293–2298. [Google Scholar] [CrossRef]
- Arosa, F.A.; Santos, S.G.; Powis, S.J. Open conformers: The hidden face of MHC-I molecules. Trends Immunol. 2007, 28, 115–123. [Google Scholar] [CrossRef]
- Ravindranath, M.H.; Ravindranath, N.M.; Selvan, S.R.; Filippone, E.J.; Amato-Menker, C.J.; El Hilali, F. Four Faces of Cell-Surface HLA Class-I: Their Antigenic and Immunogenic Divergence Generating Novel Targets for Vaccines. Vaccines 2022, 10, 339. [Google Scholar] [CrossRef]
- Ravindranath, M.H.; El Hilali, F.; Amato-Menker, C.J.; El Hilali, H.; Selvan, S.R.; Filippone, E.J. Role of HLA-I Structural Variants and the Polyreactive Antibodies They Generate in Immune Homeostasis. Antibodies 2022, 11, 58. [Google Scholar] [CrossRef]
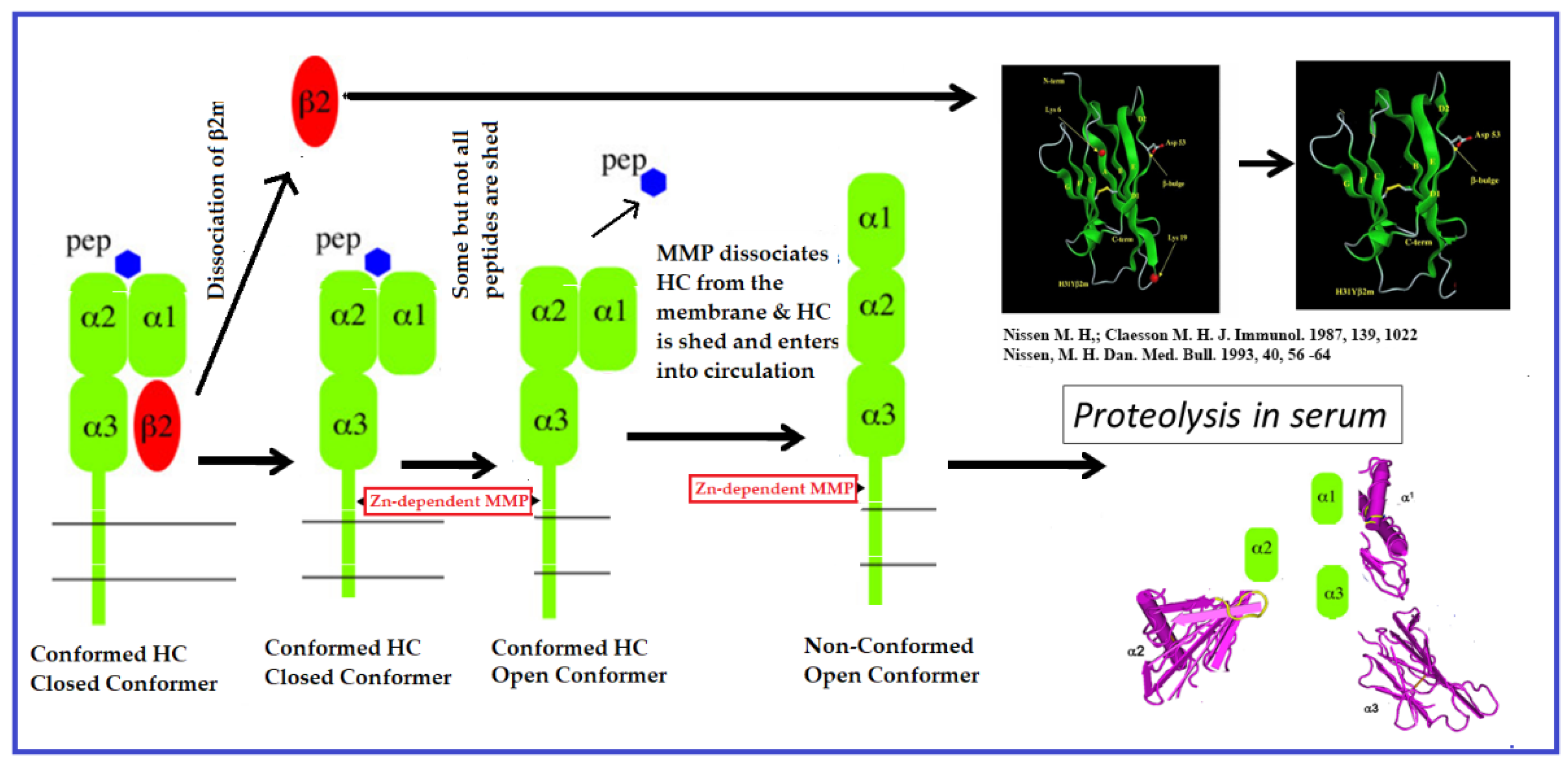
- Demaria, S.; Schwab, R.; Bushkin, Y. The origin and fate of beta 2m-free MHC class I molecules induced on activated T cells. Cell Immunol. 1992, 142, 103–113. [Google Scholar] [CrossRef] [PubMed]
- DeVito-Haynes, L.D.; Demaria, S.; Bushkin, Y.; Burlingham, W.J. The metalloproteinase-mediated pathway is essential for generation of soluble HLA class I proteins by activated cells in vitro: Proposed mechanism for soluble HLA release in transplant rejection. Hum. Immunol. 1998, 59, 426–434. [Google Scholar] [CrossRef] [PubMed]
- Demaria, S.; DeVito-Haynes, L.D.; Salter, R.D.; Burlingham, W.J.; Bushkin, Y. Peptide-conformed beta2m-free class I heavy chains are intermediates in generation of soluble HLA by the membrane-bound metalloproteinase. Hum. Immunol. 1999, 60, 1216–1226. [Google Scholar] [CrossRef] [PubMed]
- Elliott, T. How do peptides associate with MHC class I molecules? Immunol. Today 1991, 12, 386–388. [Google Scholar] [CrossRef]
- Elliott, T.; Cerundolo, V.; Elvin, J.; Townsend, A. Peptide-induced conformational change of the class I heavy chain. Nature 1991, 351, 402–406. [Google Scholar] [CrossRef]
- Cerundolo, V.; Elliott, T.; Elvin, J.; Bastin, J.; Rammensee, H.G.; Townsend, A. The binding affinity and dissociation rates of peptides for class I major histocompatibility complex molecules. Eur. J. Immunol. 1991, 21, 2069–2075. [Google Scholar] [CrossRef]
- Bix, M.; Raulet, D. Functionally conformed free class I heavy chains exist on the surface of β2 microglobulin negative cells. J. Exp. Med. 1992, 176, 829–834. [Google Scholar] [CrossRef]
- Nissen, M.H.; Claësson, M.H. Proteolytically modified human beta 2-microglobulin augments the specific cytotoxic activity in murine mixed lymphocyte culture. J. Immunol. 1987, 139, 1022–1029. [Google Scholar] [CrossRef]
- Nissen, M.H. Proteolytic modification of beta 2-microglobulin in human serum. Dan. Med. Bull. 1993, 40, 56–64. [Google Scholar]
- Fellous, M.; Kamoun, M.; Wiels, J.; Dausset, J.; Clements, C.; Zeuthen, J.; Klein, G. Induction of HLA expression in Daudi cells after cell fusion. lmmunogenetics 1977, 5, 423. [Google Scholar] [CrossRef]
- Hyman, R.; Stallings, V. Analysis of hybrids between a H-2+TL − lymphoma and an H-2÷TL + lymphoma and its H-2-TL- variant subline. Immunogenetics 1977, 4, 171. [Google Scholar] [CrossRef]
- Krangel, M.S.; Orr, H.T.; Strominger, J.L. Assembly and maturation of HLA-A and HLA-B antigens in vivo. Cell 1979, 18, 979–991. [Google Scholar] [CrossRef]
- Potter, T.A.; Boyer, C.; Verhulst, A.M.; Golstein, P.; Rajan, T.V. Expression of H-2Db on the cell surface in the absence of detectable beta 2 microglobulin. J. Exp. Med. 1984, 160, 317–322. [Google Scholar] [CrossRef]
- Potter, T.A.; Zeff, R.A.; Schmitt-Verhulst, A.M.; Rajan, T.V. Molecular analysis of an EL4 cell line that expresses H-2Db but not H-2Kb or beta 2-microglobulin. Proc. Natl. Acad. Sci. USA 1985, 82, 2950–2954. [Google Scholar] [CrossRef] [PubMed]
- Allen, H.; Fraser, J.; Flyer, D.; Calvin, S.; Flavell, R. Beta 2-microglobulin is not required for cell surface expression of the murine class I histocompatibility antigen H-2Db or of a truncated H-2Db. Proc. Natl. Acad. Sci. USA 1986, 83, 7447–7451. [Google Scholar] [CrossRef]
- Giacomini, P.; Aguzzi, A.; Tecce, R.; Fisher, P.B.; Ferrone, S. A third polypeptide associated with heavy and light chain subunits of class I HLA antigens in immune interferon-treated human melanoma cells. Eur. J. Immunol. 1985, 15, 946–951. [Google Scholar] [CrossRef] [PubMed]
- Rebaï, N.; Malissen, B. Structural and genetic analyses of HLA class I molecules using monoclonal xenoantibodies. Tissue Antigens 1983, 22, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Bushkin, Y.; Posnett, D.N.; Pernis, B.; Wang, C.Y. A new HLA-linked T cell membrane molecule, related to the chain of the clonotypic receptor, is associated with T3. J. Exp. Med. 1986, 164, 458–473. [Google Scholar] [CrossRef]
- Bushkin, Y.; Demaria, S.; Le, J.; Schwab, R. Physical association between the CD8 and HLA class I molecules on the surface of activated human T lymphocytes. Proc. Natl. Acad. Sci. USA 1988, 85, 3985–3989. [Google Scholar] [CrossRef]
- Marozzi, A.; Meneveri, R.; Bunone, G.; De Santis, C.; Lopalco, L.; Beretta, A.; Agresti, A.; Siccardi, A.G.; Della Valle, G.; Ginelli, E. Expression of beta 2m-free HLA class I heavy chains in neuroblastoma cell lines. Scand. J. Immunol. 1993, 37, 661–667. [Google Scholar] [CrossRef]
- Grassi, F.; Meneveri, R.; Gullberg, M.; Lopalco, L.; Rossi, G.B.; Lanza, P.; De Santis, C.; Brattsand, G.; Buttò, S.; Ginelli, E.; et al. Human immunodeficiency virus type I gpl20 mimics a hidden epitope borne by Class I major histocompatibility complex heavy chains. J. Exp. Med. 1991, 174, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Martayan, A.; Fiscella, M.; Setini, A.; Ciccarelli, G.; Gambari, R.; Feriotto, G.; Beretta, A.; Siccardi, A.G.; Appella, E.; Giacomini, P. Conformation and surface expression of free HLA-CW1 heavy chains in the absence of beta 2-microglobulin. Hum. Immunol. 1997, 53, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Giacomini, P.; Beretta, A.; Nicotra, M.R.; Ciccarelli, G.; Martayan, A.; Cerboni, C.; Lopalco, L.; Bini, D.; Delfino, L.; Ferrara, G.B.; et al. HLA-C heavy chains free of beta2-microglobulin: Distribution in normal tissues and neoplastic lesions of non-lymphoid origin and interferon-y responsiveness. Tissue Antigens 1997, 50, 555–566. [Google Scholar] [CrossRef]
- Garrido, F. MHC Class-I Loss and Cancer Immune Escape; Advances in Experimental Medicine and Biology; Springer Nature: Cham, Switzerland, 2019; Volume 1151, pp. 1–101. [Google Scholar]
- Menier, C.; Saez, B.; Horejsi, V.; Martinozzi, S.; Krawice-Radanne, I.; Bruel, S.; Le Danff, C.; Reboul, M.; Hilgert, I.; Rabreau, M.; et al. Characterization of monoclonal antibodies recognizing HLA-G or HLA-E: New tools to analyze the expression of nonclassical HLA class I molecules. Hum. Immunol. 2003, 64, 315–326. [Google Scholar] [CrossRef] [PubMed]
- LoMonaco, E.; Sibilio, L.; Melucci, E.; Tremante, E.; Suchànek, M.; Horejsi, V.; Martayan, A.; Giacomini, P. HLA-E: Strong association with beta2-microglobulin and surface expression in the absence of HLA class I signal sequence-derived peptides. J. Immunol. 2008, 181, 5442–5450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
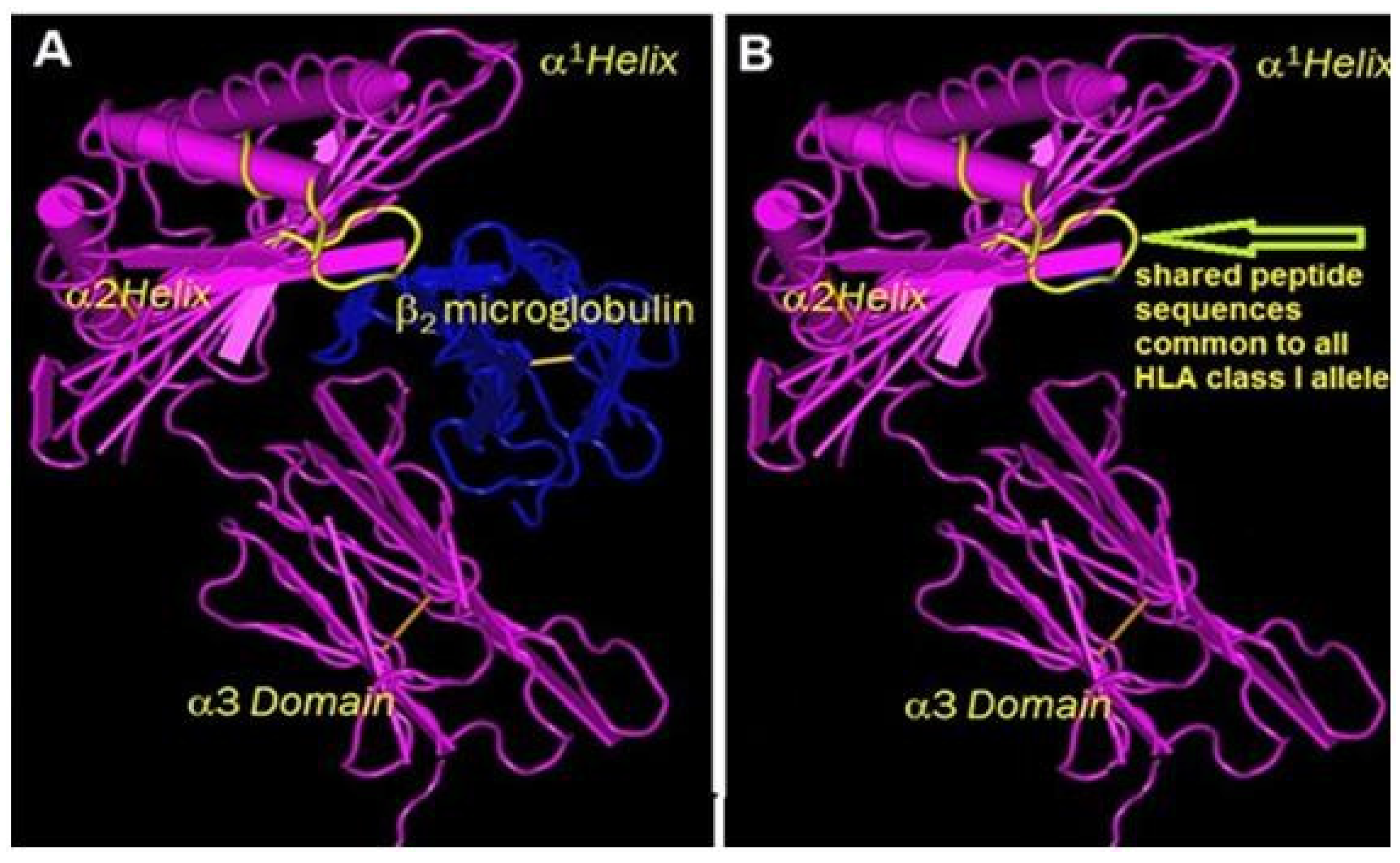
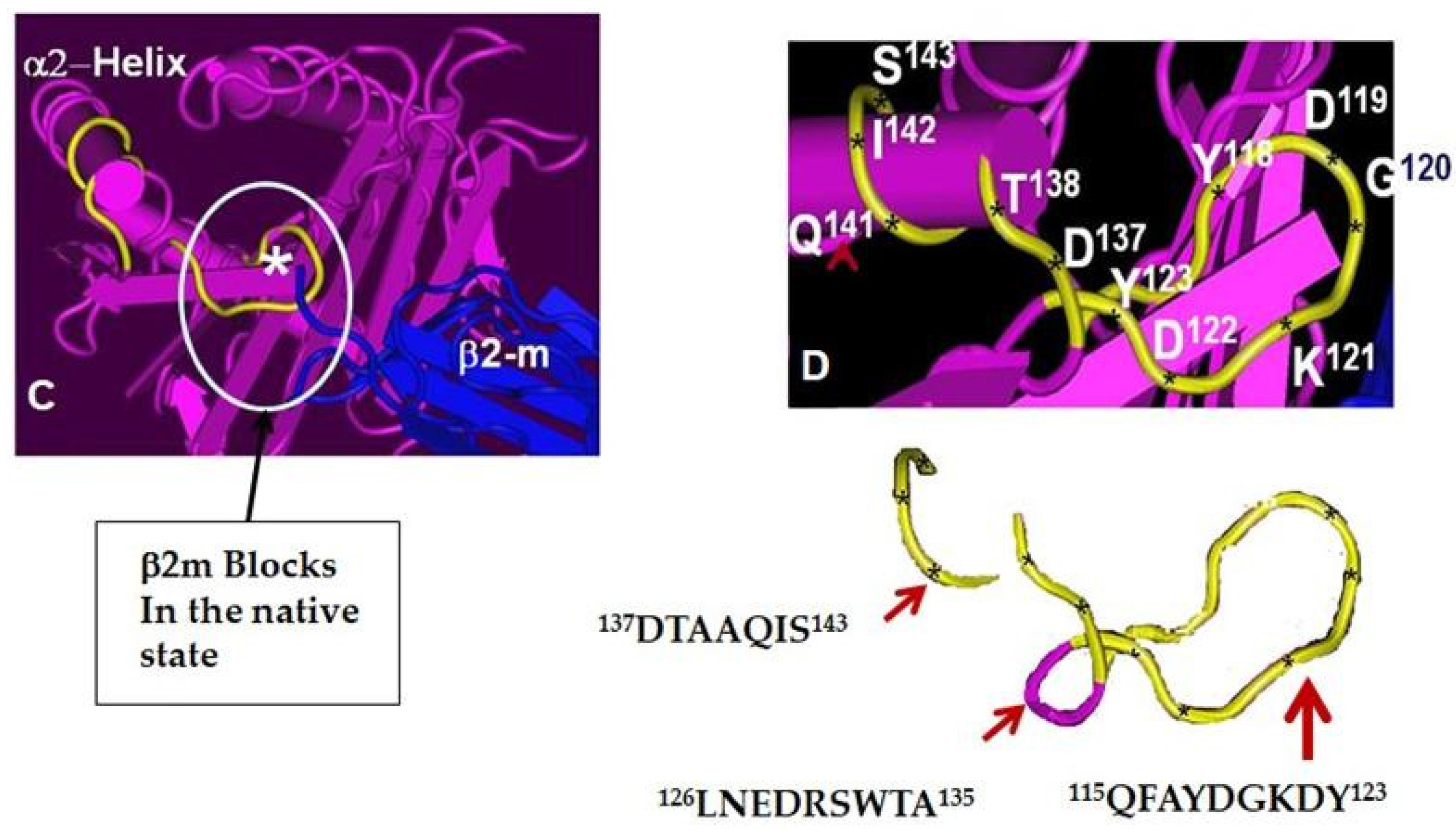
- Ravindranath, M.H.; Taniguchi, M.; Chen, C.W.; Ozawa, M.; Kaneku, H.; El-Awar, N.; Cai, J.; Terasaki, P.I. HLA-E monoclonal antibodies recognize shared peptide sequences on classical HLA class Ia: Relevance to human natural HLA antibodies. Mol. Immunol. 2010, 47, 1121–1131. [Google Scholar] [CrossRef] [PubMed]
- Ravindranath, M.H.; Pham, T.; El-Awar, N.; Kaneku, H.; Terasaki, P.I. Anti-HLAE mAb 3D12 mimics MEM-E/02 in binding to HLA-B and HLA-C alleles: Web-tools validate the immunogenic epitopes of HLA-E recognized by the antibodies. Mol. Immunol. 2011, 48, 423–430. [Google Scholar] [CrossRef]
- Ravindranath, M.H.; Kaneku, H.; El-Awar, N.; Morales-Buenrostro, L.E.; Terasaki, P.I. Antibodies to HLA-E in non-alloimmunized males: Pattern of HLA-Ia reactivity of anti-HLA-E-positive sera. J. Immunol. 2010, 185, 1935–1948. [Google Scholar] [CrossRef] [Green Version]
- Ravindranath, M.H.; Terasaki, P.I. Diagnostic and Therapeutic Potential of HLA-E Monospecific Monoclonal IgG Antibodies Directed against Tumor Cell Surface and Soluble HLA-E. U.S. Patent 10656156, 5 July 2012. [Google Scholar]
- Sasaki, T.; Ravindranath, M.H.; Terasaki, P.I.; Freitas, M.C.; Kawakita, S.; Jucaud, V. Gastric cancer progression may involve a shift in HLA-E profile from an intact heterodimer to β2-microglobulin-free monomer. Int. J. Cancer 2014, 134, 1558–1570. [Google Scholar] [CrossRef]
- Ravindranath, M.H.; Terasaki, P.I.; Pham, T.; Jucaud, V. The Monospecificity of Novel Anti-HLA-E Monoclonal Antibodies Enables Reliable Immunodiagnosis, Immunomodulation of HLA-E, and Upregulation of CD8+ T Lymphocytes. Monoclon. Antib. Immunodiagn. Immunother. 2015, 34, 135–153. [Google Scholar] [CrossRef]
- Ravindranath, M.H.; El Hilali, F. Monospecific and Polyreactive Monoclonal Antibodies against human Leukocyte Antigen E: Diagnostic and Therapeutic relevance. In Monoclonal Antibodies; Rezaei, N., Ed.; Intech Open: London, UK, 2021; ISBN 978-1-83968-369-5. [Google Scholar]
- Ravindranath, M.H.; Ravindranath, N.M.; El Hilali, F.; Selvan, S.R.; Filippone, E.J. Ramifications of the HLA-I Allelic Reactivity of Anti-HLA-E*01:01 and Anti-HLA-E*01:03 Heavy Chain Monoclonal Antibodies in Comparison with Anti-HLA-I IgG Reactivity in Non-Alloimmunized Males, Melanoma-Vaccine Recipients, and End-Stage Renal Disease Patients. Antibodies 2022, 11, 18. [Google Scholar] [CrossRef]
- Ravindranath, M.H.; Filippone, E.J.; Mahowald, G.; Callender, C.; Babu, A.; Saidman, S.; Ferrone, S. Significance of the intraindividual variability of HLA IgG antibodies in renal disease patients observed with different beadsets monitored with two different secondary antibodies on a Luminex platform. Immunol. Res. 2018, 66, 584–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravindranath, M.H.; Ravindranath, N.M.; Amato-Menker, C.J. Luminex multiplex bead Assay monitoring HLA IgG antibodies in sensitized pre- and post-transplant patients: Clonality of the detection antibody impacts specificity and sensitivity. Appl. Sci. 2021, 11, 6430. [Google Scholar] [CrossRef]
- Ravindranath, M.H.; Terasaki, P.I.; Pham, T.; Jucaud, V.; Kawakita, S. Therapeutic preparations of IVIg contain naturally occurring anti-HLA-E Abs that react with HLA-Ia (HLA-A/-B/-Cw) alleles. Blood 2013, 121, 2013–2028. [Google Scholar] [CrossRef] [Green Version]
- Ravindranath, M.H.; Zhu, D.; Pham, T.; Jucaud, V.; Hopfield, J.; Kawakita, S.; Terasaki, P.I. Anti-HLA-E monoclonal antibodies reacting with HLA-la and lb alleles like IVIg as potential IVIg-immunomimetics: An evolving therapeutic concept. Clin. Transpl. 2013, 2013, 293–305. [Google Scholar]
- Ravindranath, M.H.; Jucaud, V.; Ferrone, S. Monitoring native HLA-I trimer specific antibodies in Luminex multiplex single antigen bead assay: Evaluation of beadsets from different manufacturers. J. Immunol. Methods 2017, 450, 73–80. [Google Scholar] [CrossRef]
- Derré, L.; Corvaisier, M.; Charreau, B.; Moreau, A.; Godefroy, E.; Moreau-Aubry, A.; Jotereau, F.; Gervois, N.J. Expression and release of HLA-E by melanoma cells and melanocytes: Potential impact on the response of cytotoxic effector cells. J. Immunol. 2006, 177, 3100–3107. [Google Scholar] [CrossRef] [Green Version]
- Allard, M.; Oger, R.; Vignard, V.; Percier, J.M.; Fregni, G.; Périer, A.; Caignard, A.; Charreau, B.; Bernardeau, K.; Khammari, A.; et al. Serum soluble HLA-E in melanoma: A new potential immune-related marker in cancer. PLoS ONE 2011, 6, e21118. [Google Scholar] [CrossRef]
- Gonçalves, A.S.; Oliveira, J.P.; Oliveira, C.F.; Silva, T.A.; Mendonça, E.F.; Wastowski, I.J.; Batista, A.C. Relevance of HLA-G, HLA-E and IL-10 expression in lip carcinogenesis. Hum. Immunol. 2016, 77, 785–790. [Google Scholar] [CrossRef]
- Silva, T.G.; Crispim, J.C.; Miranda, F.A.; Hassumi, M.K.; de Mello, J.M.; Simões, R.T.; Souto, F.; Soares, E.G.; Donadi, E.A.; Soares, C.P. Expression of the nonclassical HLA-G and HLA-E molecules in laryngeal lesions as biomarkers of tumor invasiveness. Histol. Histopathol. 2011, 26, 1487–1497. [Google Scholar] [CrossRef]
- Van Esch, E.M.G.; Tummers, B.; Baartmans, V.; Osse, E.M.; Ter Haar, N.; Trietsch, M.D.; Hellebrekers, B.W.J.; Holleboom, C.A.G.; Nagel, H.T.C.; Tan, L.T.; et al. Alterations in classical and nonclassical HLA expression in recurrent and progressive HPV-induced usual vulvar intraepithelial neoplasia and implications for immunotherapy. Int. J. Cancer 2014, 135, 830–842. [Google Scholar] [CrossRef] [PubMed]
- Djajadiningrat, R.S.; Horenblas, S.D.; Heideman, A.M.; Sanders, J.; de Jong, J.; Jordanova, E.S. Classic and nonclassic HLA class I expression in penile cancer and relation to HPV status and clinical outcome. J. Urol. 2015, 193, 1245–1251. [Google Scholar] [CrossRef]
- Mittelbronn, M.; Simon, P.; Löffler, C.; Capper, D.; Bunz, B.; Harter, P.; Schlaszus, H.; Schleich, A.; Tabatabai, G.; Goeppert, B.; et al. Elevated HLA-E levels in human glioblastomas but not in grade I to III astrocytomas correlate with infiltrating CD8+ cells. J. Neuroimmunol. 2007, 189, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Kren, L.; Muckova, K.; Lzicarova, E.; Sova, M.; Vybihal, V.; Svoboda, T.; Fadrus, P.; Smrcka, M.; Slaby, O.; Lakomy, R.; et al. Production of immune-modulatory nonclassical molecules HLA-G and HLA-E by tumor infiltrating ameboid microglia/macrophages in glioblastomas: A role in innate immunity? J. Neuroimmunol. 2010, 220, 131–135. [Google Scholar] [CrossRef] [PubMed]
- Kren, L.; Slaby, O.; Muckova, K.; Lzicarova, E.; Sova, M.; Vybihal, V.; Svoboda, T.; Fadrus, P.; Lakomy, R.; Vanhara, P.; et al. Expression of immune-modulatory molecules HLA-G and HLA-E by tumor cells in glioblastomas: An unexpected prognostic significance? Neuropathology 2011, 31, 129–134. [Google Scholar] [CrossRef]
- Arantes, D.A.C.; Gonçalves, A.S.; Jham, B.C.; Duarte, E.C.B.; de Paula, E.C.; de Paula, H.M.; Mendonça, E.F.; Batista, A.C. Evaluation of HLA-G, HLA-E, and PD-L1 proteins in oral osteosarcomas. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. 2017, 123, e188–e196. [Google Scholar] [CrossRef]
- Moscon, C.; Arantes, D.A.C.; Gonçalves, A.S.; Alencar, R.C.G.; Oliveira, J.C.; Silva, T.A.; Mendonça, E.F.; Batista, A.C. Immunohistochemical investigations on the expression of programmed cell death ligand 1, human leukocyte antigens G and E, and granzyme B in intraoral mucoepidermoid carcinoma. Arch. Oral Biol. 2017, 83, 55–62. [Google Scholar] [CrossRef]
- Reimers, M.S.; Engels, C.C.; Putter, H.; Morreau, H.; Liefers, G.J.; van de Velde, C.J.H.; Kuppen, P.J. Prognostic value of HLA class I, HLA-E, HLA-G and Tregs in rectal cancer: A retrospective cohort study. BMC Cancer 2014, 14, 486. [Google Scholar] [CrossRef] [Green Version]
- Benevolo, M.; Mottolese, M.; Tremante, E.; Rollo, F.; Diodoro, M.G.; Ercolani, C.; Sperduti, I.; Lo Monaco, E.; Cosimelli, M.; Giacomini, P.J. High expression of HLA-E in colorectal carcinoma is associated with a favorable prognosis. Transl. Med. 2011, 9, 184. [Google Scholar] [CrossRef] [Green Version]
- Zeestraten, E.C.; Reimers, M.S.; Saadatmand, S.; Goossens-Beumer, I.J.; Dekker, J.W.; Liefers, G.J.; van den Elsen, P.J.; van de Velde, C.J.; Kuppen, P.J. Combined analysis of HLA class I, HLA-E and HLA-G predicts prognosis in colon cancer patients. Br. J. Cancer. 2014, 110, 459–468. [Google Scholar] [CrossRef]
- Guo, Z.Y.; Lv, Y.G.; Wang, L.; Shi, S.J.; Yang, F.; Zheng, G.X.; Wen, W.H.; Yang, A.G. Predictive value of HLA-G and HLA-E in the prognosis of colorectal cancer patients. Cell Immunol. 2015, 293, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Zhang, D.; Li, F.; Xiao, Z.; Wu, M.; Shi, D.; Xiang, P.; Bao, Z. Loss of Fas expression and high expression of HLA-E promoting the immune escape of early colorectal cancer cells. Oncol. Lett. 2017, 13, 3379–3386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levy, E.M.; Bianchini, M.; Von Euw, E.M.; Barrio, M.M.; Bravo, A.I.; Furman, D.; Domenichini, E.; Macagno, C.; Pinsky, V.; Zucchini, C.; et al. Human leukocyte antigen-E protein is overexpressed in primary human colorectal cancer. Int. J. Oncol. 2008, 32, 633–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levy, E.M.; Sycz, G.; Arriaga, J.M.; Barrio, M.M.; von Euw, E.M.; Morales, S.B.; González, M.; Mordoh, J.; Bianchini, M. Cetuximab-mediated cellular cytotoxicity is inhibited by HLA-E membrane expression in colon cancer cells. Innate Immun. 2009, 15, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Stangl, S.; Gross, C.; Pockley, A.G.; Asea, A.A.; Multhoff, G. lnfluence of Hsp70 and HLA-E on the killing of leukemic blasts by cytokine/Hsp70 peptide-activated human natural killer (NK) cells. Cell Stress Chaperones 2008, 13, 221–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishigami, S.; Arigami, T.; Okumura, I.; Uchikado, Y.; Kita, Y.; Kurahara, H.; Maemura, K.; Kijima, Y.; Ishihara, Y.; Sasaki, K.; et al. Human leukocyte antigen (HLA)-E and HLA-F expression in gastric cancer. Anticancer Res. 2015, 35, 2279–2285. [Google Scholar]
- Chen, A.; Shen, Y.; Xia, M.; Xu, L.; Pan, N.; Yin, Y.; Miao, F.; Shen, C.; Xie, W.; Zhang, J. Expression of the nonclassical HLA class I and MICA/B molecules in human hepatocellular carcinoma. Neoplasma 2011, 58, 371–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yazdi, T.M.; van Riet, S.; van Schadewijk, A.; Fiocco, M.; van Hall, T.; Taube, C.; Hiemstra, P.S.; van der Burg, S.H. The positive prognostic effect of stromal CD8+ tumor-infiltrating T cells is restrained by the expression of HLA-E in non-small cell lung carcinoma. Oncotarget 2016, 7, 3477–3488. [Google Scholar] [CrossRef] [Green Version]
- De Kruijf, E.M.; Sajet, A.; van Nes, J.G.; Natanov, R.; Putter, H.; Smit, V.T.; Liefers, G.J.; van den Elsen, P.J.; van de Velde, C.J.; Kuppen, P.J. HLA-E and HLA-G expression in classical HLA class I-negative tumors is of prognostic value for clinical outcome of early breast cancer patients. J. Immunol. 2010, 185, 7452–7459. [Google Scholar] [CrossRef] [Green Version]
- Da Silva, G.B.; Silva, T.G.; Duarte, R.A.; Neto, N.L.; Carrara, H.H.; Donadi, E.A.; Gonçalves, M.A.; Soares, E.G.; Soares, C.P. Expression of the Classical and Nonclassical HLA Molecules in Breast Cancer. Int. J. Breast Cancer 2013, 2013, 250435. [Google Scholar] [CrossRef] [Green Version]
- Gooden, M.; Lampen, M.; Jordanova, E.S.; Leffers, N.; Trimbos, J.B.; van der Burg, S.H.; Nijman, H.; van Hall, T. HLA-E expression by gynecological cancers restrains tumor-infiltrating CD8⁺ T lymphocytes. Proc. Natl. Acad. Sci. USA 2011, 108, 10656–10661. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, M.A.; Le Discorde, M.; Simões, R.T.; Rabreau, M.; Soares, E.G.; Donadi, E.A.; Carosella, E.D. Classical and non-classical HLA molecules and p16(INK4a) expression in precursors lesions and invasive cervical cancer. Eur. J. Obstet. Gynecol. Reprod. Biol. 2008, 141, 70–74. [Google Scholar] [CrossRef]
- Spaans, V.M.; Peters, A.A.; Fleuren, G.J.; Jordanova, E.S. HLA-E expression in cervical adenocarcinomas: Association with improved long-term survival. J. Transl. Med. 2012, 10, 184. [Google Scholar] [CrossRef] [Green Version]
- Ferns, D.M.; Heeren, A.M.; Samuels, S.; Bleeker, M.C.G.; de Gruijl, T.D.; Kenter, G.G.; Jordanova, E.S. Classical and non-classical HLA class I aberrations in primary cervical squamous- and adenocarcinomas and paired lymph node metastases. J. Immunother. Cancer 2016, 4, 78. [Google Scholar] [CrossRef] [Green Version]
- Zheng, H.; Lu, R.; Xie, S.; Wen, X.; Wang, H.; Gao, X.; Guo, L. Human leukocyte antigen-E alleles and expression in patients with serous ovarian cancer. Cancer Sci. 2015, 106, 522–528. [Google Scholar] [CrossRef] [Green Version]
- Andersson, E.; Poschke, I.; Villabona, L.; Carlson, J.W.; Lundqvist, A.; Kiessling, R.; Seliger, B.; Masucci, G.V. Non-classical HLA-class I expression in serous ovarian carcinoma: Correlation with the HLA-genotype, tumor infiltrating immune cells and prognosis. Oncoimmunology 2015, 5, e1052213. [Google Scholar] [CrossRef] [Green Version]
- Hanak, L.; Slaby, O.; Lauerova, L.; Kren, L.; Nenutil, R.; Michalek, J. Expression pattern of HLA class I antigens in renal cell carcinoma and primary cell line cultures: Methodological implications for immunotherapy. Med. Sci. Monit. 2009, 15, CR638–CR643. [Google Scholar] [PubMed]
- Kren, L.; Valkovsky, I.; Dolezel, J.; Capak, I.; Pacik, D.; Poprach, A.; Lakomy, R.; Redova, M.; Fabian, P.; Krenova, Z.; et al. HLA-G and HLA-E specific mRNAs connote opposite prognostic significance in renal cell carcinoma. Diagn. Pathol. 2012, 7, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zanetti, B.R.; Carvalho-Galano, D.F.; Feitosa, N.L.; Hassumi-Fukasawa, M.K.; Miranda-Camargo, F.A.; Maciel, L.M.; Ribeiro-Silva, A.; Soares, E.G. Differential expression of immune-modulatory molecule HLA-E in non-neoplastic and neoplastic lesions of the thyroid. Int. J. Immunopathol. Pharmacol. 2013, 26, 889–896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kren, L.; Fabian, P.; Slaby, O.; Janikova, A.; Soucek, O.; Sterba, J.; Krenova, Z.; Michalek, J.; Kral, Z. Multifunctional immune-modulatory protein HLA-E identified in classical Hodgkin lymphoma: Possible implications. Pathol. Res. Pract. 2012, 208, 45–49. [Google Scholar] [CrossRef] [PubMed]
- Schnabl, E.; Stockinger, H.; Majdic, O.; Gaugitsch, H.; Lindley, I.J.; Maurer, D.; Hajek-Rosenmayr, A.; Knapp, W. J Activated human T lymphocytes express MHC class I heavy chains not associated with beta 2-microglobulin. Exp. Med. 1990, 171, 1431–1442. [Google Scholar] [CrossRef]
- Madrigal, J.A.; Belich, M.P.; Benjamin, R.J.; Little, A.M.; Hildebrand, W.H.; Mann, D.L.; Parham, P.J. Molecular definition of a polymorphic antigen (LA45) of free HLA-A and -B heavy chains found on the surfaces of activated B and T cells. Exp. Med. 1991, 174, 1085–1095. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.M.; Ivanyi, P.; Hilgert, I.; Brdicka, T.; Pla, M.; Breur, B.; Flieger, M.; Ivasková, E.; Horejsí, V. The epitope recognized by pan-HLA class I-reactive monoclonal antibody W6/32 and its relationship to unusual stability of the HLA-B27/beta2-microglobulin complex. Immunogenetics 2001, 53, 440–446. [Google Scholar] [CrossRef] [PubMed]
- Martayan, A.; Sibilio, L.; Tremante, E.; Lo Monaco, E.; Mulder, A.; Fruci, D.; Cova, A.; Rivoltini, L.; Giacomini, P. Class I HLA folding and antigen presentation in beta 2-microglobulin defective Daudi cells. J. Immunol. 2009, 182, 3609–3617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Misumi, Y.; Misumi, Y.; Miki, K.; Takatsuki, A.; Tamura, G.; Ikehara, Y. Novel blockade by brefeldin A of intracellular transport of secretory proteins in cultured rat hepatocytes. J. Biol. Chem. 1986, 261, 11398–11403. [Google Scholar] [CrossRef] [PubMed]
- Benjamin, R.J.; Madrigal, J.A.; Parham, P. Peptide binding to empty HLA-B27 molecules of viable human cells. Nature 1991, 351, 74–77. [Google Scholar] [CrossRef]
- Setini, A.; Beretta, A.; De Santis, C.; Meneveri, R.; Martayan, A.; Mazzilli, M.C.; Appella, E.; Siccardi, A.G.; Natali, P.G.; Giacomini, P. Distinctive features of the alpha 1- domain alpha helix of HLA-C heavy chains free of beta 2-microglobulin. Hum. Immunol. 1996, 46, 69–81. [Google Scholar] [CrossRef] [PubMed]
- Stam, N.J.; Spits, H.; Ploegh, H.L. Monoclonal antibodies raised against denatured HLA-B locus heavy chains permit biochemical characterization of certain HLA-C locus products. J. Immunol. 1986, 137, 2299–2306. [Google Scholar] [CrossRef] [PubMed]
- Neefjes, J.J.; Ploegh, H.L. Allele and locus-specific differences in cell surface expression and the association of HLA class I heavy chain with beta 2-microglobulin: Differential effects of inhibition of glycosylation on class I subunit association. Eur. J. Immunol. 1988, 18, 801–810. [Google Scholar] [CrossRef] [PubMed]
- Gillet, A.C.; Pérarnau, B.; Mercier, P.; Lemonnier, F.A. Serological analysis of the dissociation process of HLA-B and C class I molecules. Eur. J. Immunol. 1990, 20, 759–764. [Google Scholar] [CrossRef]
- Carreno, B.M.; Hansen, T.H. Exogenous peptide ligand influences the expression and half-life of free HLA class I heavy chains ubiquitously detected at the cell surface. Eur. J. Immunol. 1994, 24, 1285–1292. [Google Scholar] [CrossRef]
- Amiot, L.; Onno, M.; Drenou, B.; Monvoisin, C.; Fauchet, R. HLA-G class I gene expression in normal and malignant hematopoietic cells. Hum. Immunol. 1998, 59, 524–528. [Google Scholar] [CrossRef]
- Le Bouteiller, P.; Lenfant, F. Antigen-presenting function(s) of the non-classical HLA-E, -F and -G class I molecules: The beginning of a story. Res. Immunol. 1996, 147, 301–313. [Google Scholar] [CrossRef]
- Goodridge, J.P.; Burian, A.; Lee, N.; Geraghty, D.E. HLA-F complex without peptide binds to MHC class I protein in the open conformer form. J. Immunol. 2010, 184, 6199–6208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Chu, W.; Geraghty, D.F.; Hunt, J.S. Expression of HLA-G in human mononuclear phagocytes and selective induction by IFN-gamma. J. Immunol. 1996, 156, 4224–4311. [Google Scholar] [CrossRef] [PubMed]
- Gonen-Gross, T.; Achdout, H.; Arnon, T.I.; Gazit, R.; Stern, N.; Horejsi, V.; Goldman-Wohl, D.; Yagel, S.; Mandelboim, O. The CD85J/leukocyte inhibitory receptor-1 distinguishes between conformed and beta2-microglobulin-free HLA-G molecules. J. Immunol. 2005, 175, 4866–4874. [Google Scholar] [CrossRef] [Green Version]
- Gonen-Gross, T.; Mandelboim, O. HLA-G complexes are observed on the cell surface. Hum. Immunol. 2007, 68, 227–232. [Google Scholar] [CrossRef] [PubMed]
- Boyle, L.H.; Gillingham, A.K.; Munro, S.; Trowsdale, J. Selective export of HLA-F by its cytoplasmic tail. J. Immunol. 2006, 176, 6464–6472. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.; Geraghty, D.E. HLA-F surface expression on B cell and monocyte cell lines is partially independent from tapasin and completely independent from TAP. J. Immunol. 2003, 171, 5264–5271. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.; Ishitani, A.; Geraghty, D.E. HLA-F is a surface marker on activated lymphocytes. Eur. J. Immunol. 2010, 40, 2308–2318. [Google Scholar] [CrossRef] [Green Version]
- Dulberger, C.L.; McMurtrey, C.P.; Hölzemer, A.; Neu, K.E.; Liu, V.; Steinbach, A.M.; Garcia-Beltran, W.F.; Sulak, M.; Jabri, B.; Lynch, V.J.; et al. Human Leukocyte Antigen F Presents Peptides and Regulates Immunity through Interactions with NK Cell Receptors. Immunity 2017, 46, 1018–1029. [Google Scholar] [CrossRef] [Green Version]
- Geraghty, D.E.; Wei, X.H.; Orr, H.T.; Koller, B.H. Human leukocyte antigen F (HLA-F). An expressed HLA gene composed of a class I coding sequence linked to a novel transcribed repetitive element. J. Exp. Med. 1990, 171, 1–18. [Google Scholar] [CrossRef]
- Wainwright, S.D.; Biro, P.A.; Holmes, C.H. HLA-F is a predominantly empty, intracellular, TAP-associated MHC class Ib protein with a restricted expression pattern. J. Immunol. 2000, 164, 319–328. [Google Scholar] [CrossRef] [PubMed]
- Goodridge, J.P.; Burian, A.; Lee, N.; Geraghty, D.E. HLA-F and MHC Class I Open Conformers Are Ligands for NK Cell Ig-like Receptors. J. Immunol. 2013, 191, 3553–3562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brewerton, D.A.; Hart, F.D.; Nicholls, A.; Caffrey, M.; James, D.C.; Sturrock, R.D. Ankylosing spondylitis and HL-A 27. Lancet 1973, 1, 904–907. [Google Scholar] [CrossRef] [PubMed]
- Hammer, R.E.; Maika, S.D.; Richardson, J.A.; Tang, J.-P.; Taurog, J.D. Spontaneous inflammatory disease in transgenic rats expressing HLA-B27 and human 2m: An animal model of HLA-B27 associated human disorders. Cell 1990, 63, 1099–1112. [Google Scholar] [CrossRef]
- Benjamin, R.; Parham, P. Guilt by association: HLA-B27 and ankylosing spondylitis. Immunol. Today 1990, 11, 137–142. [Google Scholar] [CrossRef]
- Benjamin, R.; Parham, P. HLA-B27 and disease: A consequence of inadvertent antigen presentation? Rheum. Dis. Clin. N. Am. 1992, 18, 11–21. [Google Scholar] [CrossRef]
- López de Castro, J.A. The pathogenetic role of HLA-B27 in chronic arthritis. Curr. Opin. Immunol. 1998, 10, 59–66. [Google Scholar] [CrossRef]
- Märker-Hermann, E.; Höhler, T. Pathogenesis of human leukocyte antigen B27-positive arthritis. Information from clinical materials. Rheum. Dis. Clin. N. Am. 1998, 24, 865–881. [Google Scholar] [CrossRef]
- Allen, R.L.; Bowness, P.; McMichael, A. The role of HLA-B27 in spondylo- arthritis. Immunogenetics 1999, 50, 220–227. [Google Scholar] [CrossRef]
- McMichael, A.; Bowness, P. HLA-B27: Natural function and pathogenic role in spondyloarthritis. Arthritis Res. 2002, 4 (Suppl. S3), S153–S158. [Google Scholar] [CrossRef] [PubMed]
- Bowness, P. HLA-B27. Annu. Rev. Immunol. 2015, 33, 29–48. [Google Scholar] [CrossRef] [PubMed]
- Khare, S.D.; Luthra, H.S.; David, C.S. Spontaneous inflammatory arthritis in HLA-B27 transgenic mice lacking beta 2-microglobulin: A model of human spondyloarthropathies. J. Exp. Med. 1995, 182, 1153–1158. [Google Scholar] [CrossRef]
- Khare, S.D.; Hansen, J.; Luthra, H.S.; David, C.S. HLA-B27 heavy chains contribute to spontaneous inflammatory disease in B27/human beta2-microglobulin (beta2m) double transgenic mice with disrupted mouse beta2m. J. Clin. Investig. 1996, 98, 2746–2755. [Google Scholar] [CrossRef] [PubMed]
- Vázquez, M.N.; López de Castro, J.A. Similar cell surface expression of beta2-microglobulin-free heavy chains by HLA-B27 subtypes differentially associated with ankylosing spondylitis. Arthritis Rheum. 2005, 52, 3290–3299. [Google Scholar] [CrossRef] [PubMed]
- Bird, L.A.; Peh, C.A.; Kollnberger, S.; Elliott, T.; McMichael, A.J.; Bowness, P. Lymphoblastoid cells express HLA-B27 homodimers both intracellularly and at the cell surface following endosomal recycling. Eur. J. Immunol. 2003, 33, 748–759. [Google Scholar] [CrossRef]
- Brown, M.A.; Pile, K.D.; Kennedy, L.G.; Calin, A.; Darke, C.; Bell, J.; Wordsworth, B.P.; Cornelis, F. HLA class I associations of ankylosing spondylitis in the white population in the United Kingdom. Ann. Rheum Dis. 1996, 55, 268–270. [Google Scholar] [CrossRef] [PubMed]
- Orchard, T.R.; Thiyagaraja, S.; Welsh, K.I.; Wordsworth, B.P.; Hill Gaston, J.S.; Jewell, D.P. Clinical phenotype is related to HLA genotype in the peripheral arthropathies of inflammatory bowel disease. Gastroenterology 2000, 118, 274–278. [Google Scholar] [CrossRef] [PubMed]
- Belaunzaran, M.O.; Kleber, S.; Schauer, S.; Hausmann, M.; Nicholls, F.; Van den Broek, M.; Payeli, S.; Ciurea, A.; Milling, S.; Stenner, F.; et al. HLA-B27-Homodimer-Specific Antibody Modulates the Expansion of Pro-Inflammatory T-Cells in HLA-B27 Transgenic Rats. PLoS ONE 2015, 10, e0130811. [Google Scholar] [CrossRef] [Green Version]
- Chakrabarti, A.; Matko, J.; Rahman, N.A.; Barisas, B.G.; Edidin, M. Self-association of class I major histocompatibility complex molecules in liposome and cell surface membranes. Biochemistry 1992, 31, 7182–7189. [Google Scholar] [CrossRef] [PubMed]
- Matko, J.; Bushkin, Y.; Wei, T.; Edidin, M. Clustering of class I HLA molecules on the surfaces of activated and transformed human cells. J. Immunol. 1994, 152, 3353–3360. [Google Scholar] [CrossRef] [PubMed]
- Damjanovich, S.; Vereb, G.; Schaper, A.; Jenei, A.; Matkó, J.; Starink, P.; Fox, G.Q.; Arndt-Jovin, D.J.; Jovin, T.M. Structural hierarchy in the clustering of HLA class I molecules in the plasma membrane of human lymphoblastoid cells. Proc. Natl. Acad. Sci. USA 1995, 92, 1122–1126. [Google Scholar] [CrossRef] [PubMed]
- Bodnár, A.; Bacsó, Z.; Jenei, A.; Jovin, T.M.; Edidin, M.; Damjanovich, S.; Matkó, J. Class I HLA oligomerization at the surface of B cells is controlled by exogenous beta(2)-microglobulin: Implications in activation of cytotoxic T lymphocytes. Int. Immunol. 2003, 15, 331–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kennedy, P.R.; Barthen, C.; Williamson, D.J.; Davies, D.M. HLA-B and HLA-C differ in their nanoscale organization at cell surfaces. Front. Immunol. 2019, 10, 61. [Google Scholar] [CrossRef] [Green Version]
- Capps, G.G.; Robinson, B.E.; Lewis, K.D.; Zuniga, M.C. In vivo dimeric association of class I MHC heavy chains. Possible relationship to class I MHC heavy chain-beta 2-microglobulin dissociation. J. Immunol. 1993, 151, 159–169. [Google Scholar] [CrossRef]
- Capps, G.G.; Zúñiga, M.C. The cytoplasmic domain of the H-2Ld class I major histocompatibility complex molecule is differentially accessible to immunological and biochemical probes during transport to the cell surface. J. Biol. Chem. 1993, 268, 21263–21270. [Google Scholar] [CrossRef]
- Allen, R.L.; O’Callaghan, C.A.; McMichael, A.J.; Bowness, P. Cutting edge: HLA-B27 can form a novel β2-microglobulin-free heavy chain homodimer structure. J. Immunol. 1999, 162, 5045–5048. [Google Scholar] [CrossRef]
- Tran, T.M.; Satumtira, N.; Dorris, M.L.; May, E.; Wang, A.; Furuta, E.; Taurog, J.D. HLA-B27in transgenic rats forms disulfide-linked heavy chain oligomers and multimers that bind to the chaperone BiP. J. Immunol. 2004, 172, 5110–5119. [Google Scholar] [CrossRef] [Green Version]
- Lynch, S.; Santos, S.G.; Campbell, E.C.; Nimmo, A.M.; Botting, C.; Prescott, A.; Antoniou, A.N.; Powis, S.J. Novel MHC class I structures on exosomes. J. Immunol. 2009, 183, 1884–1891. [Google Scholar] [CrossRef] [Green Version]
- Makhadiyeva, D.; Lam, L.; Moatari, M.; Vallance, J.; Zheng, Y.; Campbell, E.C.; Powis, S.J. MHC class I dimer formation by alteration of the cellular redox environment and induction of apoptosis. Immunology 2012, 135, 133–139. [Google Scholar] [CrossRef]
- Santos, S.G.; Lynch, S.; Campbell, E.C.; Antoniou, A.N.; Powis, S.J. Induction of HLA-B27 heavy chain homodimer formation after activation in dendritic cells. Arthritis Res. Ther. 2008, 4, R100. [Google Scholar] [CrossRef] [Green Version]
- Morales, P.J.; Pace, J.L.; Platt, J.S.; Langat, D.K.; Hunt, J.S. Synthesis of beta(2)-microglobulin-free, disulphide-linked HLA-G5 homodimers in human placental villous cytotrophoblast cells. Immunology 2007, 122, 179–188. [Google Scholar] [CrossRef]
- Little, A.M.; Nössner, E.; Parham, P.J. Dissociation of beta 2-microglobulin from HLA class I heavy chains correlates with acquisition of epitopes in the cytoplasmic tail. J. Immunol. 1995, 154, 5205–5215. [Google Scholar] [CrossRef] [PubMed]
- Breman, E.; van Miert, P.P.; van der Steen, D.M.; Heemskerk, M.H.; Doxiadis, I.I.; Roelen, D.; Claas, F.H.; van Kooten, C. HLA monomers as a tool to monitor indirect allorecognition. Transplantation 2014, 97, 1119–1127. [Google Scholar] [CrossRef] [Green Version]
- Ezeakile, M.; Portik-Dobos, V.; Wu, J.; Horuzsko, D.D.; Kapoor, R.; Jagadeesan, M.; Mulloy, L.L.; Horuzsko, A. HLA-G dimers in the prolongation of kidney allograft survival. J. Immunol. Res. 2014, 2014, 153981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ajith, A.; Portik-Dobos, V.; Nguyen-Lefebvre, A.T.; Callaway, C.; Horuzsko, D.D.; Kapoor, R.; Zayas, C.; Maenaka, K.; Mulloy, L.L.; Horuzsko, A. HLA-G dimer targets Granzyme B pathway to prolong human renal allograft survival. FASEB J. 2019, 33, 5220–5236. [Google Scholar] [CrossRef]
- HoWangYin, K.Y.; Loustau, M.; Wu, J.; Alegre, E.; Daouya, M.; Caumartin, J.; Sousa, S.; Horuzsko, A.; Carosella, E.D.; LeMaoult, J. Multimeric structures of HLA-G isoforms function through differential binding to LILRB receptors. Cell Mol. Life Sci. 2012, 69, 4041–4049. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Zhang, W.; Hernandez-Lopez, P.; Fabelo, E.; Parikh, M.; Mulloy, L.L.; Horuzsko, A. Isoforms of human leukocyte antigen-G and their inhibitory receptors in human kidney allograft acceptance. Hum. Immunol. 2009, 70, 988–994. [Google Scholar] [CrossRef] [PubMed]
- Triantafilou, K.; Triantafilou, M.; Wilson, K.M.; Fernandez, N. Human histocompatibility molecules have the intrinsic ability to form homotypic association. Hum. Immunol. 2000, 61, 585–598. [Google Scholar] [CrossRef]
- Mear, J.P.; Schreiber, K.L.; Munz, C.; Zhu, X.; Stevanovi, S.; Rammensee, H.G.; Rowland-Hones, S.L.; Colbert, R.A. Misfolding of HLA-B27 as a result of its B pocket suggests a novel mechanism for its role in susceptibility to spondyloarthropathies. J. Immunol. 1999, 163, 6665–6670. [Google Scholar] [CrossRef]
- Antoniou, A.N.; Ford, S.; Taurog, J.D.; Butcher, G.W.; Powis, S.J. Formation of HLA-B27 homodimers and their relationship to assembly kinetics. J. Biol. Chem. 2004, 279, 8895–8902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dangoria, N.S.; DeLay, M.L.; Kingsbury, D.J.; Mear, J.P.; Uchanska-Ziegler, B.; Ziegler, A.; Colbert, R.A. HLA-B27 misfolding is associated with aberrant intermolecular disulfide bond formation (dimerization) in the endoplasmic reticulum. J. Biol. Chem. 2002, 277, 23459–23468. [Google Scholar] [CrossRef] [Green Version]
- Taurog, J.D.; Maika, S.D.; Satumtira, N.; Dorris, M.L.; McLean, I.L.; Yanagisawa, H.; Sayad, A.; Stagg, A.J.; Fox, G.M.; Lê O’Brien, A.; et al. Inflammatory disease in HLA-B27 transgenic rats. Immunol. Rev. 1999, 169, 209–223. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, A.; Tsuchiya, N.; Mitsui, H.; Shiota, M.; Ogawa, A.; Tokunaga, K.; Yoshinoya, S.; Juji, T.; Ito, K. Association of HLA-B39 with HLA-B27-negative ankylosing spondylitis and pauciarticular juvenile rheumatoid arthritis in japanese patients. Arthritis Rheum. 1995, 38, 1672–1677. [Google Scholar] [CrossRef] [PubMed]
- Ramos, M.; De Castro, J.A. HLA-B27 and the pathogenesis of spondyloarthritis. Tissue Antigens 2002, 60, 191–205. [Google Scholar] [CrossRef] [PubMed]
- Nössner, E.; Parham, P. Species-specific differences in chaperone interaction of human and mouse major histocompatibility complex class I molecules. J. Exp. Med. 1995, 181, 327–337. [Google Scholar] [CrossRef] [PubMed]
- Whelan, M.A.; Archer, J.R. Chemical reactivity of an HLA-B27 thiol group. Eur. J. Immunol. 1993, 23, 3278–3285. [Google Scholar] [CrossRef]
- Boyson, J.E.; Erskine, R.; Whitman, M.C.; Chiu, M.; Lau, J.M.; Koopman, L.A.; Valter, M.M.; Angelisova, P.; Horejsi, V.; Strominger, J.L. Disulfide bond-mediated dimerization of HLA-G on the cell surface. Proc. Natl. Acad. Sci. USA 2002, 99, 16180–16185. [Google Scholar] [CrossRef]
- Springer, T.A.; Robb, R.J.; Terhorst, C.; Strominger, J.L. Submit and disulfide structure of monomeric and dimeric forms of detergent-soluble HLA antigens. J. Biol. Chem. 1977, 252, 4694–4700. [Google Scholar] [CrossRef]
- Shiroishi, M.; Kuroki, K.; Rasubala, L.; Tsumoto, K.; Kumagai, I.; Kurimoto, E.; Kato, K.; Kohda, D.; Maenaka, K. Structural basis for recognition of the nonclassical MHC molecule HLA-G by the leukocyte Ig-like receptor B2 (LILRB2/LIR2/ILT4/CD85d). Proc. Natl. Acad. Sci. USA 2006, 103, 16412–16417. [Google Scholar] [CrossRef] [PubMed]
- Apps, R.; Gardner, L.; Sharkey, A.M.; Holmes, N.; Moffett, A. A homodimeric complex of HLA-G on normal trophoblast cells modulates antigen-presenting cells via LILRB1. Eur. J. Immunol. 2007, 37, 1924–1937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davies, M.J. Protein oxidation and peroxidation. Biochem. J. 2016, 473, 805–825. [Google Scholar] [CrossRef] [Green Version]
- Hägglund, P.; Mariotti, M.; Davies, M.J. Identification and characterization of protein cross-links induced by oxidative reactions. Expert Rev. Proteom. 2018, 15, 665–681. [Google Scholar] [CrossRef]
- Hawkins, C.L.; Davies, M.J. Detection, identification, and quantification of oxidative protein modifications. J. Biol. Chem. 2019, 294, 19683–19708. [Google Scholar] [CrossRef] [Green Version]
- Jiang, S.; Carroll, L.; Mariotti, M.; Hägglund, P.; Davies, M.J. Formation of protein cross-links by singlet oxygen-mediated disulfide oxidation. Redox Biol. 2021, 41, 101874. [Google Scholar] [CrossRef]
- Fuentes-Lemus, E.; Hägglund, P.; López-Alarcón, C.; Davies, M.J. Oxidative Crosslinking of Peptides and Proteins: Mechanisms of Formation, Detection, Characterization and Quantification. Molecules 2021, 27, 15. [Google Scholar] [CrossRef]
- Jiang, S.; Fuentes-Lemus, E.; Davies, M.J. Oxidant-mediated modification and cross-linking of beta-2-microglobulin. Free Radic. Biol. Med. 2022, 187, 59–71. [Google Scholar] [CrossRef]
- Carrero, J.J.; Stenvinkel, P. Persistent inflammation as a catalyst for other risk factors in chronic kidney disease: A hypothesis proposal. Clin. J. Am. Soc. Nephrol. 2009, 4 (Suppl. S1), S49–S55. [Google Scholar] [CrossRef] [Green Version]
- Popolo, A.; Autore, G.; Pinto, A.; Marzocco, S. Oxidative stress in patients with cardiovascular disease and chronic renal failure. Free Radic. Res. 2013, 47, 346–356. [Google Scholar] [CrossRef]
- Modaresi, A.; Nafar, M.; Sahraei, Z. Oxidative stress in chronic kidney disease. Iran. J. Kidney Dis. 2015, 9, 165–179. [Google Scholar] [PubMed]
- Wratten, M.L.; Galaris, D.; Tetta, C.; Sevanian, A. Evolution of oxidative stress and inflammation during hemodialysis and their contribution to cardiovascular disease. Antioxid. Redox Signal. 2002, 4, 935–944. [Google Scholar] [CrossRef] [PubMed]
- Colombo, G.; Reggiani, F.; Cucchiari, D.; Portinaro, N.M.; Giustarini, D.; Rossi, R.; Garavaglia, M.L.; Saino, N.; Milzani, A.; Badalamenti, S.; et al. Plasma protein-bound di-tyrosines as biomarkers of oxidative stress in end stage renal disease patients on maintenance haemodialysis. BBA Clin. 2017, 7, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Schreiber, A.B.; Schlessinger, J.; Edidin, M. Interaction between major histocompatibility complex antigens and epidermal growth factor receptors on human cells. J. Cell Biol. 1984, 98, 725–731. [Google Scholar] [CrossRef] [PubMed]
- Kittur, D.; Shimizu, Y.; DeMars, R.; Edidin, M. Insulin binding to human B lymphoblasts is a function of HLA haplotype. Proc. Natl. Acad. Sci. USA 1987, 84, 1351–1355. [Google Scholar] [CrossRef] [PubMed]
- Ramalingam, T.S.; Chakrabarti, A.; Edidin, M. Interaction of class I human leukocyte antigen (HLA-I) molecules with insulin receptors and its effect on the insulin-signaling cascade. Mol. Biol. Cell 1997, 8, 2463–2474. [Google Scholar] [CrossRef] [Green Version]
- Claas, F.H.; van der Poel, J.J.; Castelli-Visser, R.; Pool, J.; Chen, R.B.; Xu, K.Y.; van Rood, J.J. Interaction between des-Tyr1-gamma-endorphin and HLA class I molecules: Serological detection of an HLA-A2 subtype. Immunogenetics 1985, 22, 309–314. [Google Scholar] [CrossRef]
- Claas, F.H.J.; van Rood, J.J. The interaction of drugs and endogeneous substances with HLA class-I antigens. Prog. Allergy 1985, 36, 135–150. [Google Scholar]
- Kulkarni, S.; Martin, M.P.; Carrington, M. The Yin and Yang of HLA and KIR in human disease. Semin. Immunol. 2008, 20, 343–352. [Google Scholar] [CrossRef] [Green Version]
- Kollnberger, S.; Bird, L.; Sun, M.Y.; Retiere, C.; Braud, V.M.; McMichael, A.; Bowness, P. Cell-surface expression and immune receptor recognition of HLA-B27 homodimers. Arthritis Rheum. 2002, 46, 2972–2982. [Google Scholar] [CrossRef]
- Chan, A.T.; Kollnberger, S.D.; Wedderburn, L.R.; Bowness, P. Expansion and enhanced survival of natural killer cells expressing the killer immunoglobulin-like receptor KIR3DL2 in spondylarthritis. Arthritis Rheum. 2005, 52, 3586–3595. [Google Scholar] [CrossRef]
- Kollnberger, S.; Chan, A.; Sun, M.Y.; Chen, L.Y.; Wright, C.; di Gleria, K.; McMichael, A.; Bowness, P. Interaction of HLA-B27 homodimers with KIR3DL1 and KIR3DL2, unlike HLA-B27 heterotrimers, is independent of the sequence of bound peptide. Eur. J. Immunol. 2007, 37, 1313–1322. [Google Scholar] [CrossRef] [PubMed]
- Bowness, P.; Ridley, A.; Shaw, J.; Chan, A.T.; Wong-Baeza, I.; Fleming, M.; Cummings, F.; McMichael, A.; Kollnberger, S. Th17 cells expressing KIR3DL2+ and responsive to HLA-B27 homodimers are increased in ankylosing spondylitis. J. Immunol. 2011, 186, 2672–2680. [Google Scholar] [CrossRef] [Green Version]
- Giles, J.; Shaw, J.; Piper, C.; Wong-Baeza, I.; McHugh, K.; Ridley, A.; Li, D.; Lenart, I.; Antoniou, A.N.; DiGleria, K.; et al. HLA-B27 homodimers and free H chains are stronger ligands for leukocyte Ig-like receptor B2 than classical HLA class I. J. Immunol. 2012, 188, 6184–6193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaw, J.; Kollnberger, S. New perspectives on the ligands and function of the killer cell immunoglobulin-like receptor KIR3DL2 in health and disease. Front. Immunol. 2012, 3, 339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong-Baeza, I.; Ridley, A.; Shaw, J.; Hatano, H.; Rysnik, O.; McHugh, K.; Piper, C.; Brackenbridge, S.; Fernandes, R.; Chan, A.; et al. KIR3DL2 binds to HLA-B27 dimers and free H chains more strongly than other HLA class I and promotes the expansion of T cells in ankylosing spondylitis. J. Immunol. 2013, 190, 3216–3224. [Google Scholar] [CrossRef] [Green Version]
- Cauli, A.; Shaw, J.; Giles, J.; Hatano, H.; Rysnik, O.; Payeli, S.; McHugh, K.; Dessole, G.; Porru, G.; Desogus, E.; et al. The arthritis-associated HLA-B*27:05 allele forms more cell surface B27 dimer and free heavy chain ligands for KIR3DL2 than HLA-B*27:09. Rheumatology 2013, 52, 1952–1962. [Google Scholar] [CrossRef] [Green Version]
- Hatano, H.; Shaw, J.; Marquardt, K.; Zhang, Z.; Gauthier, L.; Chanteux, S.; Rossi, B.; Li, D.; Mitchell, J.; Kollnberger, S. The D0 Ig-like Domain Plays a Central Role in the Stronger Binding of KIR3DL2 to B27 Free H Chain Dimers. J. Immunol. 2015, 194, 1591–1601. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Hatano, H.; Shaw, J.; Olde Nordkamp, M.; Jiang, G.; Li, D.; Kollnberger, S. The Leukocyte Immunoglobulin-like Receptor family member LILRB5 binds to HLA-class I heavy chains. PLoS ONE 2015, 10, e0129063. [Google Scholar] [CrossRef]
- Kollnberger, S. The Role of HLA-Class I Heavy-Chain Interactions with Killer-Cell Immunoglobulin-Like Receptors in Immune Regulation. Crit. Rev. Immunol. 2016, 36, 269–282. [Google Scholar] [CrossRef]
- Dębska-Zielkowska, J.; Moszkowska, G.; Zieliński, M.; Zielińska, H.; Dukat-Mazurek, A.; Trzonkowski, P.; Stefańska, K. KIR Receptors as Key Regulators of NK Cells Activity in Health and Disease. Cells 2021, 10, 1777. [Google Scholar] [CrossRef]
- Maiers, M.; Mehr, R.; Raghavan, M.; Kaufman, J.; Louzoun, Y. Editorial: HLA and KIR Diversity and Polymorphisms: Emerging Concepts. Front. Immunol. 2021, 12, 701398. [Google Scholar] [CrossRef]
- Goodridge, J.P.; Lee, N.; Burian, A.; Pyo, C.W.; Tykodi, S.S.; Warren, E.H.; Yee, C.; Riddell, S.R.; Geraghty, D.E. HLA-F and MHC-I open conformerscooperate in a MHC-I antigen cross-presentation pathway. J. Immunol. 2013, 191, 1567–1577. [Google Scholar] [CrossRef] [Green Version]
- Burian, A.; Wang, K.L.; Finton, K.A.; Lee, N.; Ishitani, A.; Strong, R.K.; Geraghty, D.E. HLA -F and MHC-I Open Conformers Bind Natural Killer Cell Ig-Like Receptor KIR3DS1. PLoS ONE 2016, 11, e0163297. [Google Scholar] [CrossRef]
- Chazara, O.; Xiong, S.; Moffett, A. Maternal IR and fetal HLA-C: A fine balance. J. Leuk. Biol. 2011, 90, 703–716. [Google Scholar] [CrossRef]
- Velastegui, E.; Vera, E.; Berghe, V.W.; Muñoz, M.S.; Orellana-Manzano, A. HLA-C: Evolution, epigenetics, and pathological implications in the major histocompatibility complex. Front Genet. 2023, 14, 1206034. [Google Scholar] [CrossRef]
- Lash, G.E.; Robson, S.C.; Bulmer, J.N. Review: Functional role of uterine natural killer (uNK) cells in human early pregnancy decidua. Placenta 2010, 31, S87–S92. [Google Scholar] [CrossRef]
- Ishitani, A.; Sageshima, N.; Lee, N.; Dorofeeva, N.; Hatake, K.; Marquardt, H.; Geraghty, D.E. Protein expression and peptide binding suggest unique and interacting functional roles for HLA-E, F, and G in maternal-placental immune recognition. J. Immunol. 2003, 171, 1376–1384. [Google Scholar] [CrossRef]
- Koopman, L.A.; Kopcow, H.D.; Rybalov, B.; Boyson, J.E.; Orange, J.S.; Schatz, F.; Masch, R.; Lockwood, C.J.; Schachter, A.D.; Park, P.J.; et al. Human decidual natural killer cells are a unique NK cell subset with immunomodulatory potential. J. Exp. Med. 2003, 198, 1201–1212. [Google Scholar] [CrossRef]
- Garcia-Beltran, W.F.; Holzemer, A.; Martrus, G.; Chung, A.W.; Pacheco, Y.; Simoneau, C.R.; Rucevic, M.; Lamothe-Molina, P.A.; Pertel, T.; Kim, T.-E.; et al. Open conformers of HLA-F are high-affinity ligands of the activating NK-cell receptor KIR3DS1. Nat. Immunol. 2016, 17, 1067–1074. [Google Scholar] [CrossRef] [Green Version]
- Dijkstra, J.M.; Grimholt, U. Major Histocompatibility Complex (MHC) Fragment Numbers Alone—In Atlantic Cod and in General—Do Not Represent Functional Variability. F1000Res 2018, 7, 963. [Google Scholar] [CrossRef] [PubMed]
- Dijkstra, J.M.; Grimholt, U.; Leong, J.; Koop, B.F.; Hashimoto, K. Comprehensive Analysis of MHC Class II Genes in Teleost Fish Genomes Reveals Dispensability of the Peptide-Loading DM System in a Large Part of Vertebrates. BMC Evol. Biol. 2013, 13, 260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grimholt, U.; Tsukamoto, K.; Azuma, T.; Leong, J.; Koop, B.F.; Dijkstra, J.M. A Comprehensive Analysis of Teleost MHC Class I Sequences. BMC Evol. Biol. 2015, 15, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dijkstra, J.M.; Yamaguchi, T.; Grimholt, U. Conservation of Sequence Motifs Suggests That the Nonclassical MHC Class I Lineages CD1/PROCR and UT Were Established Before the Emergence of Tetrapod Species. Immunogenetics 2018, 70, 459–476. [Google Scholar] [CrossRef]
- Dijkstra, J.M.; Yamaguchi, T. Ancient Features of the MHC Class II Presentation Pathway, and a Model for the Possible Origin of MHC Molecules. Immunogenetics 2019, 71, 233–249. [Google Scholar] [CrossRef]
- Wu, Y.; Zhang, N.; Wei, X.; Lu, S.; Li, S.; Hashimoto, K.; Dijkstra, J.M.; Xia, C.J. The Structure of a Peptide-Loaded Shark MHC Class I Molecule Reveals Features of the Binding between β2-Microglobulin and H Chain Conserved in Evolution. J. Immunol. 2021, 207, 308–321. [Google Scholar] [CrossRef]
- Kaufman, J.F.; Auffray, C.; Korman, A.J.; Shackelford, D.A.; Strominger, J. The Class II Molecules of the Human and Murine Major Histocompatibility Complex. Cell 1984, 36, 1–13. [Google Scholar] [CrossRef]
- Kaufman, J. Vertebrates and the Evolution of the Major Histocompatibility Complex (MHC) Class I and Class II Molecules. Verh. Dtsch. Zool. Ges. 1988, 81, 131–144. [Google Scholar]
- Kaufman, J.; Andersen, R.; Avila, D.; Engberg, J.; Lambris, J.; Salomonsen, J.; Welinder, K.; Skjødt, K. Different Features of the MHC Class I Heterodimer Have Evolved at Different -Rates. Chicken B-F and Beta 2-Microglobulin Sequences Reveal Invariant Surface Residues. J. Immunol. 1992, 148, 1532–1546. [Google Scholar] [CrossRef]
- Hashimoto, K.; Nakanishi, T.; Kurosawa, Y. Identification of a shark sequence resembling the major histocompatibility complex class I alpha 3 domain. Proc. Natl. Acad. Sci. USA 1992, 89, 2209–2212. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, K.; Okamura, K.; Yamaguchi, H.; Ototake, M.; Nakanishi, T.; Kurosawa, Y. Conservation and Diversification of MHC Class I and Its Related Molecules in Vertebrates. Immunol. Rev. 1999, 167, 81–100. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, K. The Transformation Hypothesis for the Origin of the Major Histocompatibility Complex (MHC) Family Molecules and the Elucidation of the Fundamental Ancient Duplication Unit (FADU) in the Genome. Recent Res. Dev. Immun. 2003, 1, 55–80. [Google Scholar]
- Dirscherl, C.; Löchte, S.; Hein, Z.; Kopicki, J.D.; Harders, A.R.; Linden, N.; Karner, A.; Preiner, J.; Weghuber, J.; Garcia-Alai, M.; et al. Dissociation of β2m from MHC class I triggers formation of noncovalent transient heavy chain dimers. J. Cell Sci. 2022, 135, jcs259489. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HLA-Ia Alleles | mAb | mAb | mAb | mAb MEM Series | HLA-Ia Alleles | mAb | mAb | mAb | mAb MEM Series | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| TFL-033 | TFL-006 | 3D12 | E/02 | E/06 | E/07 | E/08 | TFL-033 | TFL-006 | 3D12 | E/02 | E/06 | E/07 | E/08 | ||
| HLA-E specific | HLA-E non-specific mAbs | HLA-E specific | HLA-E non-specific mAbs | ||||||||||||
| A*01:01 | 1684 | 11,453 | B*07:02 | 1080 | 1910 | 12,951 | 1375 | 618 | |||||||
| A*03:01 | 649 | 789 | 9525 | B*08:01 | 2422 | 1062 | 15,204 | 4281 | |||||||
| A*11:01 | 4600 | 2940 | 7067 | 4992 | 1321 | B*13:01 | 1956 | 1820 | 5700 | 15,934 | 6182 | 3195 | |||
| A*11:02 | 607 | 559 | 9404 | 580 | B*13:02 | 5604 | 1171 | 1326 | 16,249 | 393 | |||||
| A*23:01 | 12,666 | B*14:01 | 2919 | 3135 | 14,876 | 4103 | 1308 | ||||||||
| A*24:02 | 1256 | 4096 | 14,316 | 10,991 | 2739 | B*14:02 | 942 | 12,447 | 634 | ||||||
| A*24:03 | 1924 | 2505 | 15,894 | 9981 | 2094 | B*15:01 | 3440 | 832 | 14,606 | ||||||
| A*25:01 | 788 | 629 | B*15:02 | 1345 | 3250 | 15,676 | 3860 | 1051 | |||||||
| A*29:02 | 1204 | 1593 | 1770 | 643 | B*15:03 | 1874 | 4731 | 15,401 | 571 | ||||||
| A*30:01 | 2232 | 526 | 10,072 | 551 | B*15:10 | 1292 | 768 | 15,482 | |||||||
| A*30:02 | 1368 | 5296 | B*15:12 | 2619 | 1903 | 15,670 | 850 | ||||||||
| A*32:01 | 955 | 603 | B*15:13 | 2833 | 591 | 3400 | 15,450 | 4023 | 982 | ||||||
| A*33:01 | 1079 | 3037 | 1195 | 3469 | 1163 | B*15:16 | 3503 | 13,159 | |||||||
| A*33:03 | 1204 | 1604 | 1228 | 511 | B*18:01 | 1352 | 4392 | 16,138 | 3923 | 1665 | |||||
| A*34:01 | 2362 | 991 | 757 | 754 | B*27:05 | 3518 | 942 | 13,985 | |||||||
| A*36:01 | 4993 | 1219 | 9451 | 1493 | B*27:08 | 4668 | 3264 | 1175 | 14,289 | 640 | |||||
| A*66:01 | 1783 | 571 | B*35:01 | 2834 | 566 | 8716 | 15,768 | 12,917 | 6233 | ||||||
| A*68:01 | 4150 | 664 | B*37:01 | 1137 | 3444 | 14,356 | 3109 | 1871 | |||||||
| A*69:01 | 992 | 917 | B*38:01 | 6219 | 1672 | 968 | 14,774 | 516 | |||||||
| C*01:02 | 3238 | 966 | 3125 | 12,748 | 2998 | 3217 | B*39:01 | 3301 | 3010 | 12,219 | 3825 | 1289 | |||
| C*02:02 | 5570 | 720 | 2567 | 17,083 | 2003 | 1986 | B*40:01 | 8083 | 800 | 3478 | 15,269 | 2662 | |||
| C*03:02 | 15,443 | 1713 | 14,941 | 1705 | 624 | B*40:02 | 763 | 712 | 2442 | 9971 | 2166 | 631 | |||
| C*0303 | 5990 | 571 | 2358 | 14,248 | 1549 | 1067 | B*40:06 | 4678 | 3216 | 9898 | 15,642 | 14,269 | 6208 | ||
| C*03:04 | 10,870 | 2585 | 13,686 | 1875 | 903 | B*41:01 | 3037 | 4987 | 14,635 | 5400 | 1331 | ||||
| C*04:03 | 5809 | 3796 | 1765 | 7077 | 2052 | 703 | B*42:01 | 2163 | 14,286 | ||||||
| C*05:01 | 3635 | 931 | 9263 | 15,923 | 15,435 | 6346 | B*44:02 | 1537 | 2621 | 15,821 | 1525 | ||||
| C*06:02 | 8171 | 3076 | 17,836 | 1914 | 10,260 | B*44:03 | 1584 | 1321 | 2654 | 13,339 | 1625 | ||||
| C*07:02 | 3082 | 1640 | 6680 | 1911 | 7149 | 13,655 | B*45:01 | 4122 | 604 | 3134 | 14,671 | 3234 | 748 | ||
| C*08:01 | 8093 | 592 | 2481 | 15,461 | 2001 | B*46:01 | 4176 | 3042 | 16,173 | 3389 | 1062 | ||||
| C*12:03 | 9256 | 1020 | 1692 | 15,372 | 1035 | 771 | B*47:01 | 2630 | 777 | 8849 | |||||
| C*14:02 | 5143 | 1889 | 12,570 | 1839 | 1637 | B*48:01 | 2822 | 3577 | 10,982 | 514 | |||||
| C*15:02 | 5101 | 2688 | 16,008 | 918 | 1080 | B*49:01 | 3201 | 1588 | 15,804 | ||||||
| C*16:01 | 3833 | 530 | 1128 | 15,803 | 590 | 735 | B*50:01 | 4895 | 769 | 15,575 | |||||
| C*17:01 | 7054 | 5554 | 1869 | 1361 | 841 | B*51:01 | 6464 | 841 | 2485 | 13,576 | 2619 | 884 | |||
| C*18:02 | 1402 | 1095 | 7779 | 14,530 | 11,223 | 8373 | B*51:02 | 3766 | 2303 | 13,138 | 2352 | 800 | |||
| B*52:01 | 1168 | 1416 | 928 | 8975 | |||||||||||
| B*53:01 | 2324 | 2754 | 13,622 | 2890 | 1384 | ||||||||||
| B*54:01 | 1186 | 1910 | 13,506 | 1274 | 598 | ||||||||||
| B*55:01 | 3525 | 1287 | 14,420 | 892 | 180 | ||||||||||
| B*56:01 | 6569 | 5352 | 16,067 | 5075 | 1881 | ||||||||||
| B*57:01 | 4535 | 588 | 3626 | 11,746 | 5398 | 1982 | |||||||||
| B*57:03 | 388 | 1143 | 2586 | 14,723 | 3272 | 1556 | |||||||||
| B*58:01 | 2668 | 823 | 1636 | 11,721 | 1809 | 1155 | |||||||||
| B*59:01 | 4289 | 2803 | 16,222 | 1837 | 915 | ||||||||||
| B*67:01 | 2308 | 1856 | 704 | 12,533 | |||||||||||
| B*73:01 | 5661 | 659 | 5560 | 2363 | 8347 | 3629 | |||||||||
| B*78:01 | 6082 | 4273 | 11,454 | 5927 | 1678 | ||||||||||
| B*81:01 | 7621 | 579 | 1097 | 11,758 | |||||||||||
| B*82:01 | 4317 | 5295 | 15,480 | 6315 | 1922 | ||||||||||
| Cancer Types | MEM Series | Citations |
|---|---|---|
| Melanoma | MEM-E/02 | Derré L et al. [50] |
| Melanoma | Ravindranath M. H. et al. [36] | |
| Melanoma | Allard M et al. [51] | |
| Lip Squamosal cell carcinoma | MEM-E/06 & MEM-E/07 | Goncalves A. S. et al. [52] |
| Laryngeal carcinoma | Silva T. G. et al. [53] | |
| Vulvar intraepithelial carcinoma | van Esch E. M. G. et al. [54] | |
| Penile carcinoma | Djajadiningrat R. S. et al. [55] | |
| Glioblastoma | MEM-E/02 | Mittelbronn, M. et al. [56] |
| Glioblastoma | Kren L et al. [57] | |
| Glioblastoma | Kren L et al. [58] | |
| Oral Osteosarcoma | Arantes D. A. C. et al. [59] | |
| Intraoral mucoepidermoid carcinoma | Moscon C et al. [60] | |
| Rectal Cancer | Reimers M. S. et al. [61] | |
| Colorectal carcinoma | MEM-E/02 | Benevolo M. et al. [62] |
| Colorectal carcinoma | Zeestraten E.C. et al. [63] | |
| Colorectal carcinoma | Guo Z. Y. et al. [64] | |
| Colorectal carcinoma | Huang R. et al. [65] | |
| Colorectal carcinoma | MEM-E/08 | Levy E. M. et al. [66] |
| Colorectal carcinoma | Levy, E. M. et al. [67] | |
| Colon carcinoma and leukemia | MEM-E/02 | Stangl S. et al. [68] |
| Gastric Cancer | Sasaki T. et al., [40] | |
| Gastric Cancer | Ishigami S. et al. [69] | |
| Hepatocellular carcinoma | Chen A. et al. [70] | |
| Non-small cell Lung Carcinoma | Yazdi T. M. et al. [71] | |
| Breast cancer | de Kruijf E. M. et al. [72] | |
| Breast cancer | da Silva G. B. et al. [73] | |
| Ovarian cancer/Cervical cancer | Gooden M et al. [74] | |
| Cervical cancer | Gonçalves M. A. et al. [75] | |
| Cervical cancer | Spaans V. M. et al., [76] | |
| Cervical cancer | Ferns D. M. et al. [77] | |
| Serous Ovarian Adenocarcinoma | Zheng H. et al. [78] | |
| Serous Ovarian Adenocarcinoma | Andersson E. et al. [79] | |
| Renal Cell Carcinoma | Hanak L. et al. [80] | |
| Serous Ovarian Adenocarcinoma | Kren L. et al. [81] | |
| Thyroid cancer | Zanetti B. R. et al. [82] | |
| Hodgkin Lymphoma | Kren L, et al. [83] |
| Allelles | α1 | α2 | α3 | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 7 | 9 | 27 | 59 | 67 | 74 | 84 | 85 | 99 | 101 | 113 | 116 | 118 | 123 | 159 | 164 | 171 | 203 | 209 | |
| B*07:02 | Y | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | C | Y | C | Y | ||||
| B*07:09 | Y | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | C | Y | ||||||
| B *0710 | Y | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | C | Y | C | Y | ||||
| B*07:14 | Y | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | Y | C | Y | ||||
| B*07:17 | Y | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | |||
| B*14:01 | Y | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | Y | C | C | Y | ||||
| B*14:06 | Y | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | Y | C | ||||||
| B*15:02 | Y | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | |||
| B*15:09 | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | Y | Y | C | Y | C | Y | |||
| B*15:10 | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | ||
| B*15:18 | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | ||
| B*15:21 | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | ||
| B*15:23 | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | Y | Y | C | Y | C | Y | |||
| B*15:44 | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | ||||
| B*15:45 | Y | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | Y | C | Y | ||||
| B*15:90 | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | ||||
| B*15:91 | Y | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | C | Y | ||||||
| B*15:92 | Y | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | C | Y | C | Y | ||||
| B*15:93 | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | Y | Y | C | Y | C | Y | |||
| B*15:99 | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | ||||
| B*27:01 | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | Y | C | Y | ||||||
| B*27:02 | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | ||||
| B*27:03 | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | ||||
| B*27:04:01 | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | ||||
| B*27:04:02 | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | |||||
| B*27:05:02 | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | ||||
| B*27:05:03 | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | Y | C | Y | ||||||
| B*27:05:04 | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | Y | C | Y | ||||||
| B*27:05:05 | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | ||||
| B*27:05:06 | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | Y | C | Y | ||||||
| B*27:05:07 | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | Y | C | Y | ||||||
| B*27:05:08 | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | Y | C | Y | ||||||
| B*27:05:09 | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | Y | C | Y | ||||||
| B*27:05 | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | Y | C | Y | ||||||
| B*27:06 | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | Y | Y | C | Y | C | ||||
| B*27:07 | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | |||||
| B*27:08 | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | |||||
| B*27:09 | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | |||||
| B*27:10 | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | Y | C | Y | ||||||
| B*27:11 | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | |||||
| B*27:12 | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | |||||
| B*27:13 | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | |||||
| B*27:14 | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | Y | C | Y | ||||||
| B*27:15 | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | Y | C | Y | ||||||
| B*27:16 | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | Y | C | Y | ||||||
| B*27:17 | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | Y | C | Y | Y | |||||
| B*27:18 | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | ||||||
| B*27:19 | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | Y | C | Y | ||||||
| B*27:20 | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | Y | C | Y | Y | |||||
| B*27:21 | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | Y | Y | C | Y | Y | ||||
| B*27:23 | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | C | Y | |||||||
| B*27:24 | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | Y | Y | C | Y | |||||
| B*27:25 | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | Y | C | Y | ||||||
| B*27:26 | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | Y | C | Y | ||||||
| B*27:27 | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | Y | C | Y | ||||||
| B*27:28 | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | Y | C | |||||||
| B*27:29 | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | ||||||
| B*27:30 | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | Y | C | Y | ||||||
| B*27:31 | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | Y | C | Y | ||||||
| B*27:32 | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | |||||
| B*27:33 | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | Y | C | Y | ||||||
| B*27:34 | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | Y | C | Y | ||||||
| B*27:35 | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | |||||
| B*27:36 | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | |||||
| B*35:26 | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | Y | Y | C | Y | |||||
| B*38:01:01 | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | Y | Y | C | Y | C | Y | |||
| B*38:01:02 | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | Y | Y | C | Y | |||||
| B*38:02:01 | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | Y | Y | C | Y | |||||
| B*38:02:02 | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | Y | Y | C | Y | |||||
| B*38:03 | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | C | Y | |||||||
| B*38:04 | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | Y | Y | C | Y | |||||
| B*38:05 | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | Y | Y | C | Y | C | Y | |||
| B*38:06 | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | Y | Y | C | Y | |||||
| B*38:07 | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | Y | Y | C | Y | |||||
| B*38:08 | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | Y | Y | C | Y | |||||
| B*38:09 | Y | Y | Y | Y | C | Y | Y | C | Y | Y | Y | C | Y | |||||||
| B*38:10 | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | Y | Y | C | Y | |||||
| B*38:11 | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | Y | Y | C | Y | |||||
| B*38:12 | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | Y | Y | C | Y | C | Y | |||
| B*38:13 | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | Y | Y | C | Y | |||||
| B*38:14 | Y | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | C | Y | C | Y | ||||
| B*38:15 | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | Y | Y | C | Y | |||||
| B*39:01:01:01 | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | C | Y | C | Y | |||||
| B*39:01:01:02 | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | C | Y | C | Y | |||||
| B*39:01:03 | Y | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | C | Y | C | Y | ||||
| B*39:01:04 | Y | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | C | Y | ||||||
| B*39:02:01 | Y | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | C | Y | C | Y | ||||
| B*39:02:02 | Y | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | C | Y | C | Y | ||||
| B*39:03 | Y | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | C | Y | C | Y | ||||
| B*39:04 | Y | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | C | Y | C | Y | ||||
| B*39:05 | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | Y | Y | C | Y | C | Y | |||
| B*39:06:01 | Y | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | C | Y | C | Y | ||||
| B*39:06:02 | Y | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | C | Y | C | Y | ||||
| B*39:07 | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | Y | Y | C | Y | |||||
| B*39:08 | Y | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | C | Y | C | Y | ||||
| B*39:09 | Y | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | C | Y | C | Y | ||||
| B*39:10 | Y | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | C | Y | C | Y | ||||
| B*39:11 | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | Y | Y | C | Y | C | Y | |||
| B*39:12 | Y | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | C | Y | ||||||
| B*39:13:L01 | Y | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | C | Y | ||||||
| B*39:13:02 | Y | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | C | Y | C | Y | ||||
| B*39:14 | Y | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | C | Y | ||||||
| B*39:15 | Y | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | C | Y | ||||||
| B*39:16 | Y | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | C | Y | ||||||
| B*39:17 | Y | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | Y | C | Y | |||||
| B*39:18 | Y | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | C | Y | ||||||
| B*39:19 | Y | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | C | Y | ||||||
| B*39:20 | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | Y | Y | C | Y | |||||
| B*39:22 | Y | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | C | Y | ||||||
| B*39:23 | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | C | Y | C | Y | |||||
| B*39:24 | Y | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | C | Y | C | Y | ||||
| B*39:25N | Y | Y | Y | Y | C | Y | Y | Y | C | |||||||||||
| B*39:26 | Y | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | C | Y | ||||||
| B*39:27 | Y | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | C | Y | ||||||
| B*39:28 | Y | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | C | Y | ||||||
| B*39:29 | Y | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | Y | C | Y | |||||
| B*39:30 | Y | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | C | Y | ||||||
| B*39:31 | Y | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | C | Y | ||||||
| B*39:32 | Y | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | C | |||||||
| B*39:33 | Y | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | C | Y | C | Y | ||||
| B*39:34 | Y | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | C | Y | C | Y | ||||
| B*39:35 | Y | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | C | Y | ||||||
| B*39:36 | Y | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | C | Y | C | Y | ||||
| B*39:37 | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | Y | Y | C | Y | |||||
| B*39:38Q | Y | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | Y | C | Y | |||||
| B*39:39 | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | C | Y | |||||||
| B*39:40N | Y | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | ||||||||
| B*39:41 | Y | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | C | Y | C | Y | ||||
| B*73:01 | Y | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | Y | C | C | Y | ||||
| B*78:03 | Y | Y | Y | Y | C | Y | Y | Y | C | Y | Y | Y | Y | C | ||||||
| B*95:01 | Y | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | C | Y | ||||||
| B*95:08 | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | Y | Y | C | Y | C | Y | |||
| B*95:09 | Y | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | C | Y | ||||||
| B*95:12 | Y | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | |||||
| B*95:13 | Y | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | C | Y | ||||||
| B*95:14 | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | Y | Y | C | Y | C | Y | |||
| B*95:15 | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | Y | Y | C | Y | |||||
| B*95:19 | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | Y | Y | C | Y | |||||
| ALLELES | α1 | α2 | α3 | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 7 | 9 | 27 | 59 | 67 | 84 | 85 | 99 | 101 | 113 | 116 | 118 | 123 | 159 | 164 | 171 | 203 | 209 | |
| E*01:01:0101 | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | |||||
| A*01:01 | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | ||||
| A*01:04 | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | ||||||
| A*02:01 | Y | Y | Y | Y | Y | C | C | Y | Y | Y | Y | Y | C | Y | C | Y | |||
| A*02:07 | Y | Y | Y | Y | Y | C | C | Y | Y | Y | Y | Y | C | Y | C | Y | |||
| A*02:15 | Y | Y | Y | Y | Y | C | C | Y | Y | Y | Y | Y | C | Y | C | Y | |||
| A*02:18 | Y | Y | Y | Y | Y | C | C | Y | Y | Y | Y | Y | C | Y | C | Y | |||
| A*02:10 | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | Y | C | Y | C | Y | |||
| A*23:01 | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | Y | C | Y | C | Y | ||||
| A*23:02 | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | Y | C | Y | ||||||
| A*24:62 | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | |||||||
| A*68:02:01:01 | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | Y | C | Y | C | Y | ||
| A*68:15 | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | Y | C | Y | ||||
| A*92:03 | Y | Y | Y | Y | Y | C | C | Y | Y | Y | Y | Y | C | Y | C | Y | |||
| ALLELES | α1 | α2 | α3 | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 7 | 9 | 27 | 58 | 67 | 84 | 85 | 99 | 101 | 113 | 116 | 118 | 123 | 159 | 164 | 171 | 203 | 209 | |
| E*01:01:01:01 | C | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | |||||
| C*01:02:01 | C | Y | Y | Y | Y | Y | Y | C | C | Y | Y | Y | Y | Y | C | Y | C | Y | |
| C*01:03 | C | Y | Y | Y | Y | Y | Y | C | C | Y | Y | Y | Y | C | Y | C | Y | ||
| C*01:04 | C | Y | Y | Y | Y | Y | Y | C | C | Y | Y | Y | Y | C | Y | C | Y | ||
| C*02:02:01 | C | Y | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | |
| C*02:02:02 | C | Y | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | |
| C*02:10 | C | Y | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | |
| C*02:11 | C | Y | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | |
| C*05:01:01 | C | Y | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | |
| C*05:03 | C | Y | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | |
| C*05:08 | C | Y | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | |
| C*05:09 | C | Y | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | |
| C*06:02:01:01 | C | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | ||
| C*06:02:01:02 | C | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | ||
| C*07:01:01 | C | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | ||
| C*07:01:02 | C | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | ||
| C*07:02:01:01 | C | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | |||
| C*07:02:01:02 | C | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | |||
| C*07:02:01:03 | C | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | |||
| C*07:03 | C | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | |||
| C*07:04:01 | C | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | ||
| C*07:04:02 | C | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | ||
| C*07:06 | C | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | ||
| C*07:11 | C | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | ||
| C*07:18 | C | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | ||
| C*07:19 | C | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | |||
| C*07:26 | C | Y | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | |
| C*07:29 | C | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | |||
| C*07:30 | C | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | ||
| C*07:36 | C | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | ||
| C*07:41 | C | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | ||
| C*07:42 | C | Y | Y | Y | Y | Y | Y | C | F | Y | Y | Y | C | Y | C | Y | |||
| C*08:01:01 | C | Y | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | |
| C*08:02 | C | Y | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | |
| C*08:03 | C | Y | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | |
| C*08:09 | C | Y | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | ||
| C*12:02:01 | C | Y | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | |
| C*12:02:02 | C | Y | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | |
| C*12:03:01:01 | C | Y | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | |
| C*12:04:01 | C | Y | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | ||
| C*12:04:02 | C | Y | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | |
| C*12:05 | C | Y | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | |
| C*12:08 | C | Y | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | |
| C*12:13 | C | Y | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | |
| C*12:14 | C | Y | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | |
| C*12:19 | C | Y | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | |
| C*14:02:01 | C | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | |||
| C*14:03 | C | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | |||
| C*15:02:01 | C | Y | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | C | Y | C | Y | ||
| C*15:03 | C | Y | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | C | Y | C | Y | ||
| C*15:04 | C | Y | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | |
| C*15:05:01 | C | Y | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | C | Y | C | Y | ||
| C*15:05:02 | C | Y | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | C | Y | C | Y | ||
| C*15:06 | C | Y | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | C | Y | C | Y | ||
| C*15:16 | C | Y | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | C | Y | C | Y | ||
| C*15:17 | C | Y | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | C | Y | C | Y | ||
| C*16:01:01 | C | Y | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | |
| C*16:01:02 | C | Y | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | |||
| C*16:02 | C | Y | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | |
| C*16:04:01 | C | Y | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | |
| C*17:01 | G | Y | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | |
| C*17:02 | G | Y | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | |
| C*17:03 | G | Y | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | |
| C*18:01 | C | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | |||
| C*18:02 | C | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | |||
| C*05:01:01 | C | Y | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | |
| C*07:01:01 | C | Y | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | ||
| C*18:01 | C | Y | Y | Y | Y | Y | Y | C | Y | Y | Y | Y | C | Y | C | Y | |||
| HLA class I and Proto-HLA 34V R F D S D . . . . . . E . R A . . . . . . . . E Y W 60 |
| V: VALINE (NON-POLAR); R: ARGININE (+ CHARGE); F: PHENYLALANINE (NON-POLAR); D: ASPARTIC ACID (- CHARGE); S: SERINE (POLAR); E: GLUTAMIC ACID (- CHARGE); A: ALANINE (NON-POLAR); Y: TYROSINE (POLAR); W: TRYPTOPHAN (NON-POLAR) |
| ALLELES EXAMINED for Heavy Chains HLA-A ALLELES 1-504 HLA-B ALLELES 1-846 HLA-C ALLELES 1-277 HLA-G ALLELES 4 HLA-E ALLELES 2 34V R F D N D . . . . . . V . R A . . . . . . . . E Y W 60 HLA-F ALLELE 1 34L R F D S D . . . . . . E . R E . . . . . . . . Q Y W 60 |
| HLA class II 38V R F D S D . . E . R A . . . . . . . . . E Y W 61 ALLELES EXAMINED for β-chain of HLA-II DRB ALLELES 1-512 DRB1*010101–1*030101–1*040101–1*070101–1*080101–1*090102–*110101, DRB1*130101–1*140101–1*150101–1*160101, DRB3*0106, DRB5*0102, DRB5*0202–5*0205; DQB ALLELES 1–71 DQB1*050101–DQB1*020101, DQB1*060101 36V R F D S D . . E . R A . . . . . . . . . E Y W 59 DPB ALLELES 1–123 DPB1*020102–1*030101–1*0402–1*0501–1*0601–1*0801–1*0901–1*1001, DPB1*1102, DPB1*1401, DPB1*1502, DPB1*1601–1*1701–1*1801–1*1901–1*200101 ---- 1*9901 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ravindranath, M.H.; Ravindranath, N.M.; Selvan, S.R.; Hilali, F.E.; Amato-Menker, C.J.; Filippone, E.J. Cell Surface B2m-Free Human Leukocyte Antigen (HLA) Monomers and Dimers: Are They Neo-HLA Class and Proto-HLA? Biomolecules 2023, 13, 1178. https://doi.org/10.3390/biom13081178
Ravindranath MH, Ravindranath NM, Selvan SR, Hilali FE, Amato-Menker CJ, Filippone EJ. Cell Surface B2m-Free Human Leukocyte Antigen (HLA) Monomers and Dimers: Are They Neo-HLA Class and Proto-HLA? Biomolecules. 2023; 13(8):1178. https://doi.org/10.3390/biom13081178
Chicago/Turabian StyleRavindranath, Mepur H., Narendranath M. Ravindranath, Senthamil R. Selvan, Fatiha El Hilali, Carly J. Amato-Menker, and Edward J. Filippone. 2023. "Cell Surface B2m-Free Human Leukocyte Antigen (HLA) Monomers and Dimers: Are They Neo-HLA Class and Proto-HLA?" Biomolecules 13, no. 8: 1178. https://doi.org/10.3390/biom13081178