Oxylipin Response to Acute and Chronic Exercise: A Systematic Review
by
 , , ,
, , ,
Étore F. Signini
1,* ,
,
David C. Nieman
2 ,
,
Claudio D. Silva
1 ,
,
Camila A. Sakaguchi
1 and
Aparecida M. Catai
1 1
Physical Therapy Department, Federal University of São Carlos, São Carlos, SP 13565-905, Brazil
2
North Carolina Research Campus, Appalachian State University, Kannapolis, NC 28081, USA
*
Author to whom correspondence should be addressed.
Metabolites 2020, 10(6), 264; https://doi.org/10.3390/metabo10060264
Submission received: 28 May 2020
/
Revised: 18 June 2020
/
Accepted: 19 June 2020
/
Published: 25 June 2020
(This article belongs to the Special Issue Fatty Acid Metabolism)
Abstract
:Oxylipins are oxidized compounds of polyunsaturated fatty acids that play important roles in the body. Recently, metabololipidomic-based studies using advanced mass spectrometry have measured the oxylipins generated during acute and chronic physical exercise and described the related physiological effects. The objective of this systematic review was to provide a panel of the primary exercise-related oxylipins and their respective functions in healthy individuals. Searches were performed in five databases (Cochrane, PubMed, Science Direct, Scopus and Web of Science) using combinations of the Medical Subject Headings (MeSH) terms: “Humans”, “Exercise”, “Physical Activity”, “Sports”, “Oxylipins”, and “Lipid Mediators”. An adapted scoring system created in a previous study from our group was used to rate the quality of the studies. Nine studies were included after examining 1749 documents. Seven studies focused on the acute effect of physical exercise while two studies determined the effects of exercise training on the oxylipin profile. Numerous oxylipins are mobilized during intensive and prolonged exercise, with most related to the inflammatory process, immune function, tissue repair, cardiovascular and renal functions, and oxidative stress.

1. Introduction
Oxylipins are oxidized polyunsaturated fatty acids (PUFAs) and function as lipid mediators for multiple physiological processes [1,2,3]. Polyunsaturated fatty acid oxidation following release from cell phospholipid membranes occurs from the activity of three enzyme systems including cyclooxygenases (COX), lipoxygenases (LOX), and cytochrome P450 (CYP) enzymes [1,4,5]. The primary fatty acids for oxylipin generation include arachidonic acid (AA), adrenic acid (AdA), linoleic acid (LA), α-linoleic acid (ALA), docosahexaenoic acid (DHA), eicosapentaenoic acid (EPA), and dihomo-γ-linolenic acid (DGLA) [1,5].
Plasma oxylipin levels can be altered in some disease states and are influenced by nutritional status and mental and physiological stressors [2,6,7,8,9,10]. Acute and chronic exercise have a strong effect on inflammation and immune function, and oxylipins may be involved at a regulatory level [2,10,11,12,13]. This potential linkage has generated interest in evaluating the effect of varying exercise workloads on oxylipin generation from COX, LOX, and CYP enzyme systems, and the interactive effects with different forms of nutritional support [10,11]. This interest has been fueled by advances in mass spectrometry (MS) and bioinformatics support that have allowed an ever increasing number of oxylipins to be measured [2]. Additionally, oxylipins are not stored but are generated by enzymatic systems in response to various types of stressors, providing a scaffold to effectively evaluate the influences of stressor doses, nutrition, obesity, medications, and other factors [2,7,8,10].
The scientific area of exercise and oxylipins is emergent, but enough studies have been published to systematically tabulate the types of oxylipins generated during different exercise workloads. The aim of this systematic review was to summarize oxylipin responses to acute and chronic exercise by their enzymatic pathways and to provide insights into potential physiological effects. The conclusions derived from this review will provide an evidence-based framework for future investigations.
2. Results
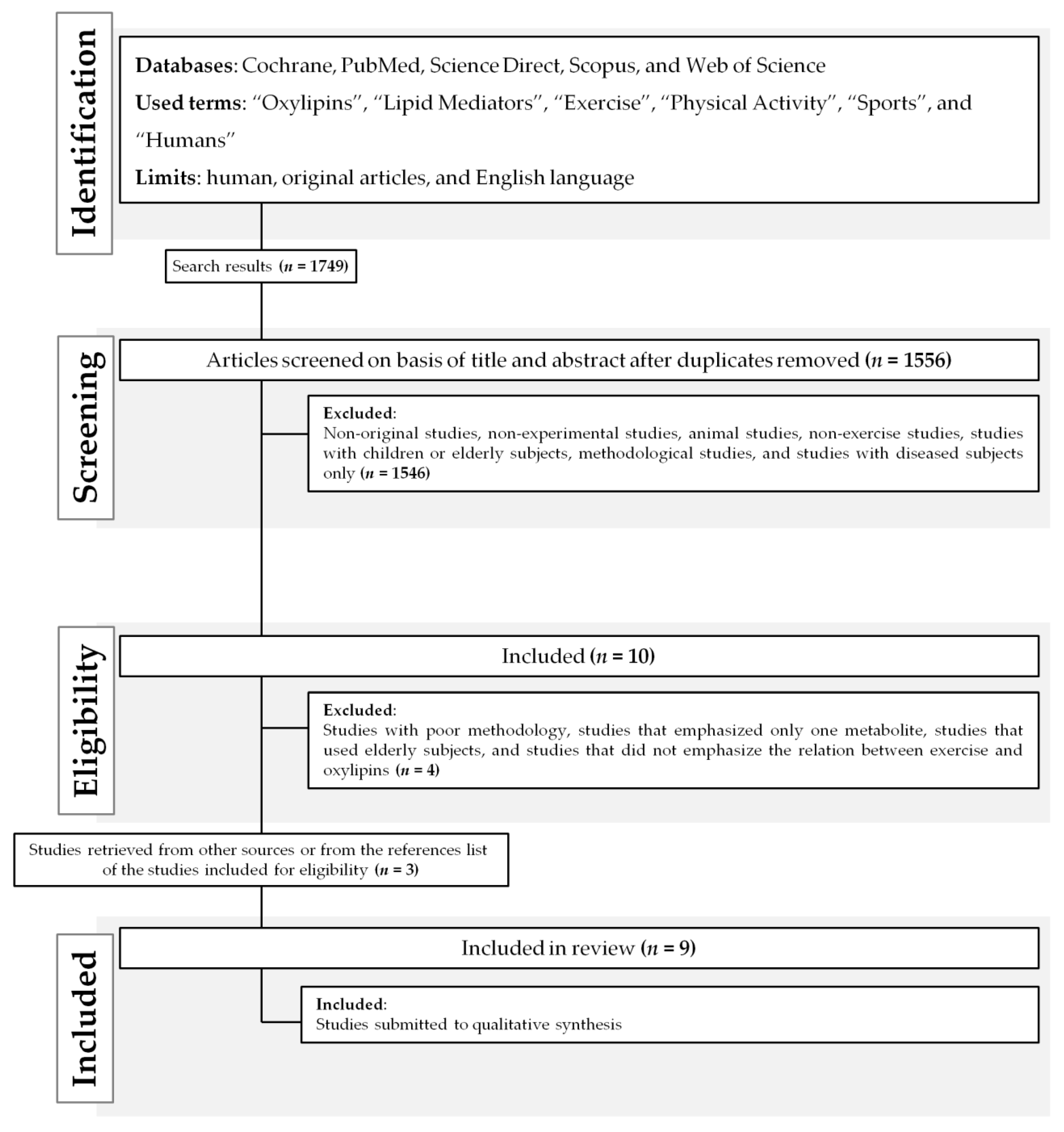
After searching the literature, 1749 documents were identified (Figure 1). Nine papers were included in the final analysis after excluding duplicates and studies that did not meet the inclusion criteria (Table 1). The main reasons for exclusion were the type of study participants (animal-based, children or elderly individuals, and those with a pathology) and the lack of focus on exercise–oxylipin effects.
A scoring system was used to rank the studies for quality of research design, analysis methods, statistical support, and novelty (Table 5). Two studies were classified as having excellent quality [14,15], three as good [16,17,18], one as fair [19], and three as poor [20,21,22]. The detailed score of each study is shown in Table 1.
Table 2 summarizes the main findings and the study design of the nine selected articles. Seven studies focused on oxylipin responses to acute exercise [14,15,16,17,19,20,22] and two studies on chronic physical training [18,21]. Two studies used acute resistance exercise [16,17], one study used a graded, maximal treadmill test [19], and six studies used varying levels of acute or chronic cardiorespiratory exercise [14,15,18,20,21,22].
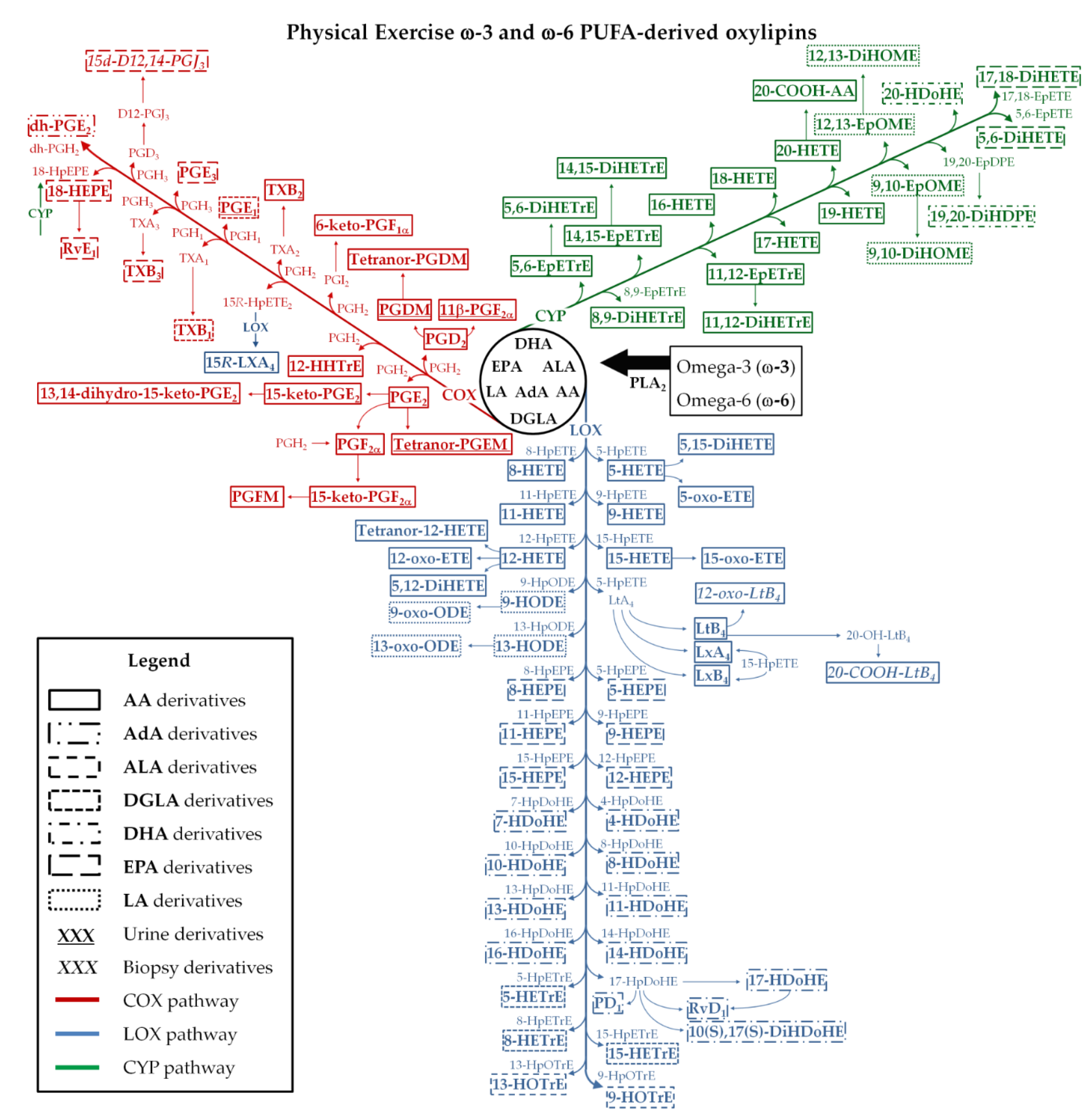
The main metabolic pathways, the oxylipins, and the magnitude of changes reported in the studies are shown in Table 3 and Figure 2. Oxylipins, from the COX, LOX, CYP, and non-enzymatic pathways were reported from urine, serum, plasma, or muscle biopsy samples depending on the research design and MS platform. Some studies included a nutritional [14,15,18] or drug (ibuprofen) [16] intervention, and this systematic review focused on the exercises’ effects on oxylipins and not the nutrition- or ibuprofen-related findings.
García-Flores et al. [18] reported small decreases in urine oxylipin levels in 16 elite triathletes after 15 days of intense training including F2-isoprostanes (F2-IsoPs) and prostaglandin F1α (PGF1α), and small increase in prostaglandins (PGs) 11-β-PGF2α, PGDM, and PGE1. Medina et al. [21] collected urine samples in 15 triathletes before and after a two-week period of intense training, and reported small decreases in F2-IsoPs and PGs (tetranor-PGEM and 11-β-PGF2α), and an increase in 6-keto prostaglandin F1α (6-keto-PGF1α).
Nieman et al. [22] described large-fold increases in LA-directed hydroxyoctadecadienoic acids (9-HODE and 13-HODE), and dihydroxyoctadecenoic acids (9,10-DiHOME and 12,13-DiHOME) in 19 male cyclists after a 75 km cycling protocol. The same author in two other more recent studies, using the same 75 km cycling protocol, reported large-fold increases in plasma levels of 43 of 45 [14] and 64 of 67 [15] oxylipins. Most of the oxylipins were from AA, EPA, and DHA fatty acid substrates, with oxidation through the COX, LOX, and CYP pathways.
Giordano et al. [20] reported small increases in dihydroxyieicosatrienoic acids (8,9-DiHETrE, 11,12-DiHETrE, 14,15-DiHETrE) after 20 min of cycling exercise at 80% of the maximum load. Small increases were shown for epoxyieicosatrienoic acid (14,15-EpETrE) and 14,15-DiHETrE following 40 min of cycling at 60% of the maximum load. Gollasch et al. [19] reported small increases in dihydroxyeicosatetraenoic acids (5,6-DiHETE, 17,18-DiHETE), epoxyoctadecenoic acid (12,13-EpOME) and 5,6-DiHETrE after a maximal graded treadmill test.
Markworth et al. [16] had 16 men engage in an intensive leg resistance exercise routine, and showed small to modest increases in thromboxane B2 (TXB2), PGs (PGE2, PGD2, and PGI2), and their derivatives, leukotriene B4 (LTB4), resolvins (RvE1 and RvD1), isomer of protectin D1 (10(S),17(S)-DiHDoHE), 5,12-DiHETE, hydroxyeicosatetraenoic acids (12-HETE, tetranor-12-HETE, 15-HETE), 15-oxo-eicosatetraenoic acid (15-oxo-ETE), 13-oxo-hydroxyoctadecadienoic acid (13-oxo-ODE), lipoxins (LXA4 and LXB4), 11,12-DiHETrE, and 14,15-DiHETrE.
Vella et al. [17] had 12 men engage in intense knee extension resistance exercise, with muscle biopsies collected pre- and post-exercise, and then after 2 h, 4 h, and 24 h recovery. Small to moderate increases were measured for TXB2, PGE2, PGF2α, 15-Deoxy-Delta12,14-prostaglandin J3 (15d-D12,14-PGJ3), 12-oxo-leukotrieneB4 (12-oxo-LTB4), 20-carboxy leukotriene B4 (20-COOH-LTB4), 5-HETE, 12-HETE, tetranor-12-HETE, 15-HETE, 12-hydroxyeicosapentaenoic acid (12-HEPE), hydroxydocosahexanoic acids (4-HDoHE, 7-HDoHE, and 14-HDoHE) and 5,6-EpETrE, 11,12-DiHETrE and 14,15-DiHETrE.
3. Discussion
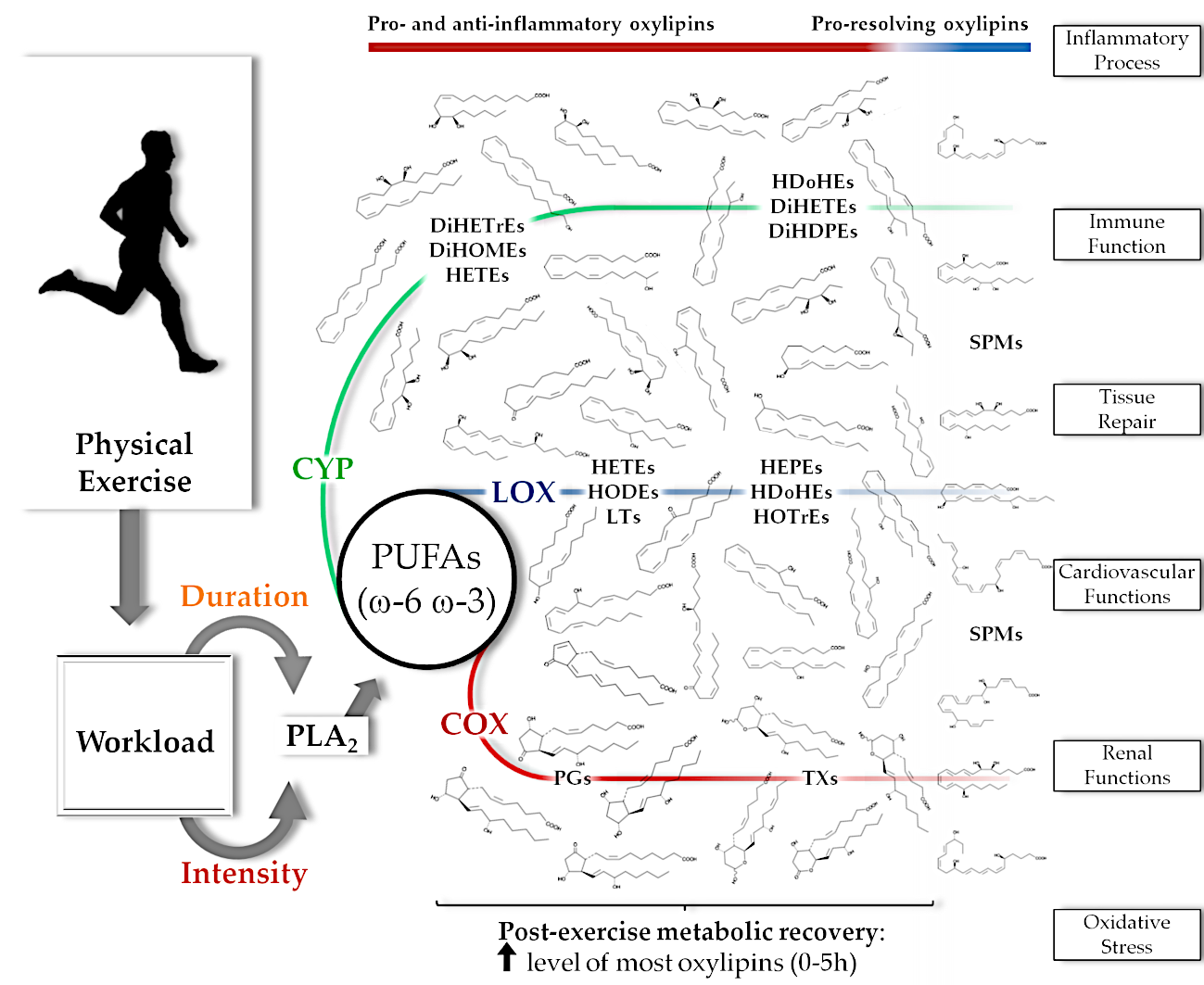
This systematic review provided an overview of the oxylipins that are altered with chronic exercise training or that increase after acute resistance and cardiorespiratory exercise in healthy individuals. Acute exercise induces changes in a high number of oxylipins, especially after prolonged and intensive exercise, and are generated by COX, LOX, CYP and non-enzymatic pathways from multiple fatty acid substrates (Figure 2). The specific roles of oxylipins during and after stressful levels of exercise are still being investigated, and may include regulation of inflammatory and immune system processes, vascular function, and kidney function [1,2,3,6,23,24,25,26]. This systematic review showed that the number of oxylipins generated and the fold increase is dependent on the exercise mode and workload. Plasma levels of oxylipins, even after prolonged and intensive exercise, are close to pre-exercise levels within 5 h of recovery.
3.1. Exercise-Related Oxylipin Formation
The release of PUFAs from cell membranes is stimulated by a group of enzymes identified as phospholipase A2 (PLA2) [1]. The PLA2 enzymes hydrolyze the phospholipids into fatty acids and lysophospholipids. This process may be activated when the cell is stimulated by several types of signaling pathways including mitogen-activated protein kinases (MAPKs) and extracellular signal-regulated kinases (ERK), transcriptional activators (e.g., nuclear factor-kappa B), pro-inflammatory cytokines, and other inflammatory stimuli [14,27,28,29,30,31]. These signaling pathways can be activated by exercise-induced muscle cell membrane injury and metabolic processes [32,33,34] increasing the release of PUFAs, oxylipin generation, and the inflammatory response.
The free PUFAs are subsequently oxidized by COX, LOX, and CYP enzyme pathways that generate the oxylipins [1,5]. The magnitude of increase and the diversity of oxylipins generated by these pathways from omega-6 (ω-6) and omega-3 (ω-3) free PUFAs (AA, AdA, ALA, DGLA, DHA, EPA, and LA) appear to be greatest with prolonged and intensive aerobic exercise workloads (Table 3 and Figure 2). There is scant evidence regarding the physiological roles of oxylipins within an exercise context, but the literature, in general, suggests regulatory roles in inflammatory processes, immune responses, cardiovascular system and kidney function, tissue repair, mitochondrial function, and oxidative stress [1,2,3,10,23,24,25,35,36].
The role of prostaglandins in muscle physiology, inflammation, and injury has been explored for decades [25,37,38,39,40]. The recent emergence of metabololipidomics procedures and bioinformatics support has identified a large number of oxylipins that are generated during exercise, opening up endless pathways for future research [2]. Inflammation regulation during recovery from demanding exercise bouts may emerge as a central role for many of these oxylipins [10,12]. Studies included in this review indicate that pro-inflammatory oxylipins generated during exercise include thromboxanes (TXs), PGs, HETEs, HODEs, and their derivatives (such as oxo-ETEs and oxo-ODEs), leukotrienes (LTs), DiHETrEs, and DiHOMEs (Table 3). These oxylipins are derived from AA, AdA, DGLA, and LA oxidation (Figure 2). The relationship of TXs, PGs, and LTs (such as TXB2, PGE2, and LTB4) with exercise has been extensively studied and are related to pro-inflammatory actions such as the increase of platelet aggregation, leukocyte activation and chemotaxis, pro-inflammatory cytokine production, vessel permeability, nociception, and changes in the vascular tone [2,25,37,38,39,41,42,43]. The recent discovery of transient but large post-exercise elevations in plasma HETEs, HODEs, DiHETrEs, and DiHOMEs underscores the complexity of this area of scientific endeavor [14,15,16,19,20,22]. These oxylipins have similar roles to TXs, PGs, and LTs, but they may also influence mitochondria respiration, skeletal muscle fatty acid uptake, myocardial and skeletal muscle blood flow, blood pressure responses, renal vessel tone and sodium excretion, and oxidative stress [2,3,19,20,22,23,24,36,44,45].
Oxylipins have physiological roles that may vary depending on the metabolic context. For example, some oxylipins are elevated with obesity and various diseases states, but may function as signaling agents during exercise [2,6,10,12]. Certain types of oxylipins exert anti-inflammatory influences to counterbalance the action of pro-inflammatory oxylipins (Table 3) [2]. These oxylipins are typically generated by ω-3 PUFAs such as ALA, DHA, and EPA, with some produced from ω-6 PUFAs [1,2] (Figure 2). Oxylipins from ω-3 PUFAs include hydroxy-octadecatrienoic acids (HOTrEs), HDoHEs, HEPEs and specialized pro-resolving mediators (SPMs), and counter pro-inflammatory actions from innate immune system cells [1,26,46]. Other physiological roles of ω-3 PUFA oxylipins include regulation of vascular tone, blood pressure, production of anti-inflammatory cytokines, tissue repair, and blood clotting [3,26,35,46,47,48]. Omega-6 PUFA oxylipins, such as 15-HETE, EpETrEs and AA lipoxins (LXs), act synergistically with ω-3 PUFA oxylipins [1,2,3,35].
Pro-resolving mediators are involved with inflammation resolution and have produced high scientific interest. They are classified into four families (i.e., lipoxins, maresins, protectins, and resolvins) [49], derived primarily from AA (15-hydroperoxy-eicosatetraenoic acid (15-HpETE)), EPA (18-HEPE), DHA (HDoHEs and hydroperoxy-docosahexaenoic acids (17-HpDoHE and 14-HpDoHE)), and play key roles in resolving inflammation in part by regulating polymorphonuclear (PMNs) leukocytes (e.g., mitigating PMN recruitment and chemotaxis), macrophages (clearance of debris and apoptotic PMNs), tissue regeneration, and nociceptive responses [10,12,26,35,50,51]. Some studies suggest that plasma levels of SPMs (e.g., LXA4, LXB4, protectins (PD1 and 10(S),17(S)-DiHDoHE) and resolvins (RvE1 and RvD1)) increase late in recovery from muscle-damaging exercise, and may regulate tissue regeneration and adaptation (Table 3 and Figure 2) [2,10]. Maresins may not accumulate in plasma even after demanding exercise bouts, but the large increase in 14-HDoHE suggests some involvement since both have the same precursor (14-HpDoHE) (Table 3).
Although this area of scientific endeavor is emerging, available data indicate that ω-6 and ω-3 oxylipin production is dependent on the intensity and duration of physical exercise. This observation is similar to what we have reported with plasma metabolites in general [52]. Data from the studies included in this systematic review showed small-fold changes in plasma levels of oxylipins after a single low intensity, short duration bout of exercise in contrast to large-fold changes after prolonged and intensive exercise (Table 2 and Table 3). The data also indicate that the increase of pro-inflammatory oxylipins tend to occur early in exercise recovery (TXs, PGs, HETEs and HODEs) with an increase of SPMs later in recovery [2,10]. Moreover, the studies also support that most oxylipins return to near pre-exercise levels within five hours of recovery [14,15,16,17,22]. These findings have implications for evaluating the influence of drugs and nutritional supplements on exercise-induced changes in oxylipins.
The chronic effect of physical exercise on the COX, LOX, and CYP pathways and oxylipin generation is largely unknown, and studies published thus far have numerous study design limitations [18,21]. Reported changes in plasma oxylipins with exercise training are variable and of small magnitude. Larger long-term exercise training studies are needed to confirm whether or not the typical anti-inflammatory response is supported through corresponding changes in pro- and anti-inflammatory oxylipins. Limited data indicate that obese compared to normal weight individuals have higher plasma levels of pro-inflammatory oxylipins and other related biomarkers [10,13].
3.2. Matrix
Six studies included in this review used blood samples (serum or plasma) for oxylipin analysis [14,15,16,19,20,22], with two using urine samples [18,21] and one using muscle biopsy samples [17]. More needs to be learned about the influence of the sample matrix on exercise-induced changes in oxylipins. The acute exercise-induced oxylipin response appeared to be comparable across serum, plasma, and muscle sample matrices. Urine samples may be more useful in long-term [53], chronic exercise training studies, with serum and plasma samples preferred for acute exercise studies due to the transient appearance of oxylipins [2,53]. The oxylipin changes following acute resistance exercise in plasma and muscle biopsy samples were somewhat comparable, but more investigation in this area is needed.
3.3. Limitations
Research in the area of exercise and oxylipins is relatively new, and only six studies included in this review included a large number of oxylipins measured with sensitive MS platforms. Thus, the conclusions drawn in this systematic review may change as more studies are published using a wider array of exercise modalities and workloads.
4. Materials and Methods
This study was conducted according to the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) guidelines [54]. The systematized data extraction and studies selection were performed using the free standardized electronic tool State of the Art through Systematic Review (StArt) [55].
4.1. Search Strategy
The electronic search was performed in March 10th, 2020, using the MeSH terms selected: “Humans”, “Exercise”, “Physical Activity”, “Sports”, “Oxylipins”, “Lipid Mediators”, according to the PICOS model (Table 4) [54]. A new search was performed in May 25th, 2020, using the same terms to an update. The articles were retrieved from the databases: PubMed (via National Library of Medicine), Web of Science, SCOPUS (Elsevier), Cochrane, and Science Direct. Moreover, the search strategy was adjusted using the databases features to retrieve only human studies and English language. After the extraction from the databases, the article selection process included these steps: (1) titles and abstracts were independently examined for relevance by two researchers (EFS and CDS); (2) the full text of potentially eligible studies was reviewed. A third independent researcher (CAS) verified the inclusion process in order to solve any disagreement between the two main researchers. References of the included studies were checked for any additional relevant papers. Figure 1 represents the flow diagram of papers through the study selection process.
4.2. Inclusion and Exclusion Criteria
Studies were evaluated according to the listed criteria: (1) human adults and healthy population as the total or part of the sample; (2) analysis of an oxylipins list in serum, plasma, urine or muscle biopsy samples using MS; and (3) exercise as the main factor inducing changes in metabolism. Article were excluded if they focused on only one metabolite, were non-English, did not include an appropriate analysis method, included children, elderly individuals, individuals with any pathology or risk factor (such as hypertension, dyslipidemia, smoking/alcoholism and obesity), included animals only or in vitro models.
4.3. Data Extraction
The following data were extracted from the selected studies using procedures previously reported by our group [52]: name of the first author and year of publication, characteristics of participants and groups (i.e., population, sample size, groups, gender, age, physical activity level), research design elements (i.e., type of research, exercise mode), exercise intensity and duration, enzymatic pathway analyzed, metabololipidomics procedures (i.e., analytical platform, metabolite data), matrix, and summary comments. In addition, the oxylipins determined after physical exercise were summarized in scheme and table. Only data from the placebo or non-dietary intervention groups or phases were used.
4.4. Studies Quality Assessment
5. Conclusions
Recent improvements in mass spectrometry and bioinformatic procedures have advanced scientific understanding of oxylipins, their physiological roles, and the effects of lifestyle interventions including exercise. Exercise-induced oxylipin production during exercise has only recently been described, and the science in this area should advance greatly during the next decade. The number of oxylipins and the magnitude of increase during acute exercise bouts are directly related to the overall workload. The accumulation of oxylipins in plasma is relatively short-lived, with levels returning close to pre-exercise levels within five hours even after about three hours of intense exercise. Although physiological roles during exercise and recovery remain to be determined, data from ancillary studies suggest widespread regulatory effects centered around inflammation and vascular function. The enzyme systems involved with oxylipin generation are complex with multiple regulatory controls, and future research, such as the National Institutes of Health project, ’Molecular Transducers of Physical Activity in Humans Consortium (MoTrPAC) [56], will better define responses to chronic exercise training, moderate- versus high-intensity exercise, muscle damage, sports nutrition, and drug interventions, and varying exercise modalities.
Author Contributions
Acquired financial support, A.M.C.; organized the study, É.F.S., D.C.N., C.A.S. and A.M.C.; created the scoring and classification system, D.C.N.; searched, extracted the studies from the databases and selected for inclusion, É.F.S., C.D.S. and C.A.S.; wrote, É.F.S., D.C.N., C.D.S. and C.A.S.; edited and reviewed the study, D.C.N., É.F.S., C.D.S., C.A.S. and A.M.C. The funders no participated in study design, data collection and analysis, in the decision to publish, or preparation of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
São Paulo Research Foundation—FAPESP (grant #2016/222157 and #2018/250823).
Acknowledgments
The authors would like to acknowledge the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior, Brazil (CAPES, Postgraduate Program in Physiotherapy, grant: 001).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gabbs, M.; Leng, S.; Devassy, J.G.; Monirujjaman, M.; Aukema, H.M. Advances in our understanding of oxylipins derived from dietary PUFAs. Adv. Nutr. 2015, 6, 513–540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markworth, J.F.; Maddipati, K.R.; Cameron-Smith, D. Emerging roles of pro-resolving lipid mediators in immunological and adaptive responses to exercise-induced muscle injury. Exerc. Immunol. Rev. 2016, 22, 110–134. [Google Scholar] [PubMed]
- Spector, A.A.; Kim, H.-Y. Cytochrome P450 epoxygenase pathway of polyunsaturated fatty acid metabolism. Biochim. Biophys. Acta 2015, 1851, 356–365. [Google Scholar] [CrossRef] [Green Version]
- Willenberg, I.; Ostermann, A.I.; Schebb, N.H. Targeted metabolomics of the arachidonic acid cascade: Current state and challenges of LC-MS analysis of oxylipins. Anal. Bioanal. Chem. 2015, 407, 2675–2683. [Google Scholar] [CrossRef] [PubMed]
- Massey, K.A.; Nicolaou, A. Lipidomics of polyunsaturated-fatty-acid-derived oxygenated metabolites. Biochem. Soc. Trans. 2011, 39, 1240–1246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shearer, G.C.; Walker, R.E. An overview of the biologic effects of omega-6 oxylipins in humans. Prostaglandins Leukot. Essent. Fatty Acids 2018, 137, 26–38. [Google Scholar] [CrossRef]
- Lynes, M.D.; Leiria, L.O.; Lundh, M.; Bartelt, A.; Shamsi, F.; Huang, T.L.; Takahashi, H.; Hirshman, M.F.; Schlein, C.; Lee, A.; et al. The cold-induced lipokine 12,13-diHOME promotes fatty acid transport into brown adipose tissue. Nat. Med. 2017, 23, 631–637. [Google Scholar] [CrossRef] [Green Version]
- Stark, K.D.; Van Elswyk, M.E.; Higgins, M.R.; Weatherford, C.A.; Salem, N. Global survey of the omega-3 fatty acids, docosahexaenoic acid and eicosapentaenoic acid in the blood stream of healthy adults. Prog. Lipid Res. 2016, 63, 132–152. [Google Scholar] [CrossRef]
- Zimmer, B.; Angioni, C.; Osthues, T.; Toewe, A.; Thomas, D.; Pierre, S.C.; Geisslinger, G.; Scholich, K.; Sisignano, M. The oxidized linoleic acid metabolite 12,13-DiHOME mediates thermal hyperalgesia during inflammatory pain. Biochim. Biophys. Acta Mol. Cell. Biol. Lipids 2018, 1863, 669–678. [Google Scholar] [CrossRef]
- Nieman, D.C.; Mitmesser, S.H. Potential impact of nutrition on immune system recovery from heavy exertion: A metabolomics perspective. Nutrients 2017, 9, 513. [Google Scholar] [CrossRef] [Green Version]
- Nieman, D.C.; Lila, M.A.; Gillitt, N.D. Immunometabolism: A multi-omics approach to interpreting the influence of exercise and diet on the immune system. Annu. Rev. Food Sci. Technol. 2019, 10, 341–363. [Google Scholar] [CrossRef] [PubMed]
- Nieman, D.C.; Pence, B.D. Exercise immunology: Future directions. J. Sport Health Sci. 2020, 1–14. [Google Scholar] [CrossRef]
- Nieman, D.C.; Wentz, L.M. The compelling link between physical activity and the body’s defense system. J. Sport Health Sci. 2019, 8, 201–217. [Google Scholar] [CrossRef]
- Nieman, D.C.; Gillitt, N.D.; Chen, G.-Y.; Zhang, Q.; Sakaguchi, C.A.; Stephan, E.H. Carbohydrate intake attenuates post-exercise plasma levels of cytochrome P450-generated oxylipins. PLoS ONE 2019, 14, e0213676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nieman, D.; Gillitt, N.; Chen, G.-Y.; Zhang, Q.; Sha, W.; Kay, C.D.; Chandra, P.; Kay, K.L.; Lila, M.A. Blueberry and banana consumption mitigate arachidonic, cytochrome P450 oxylipin generation during recovery from 75-Km cycling: A randomized trial. Front. Nutr. (in press). [CrossRef]
- Markworth, J.F.; Vella, L.; Lingard, B.S.; Tull, D.L.; Rupasinghe, T.W.; Sinclair, A.J.; Maddipati, K.R.; Cameron-Smith, D. Human inflammatory and resolving lipid mediator responses to resistance exercise and ibuprofen treatment. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 305, R1281–R1296. [Google Scholar] [CrossRef] [Green Version]
- Vella, L.; Markworth, J.F.; Farnfield, M.M.; Maddipati, K.R.; Russell, A.P.; Cameron-Smith, D. Intramuscular inflammatory and resolving lipid profile responses to an acute bout of resistance exercise in men. Physiol. Rep. 2019, 7, e14108. [Google Scholar] [CrossRef]
- García-Flores, L.A.; Medina, S.; Gómez, C.; Wheelock, C.E.; Cejuela, R.; Martínez-Sanz, J.M.; Oger, C.; Galano, J.-M.; Durand, T.; Hernández-Sáez, Á.; et al. Aronia-citrus juice (polyphenol-rich juice) intake and elite triathlon training: A lipidomic approach using representative oxylipins in urine. Food Funct. 2018, 9, 463–475. [Google Scholar] [CrossRef] [Green Version]
- Gollasch, B.; Dogan, I.; Rothe, M.; Gollasch, M.; Luft, F.C. Maximal exercise and plasma cytochrome P450 and lipoxygenase mediators: A lipidomics study. Physiol. Rep. 2019, 7, e14165. [Google Scholar] [CrossRef] [Green Version]
- Giordano, R.M.; Newman, J.W.; Pedersen, T.L.; Ramos, M.I.; Stebbins, C.L. Effects of dynamic exercise on plasma arachidonic acid epoxides and diols in human volunteers. Int. J. Sport Nutr. Exerc. Metab. 2011, 21, 471–479. [Google Scholar] [CrossRef]
- Medina, S.; Domínguez-Perles, R.; Cejuela-Anta, R.; Villaño, D.; Martínez-Sanz, J.M.; Gil, P.; García-Viguera, C.; Ferreres, F.; Gil, J.I.; Gil-Izquierdo, A. Assessment of oxidative stress markers and prostaglandins after chronic training of triathletes. Prostaglandins Other Lipid Mediat. 2012, 99, 79–86. [Google Scholar] [CrossRef]
- Nieman, D.C.; Shanely, R.A.; Luo, B.; Meaney, M.P.; Dew, D.A.; Pappan, K.L. Metabolomics approach to assessing plasma 13- and 9-hydroxy-octadecadienoic acid and linoleic acid metabolite responses to 75-km cycling. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2014, 307, R68–R74. [Google Scholar] [CrossRef] [Green Version]
- Shoieb, S.M.; El-Sherbeni, A.A.; El-Kadi, A.O. Subterminal hydroxyeicosatetraenoic acids: Crucial lipid mediators in normal physiology and disease states. Chem. Biol. Interact. 2018, 299, 140–150. [Google Scholar] [CrossRef] [PubMed]
- Powell, W.S.; Rokach, J. Biosynthesis, biological effects, and receptors of hydroxyeicosatetraenoic acids (HETEs) and oxoeicosatetraenoic acids (oxo-ETEs) derived from arachidonic acid. Biochim. Biophys. Acta 2015, 1851, 340–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubois, R.N.; Abramson, S.B.; Crofford, L.; Gupta, R.A.; Simon, L.S.; Van De Putte, L.B.; Lipsky, P.E. Cyclooxygenase in biology and disease. FASEB J. 1998, 12, 1063–1073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serhan, C.N.; Dalli, J.; Colas, R.A.; Winkler, J.W.; Chiang, N. Protectins and maresins: New pro-resolving families of mediators in acute inflammation and resolution bioactive metabolome. Biochim. Biophys. Acta 2015, 1851, 397–413. [Google Scholar] [CrossRef] [Green Version]
- Murakami, M.; Yamamoto, K.; Miki, Y.; Murase, R.; Sato, H.; Taketomi, Y. The roles of the secreted phospholipase A2 gene family in immunology. Adv. Immunol. 2016, 132, 91–134. [Google Scholar]
- Sato, H.; Taketomi, Y.; Murakami, M. Metabolic regulation by secreted phospholipase A2. Inflamm. Regen. 2016, 36, 7. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, M.; Tucker, D.E.; Burchett, S.A.; Leslie, C.C. Properties of the Group IV phospholipase A2 family. Prog. Lipid Res. 2006, 45, 487–510. [Google Scholar] [CrossRef]
- Leslie, C.C. Cytosolic phospholipase A2: Physiological function and role in disease. J. Lipid Res. 2015, 56, 1386–1402. [Google Scholar] [CrossRef] [Green Version]
- Guijas, C.; Rodríguez, J.P.; Rubio, J.M.; Balboa, M.A.; Balsinde, J. Phospholipase A2 regulation of lipid droplet formation. Biochim. Biophys. Acta (BBA)—Molec. Cell Biol. Lipids 2014, 1841, 1661–1671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kramer, H.F.; Goodyear, L.J. Exercise, MAPK, and NF-κB signaling in skeletal muscle. J. Appl. Physiol. 2007, 103, 388–395. [Google Scholar] [CrossRef] [PubMed]
- Son, Y.; Kim, S.; Chung, H.-T.; Pae, H.-O. Reactive oxygen species in the activation of MAP kinases. Meth. Enzymol. 2013, 528, 27–48. [Google Scholar] [PubMed]
- Suzuki, K. Cytokine response to exercise and its modulation. Antioxidants 2018, 7, 17. [Google Scholar] [CrossRef] [Green Version]
- Serhan, C.N.; Levy, B.D. Resolvins in inflammation: Emergence of the pro-resolving superfamily of mediators. J. Clin. Invest. 2018, 128, 2657–2669. [Google Scholar] [CrossRef]
- Stanford, K.I.; Lynes, M.D.; Takahashi, H.; Baer, L.A.; Arts, P.J.; May, F.J.; Lehnig, A.C.; Middelbeek, R.J.W.; Richard, J.J.; So, K.; et al. 12,13-diHOME: An exercise-induced lipokine that increases skeletal muscle fatty acid uptake. Cell Metab. 2018, 27, 1111–1120.e3. [Google Scholar] [CrossRef] [Green Version]
- Demers, L.M.; Harrison, T.S.; Halbert, D.R.; Santen, R.J. Effect of prolonged exercise on plasma prostaglandin levels. Prostaglandins Med. 1981, 6, 413–418. [Google Scholar] [CrossRef]
- Karamouzis, M.; Karamouzis, I.; Vamvakoudis, E.; Ampatzidis, G.; Christoulas, K.; Angelopoulou, N.; Mandroukas, K. The response of muscle interstitial prostaglandin E(2)(PGE(2)), prostacyclin I(2)(PGI(2)) and thromboxane A(2)(TXA(2)) levels during incremental dynamic exercise in humans determined by in vivo microdialysis. Prostaglandins Leukot. Essent. Fatty Acids 2001, 64, 259–263. [Google Scholar] [CrossRef]
- Nowak, J.; Wennmalm, Å. Effect of exercise on human arterial and regional venous plasma concentrations of prostaglandin E. Prostaglandins Med. 1978, 1, 489–497. [Google Scholar] [CrossRef]
- Kaley, G.; Weiner, R. Prostaglandin E1: A potential mediator of the inflammatory response. Ann. N. Y. Acad. Sci. 1971, 180, 338–350. [Google Scholar] [CrossRef]
- Carroll, C.C.; O’Connor, D.T.; Steinmeyer, R.; Del Mundo, J.D.; McMullan, D.R.; Whitt, J.A.; Ramos, J.E.; Gonzales, R.J. The influence of acute resistance exercise on cyclooxygenase-1 and -2 activity and protein levels in human skeletal muscle. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 305, R24–R30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markworth, J.F.; Cameron-Smith, D. Prostaglandin F2α stimulates PI3K/ERK/mTOR signaling and skeletal myotube hypertrophy. Am. J. Physiol. Cell Physiol. 2011, 300, C671–C682. [Google Scholar] [CrossRef] [PubMed]
- Peake, J.M.; Suzuki, K.; Hordern, M.; Wilson, G.; Nosaka, K.; Coombes, J.S. Plasma cytokine changes in relation to exercise intensity and muscle damage. Eur. J. Appl. Physiol. 2005, 95, 514–521. [Google Scholar] [CrossRef] [PubMed]
- Ackermann, J.A.; Hofheinz, K.; Zaiss, M.M.; Krönke, G. The double-edged role of 12/15-lipoxygenase during inflammation and immunity. Biochim. Biophys. Acta Mol. Cell. Biol. Lipids 2017, 1862, 371–381. [Google Scholar] [CrossRef]
- Savas, Ü.; Wei, S.; Hsu, M.-H.; Falck, J.R.; Guengerich, F.P.; Capdevila, J.H.; Johnson, E.F. 20-Hydroxyeicosatetraenoic Acid (HETE)-dependent Hypertension in Human Cytochrome P450 (CYP) 4A11 transgenic mice: Normalization of blood pressure by sodium restriction, hydrochlorothiazide, or blockade of the type 1 angiotensin II receptor. J. Biol. Chem. 2016, 291, 16904–16919. [Google Scholar] [CrossRef] [Green Version]
- Kumar, N.; Gupta, G.; Anilkumar, K.; Fatima, N.; Karnati, R.; Reddy, G.V.; Giri, P.V.; Reddanna, P. 15-Lipoxygenase metabolites of α-linolenic acid, (13-(S)-HPOTrE and 13-(S)-HOTrE), mediate anti-inflammatory effects by inactivating NLRP3 inflammasome. Sci. Rep. 2016, 6, 1–14. [Google Scholar] [CrossRef]
- Fischer, R.; Konkel, A.; Mehling, H.; Blossey, K.; Gapelyuk, A.; Wessel, N.; von Schacky, C.; Dechend, R.; Muller, D.N.; Rothe, M.; et al. Dietary omega-3 fatty acids modulate the eicosanoid profile in man primarily via the CYP-epoxygenase pathway. J. Lipid Res. 2014, 55, 1150–1164. [Google Scholar] [CrossRef] [Green Version]
- VanRollins, M. Epoxygenase metabolites of docosahexaenoic and eicosapentaenoic acids inhibit platelet aggregation at concentrations below those affecting thromboxane synthesis. J. Pharmacol. Exp. Ther. 1995, 274, 798–804. [Google Scholar]
- Kim, N.; Ramon, S.; Thatcher, T.H.; Woeller, C.F.; Sime, P.J.; Phipps, R.P. Specialized pro-resolving mediators (SPMs) inhibit human B-cell IgE production. Eur. J. Immunol. 2016, 46, 81–91. [Google Scholar] [CrossRef]
- Schwab, J.M.; Chiang, N.; Arita, M.; Serhan, C.N. Resolvin E1 and protectin D1 activate inflammation-resolution programmes. Nature 2007, 447, 869–874. [Google Scholar] [CrossRef] [Green Version]
- Bannenberg, G.L.; Chiang, N.; Ariel, A.; Arita, M.; Tjonahen, E.; Gotlinger, K.H.; Hong, S.; Serhan, C.N. Molecular circuits of resolution: Formation and actions of resolvins and protectins. J. Immunol. 2005, 174, 4345–4355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakaguchi, C.A.; Nieman, D.C.; Signini, E.F.; Abreu, R.M.; Catai, A.M. Metabolomics-based studies assessing exercise-induced alterations of the human metabolome: A systematic review. Metabolites 2019, 9, 164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolfer, A.M.; Gaudin, M.; Taylor-Robinson, S.D.; Holmes, E.; Nicholson, J.K. Development and validation of a high-throughput ultrahigh-performance liquid chromatography—Mass spectrometry approach for screening of oxylipins and their precursors. Anal. Chem. 2015, 87, 11721–11731. [Google Scholar] [CrossRef] [PubMed]
- Liberati, A.; Altman, D.G.; Tetzlaff, J.; Mulrow, C.; Gøtzsche, P.C.; Ioannidis, J.P.A.; Clarke, M.; Devereaux, P.J.; Kleijnen, J.; Moher, D. The PRISMA statement for reporting systematic reviews and meta-analyses of studies that evaluate health care interventions: Explanation and elaboration. PLoS Med. 2009, 6, e1000100. [Google Scholar] [CrossRef] [PubMed]
- Fabbri, S.; Silva, C.; Hernandes, E.; Octaviano, F.; Di Thommazo, A.; Belgamo, A. Improvements in the StArt tool to better support the systematic review process. In Proceedings of the 20th International Conference on Evaluation and Assessment in Software Engineering, Limerick, Ireland, 1–3 June 2016; ACM: New York, NY, USA, 2016. [Google Scholar]
- Sparks, L.M. Exercise training response heterogeneity: Physiological and molecular insights. Diabetologia 2017, 60, 2329–2336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Outcomes of a review flow diagram.

Figure 2.
Physical exercise-related lipid pathways and metabolites. Metabolites inside the boxes are those discussed in the articles, while those outside are not discussed in the articles but are involved in the pathways. Underlined metabolites are those from urine samples. Italicized metabolites are those from muscle biopsy samples. PUFAs: polyunsaturated fatty acids; AA: aracdonic acid; AdA: adrenic acid; ALA: α-linolenic acid; 20-COOH-AA: 20-carboxy arachidonic acid; 20-COOH-LTB4: 20-carboxy-leukotriene B4; COX: cyclooxygenase; CYP: cytochrome P450; D12-PGJ3: Delta-12-prostaglandin J3; 15d-D12,14-PGJ3: 15-deoxy-Delta-12,14-prostaglandin J3; DGLA: dihomo-γ-linolenic acid; dh-PG: dihomo-prostaglandin; DHA: docosahexaenoic acid; 13,14-dihydro-15-keto-PGE2: 13,14-dihydro-15-keto prostaglandin E2; 11-dihydro-TXB2: 11-dihydro thromboxane B2; 2,3-dinoβ-11-PGF2α: 2,3-dinor-11β-prostaglandin F2α; DiHDoHE: dihydroxy-docosahexaenoic acid; DiHDPE: dihydroxy-docosapentaenoic acid; DiHETE: dihydroxy-eicosatetraenoic acid; DiHETrE: dihydroxy-eicosatrienoic acid; DiHOME: dihydroxy-octadecenoic acid; EPA: eicosapentaenoic acid; EpDPE: epoxy-docosapentaenoic acid; EpETE: epoxy-eicosatetraenoic acid; EpETrE: epoxy-eicosatrienoic acid; EpOME: epoxy-octadecenoic acid; HDoHE: hydroxy-docosahexaenoic acid; HEPE: hydroxy-eicosapentaenoic acid; HETE: hydroxy-eicosatetraenoic acid; HETrE: hydroxy-eicosatrienoic acid; HHTrE: hydroxyheptadecatrienoic acid; HODE: hydroxy-octadecadienoic acid; HOTrE: hydroxy-octadecatrienoic acid; HpDoHE: hydroperoxy-docosahexaenoic acid; HpEPE: hydroperoxy-eicosapentaenoic acid; HpETE: hydroperoxy-eicosatetraenoic acid; HpETrE: hydroperoxy-eicosatrienoic acid; HpODE: hydroperoxy-octadecadienoic acid; HpOTrE: hydroperoxy-octadecatrienoic acid; Iso: isoprostane; 15-keto-PGE2: 15-keto prostaglandin E2; 6-keto-PGF1α: 6-keto prostaglandin F1α; 15-keto-PGF2α: 15-keto prostaglandin F2α; LA: linoleic acid; LOX: lipoxygenase; LT: leukotriene; LX: lipoxin; oxo-ETE: oxo-eicosatetraenoic acid; 12-oxo-LTB4: 12-oxo leukotriene B4; oxo-ODE: oxo-octadecadienoic acid; PD1: protectin D1; PG: prostaglandin; PGDM: prostaglandin D metabolite; 11-β-PGF2α: 11β-prostaglandin F2α; PGFM: prostaglandin F metabolite; PLA2: phospolipase A2; 15R-HpETE: 15R-hydroperoxy-eicosatetraenoic acid; 15R-LXA4: 15R-lipoxin A4; Rv: resolvina; Tetranor-PGDM: tetranor-prostaglandin D metabolite; tetranor-PGEM: tetranor-prostaglandin E metabolite; TX: thromboxane.
Figure 2.
Physical exercise-related lipid pathways and metabolites. Metabolites inside the boxes are those discussed in the articles, while those outside are not discussed in the articles but are involved in the pathways. Underlined metabolites are those from urine samples. Italicized metabolites are those from muscle biopsy samples. PUFAs: polyunsaturated fatty acids; AA: aracdonic acid; AdA: adrenic acid; ALA: α-linolenic acid; 20-COOH-AA: 20-carboxy arachidonic acid; 20-COOH-LTB4: 20-carboxy-leukotriene B4; COX: cyclooxygenase; CYP: cytochrome P450; D12-PGJ3: Delta-12-prostaglandin J3; 15d-D12,14-PGJ3: 15-deoxy-Delta-12,14-prostaglandin J3; DGLA: dihomo-γ-linolenic acid; dh-PG: dihomo-prostaglandin; DHA: docosahexaenoic acid; 13,14-dihydro-15-keto-PGE2: 13,14-dihydro-15-keto prostaglandin E2; 11-dihydro-TXB2: 11-dihydro thromboxane B2; 2,3-dinoβ-11-PGF2α: 2,3-dinor-11β-prostaglandin F2α; DiHDoHE: dihydroxy-docosahexaenoic acid; DiHDPE: dihydroxy-docosapentaenoic acid; DiHETE: dihydroxy-eicosatetraenoic acid; DiHETrE: dihydroxy-eicosatrienoic acid; DiHOME: dihydroxy-octadecenoic acid; EPA: eicosapentaenoic acid; EpDPE: epoxy-docosapentaenoic acid; EpETE: epoxy-eicosatetraenoic acid; EpETrE: epoxy-eicosatrienoic acid; EpOME: epoxy-octadecenoic acid; HDoHE: hydroxy-docosahexaenoic acid; HEPE: hydroxy-eicosapentaenoic acid; HETE: hydroxy-eicosatetraenoic acid; HETrE: hydroxy-eicosatrienoic acid; HHTrE: hydroxyheptadecatrienoic acid; HODE: hydroxy-octadecadienoic acid; HOTrE: hydroxy-octadecatrienoic acid; HpDoHE: hydroperoxy-docosahexaenoic acid; HpEPE: hydroperoxy-eicosapentaenoic acid; HpETE: hydroperoxy-eicosatetraenoic acid; HpETrE: hydroperoxy-eicosatrienoic acid; HpODE: hydroperoxy-octadecadienoic acid; HpOTrE: hydroperoxy-octadecatrienoic acid; Iso: isoprostane; 15-keto-PGE2: 15-keto prostaglandin E2; 6-keto-PGF1α: 6-keto prostaglandin F1α; 15-keto-PGF2α: 15-keto prostaglandin F2α; LA: linoleic acid; LOX: lipoxygenase; LT: leukotriene; LX: lipoxin; oxo-ETE: oxo-eicosatetraenoic acid; 12-oxo-LTB4: 12-oxo leukotriene B4; oxo-ODE: oxo-octadecadienoic acid; PD1: protectin D1; PG: prostaglandin; PGDM: prostaglandin D metabolite; 11-β-PGF2α: 11β-prostaglandin F2α; PGFM: prostaglandin F metabolite; PLA2: phospolipase A2; 15R-HpETE: 15R-hydroperoxy-eicosatetraenoic acid; 15R-LXA4: 15R-lipoxin A4; Rv: resolvina; Tetranor-PGDM: tetranor-prostaglandin D metabolite; tetranor-PGEM: tetranor-prostaglandin E metabolite; TX: thromboxane.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Study classifications according to the score system in Table 5.
| Investigators, Year Published | Research Design | Methodology | Novelty | Final Score | Classification | ||
|---|---|---|---|---|---|---|---|
| Subjects Number | Studies Characteristics | Analysis Methods | Statistical Support | ||||
| Nieman et al. (2019) [14] | 2 | 2 | 3 | 2 | 2 | 11 | Excellent |
| Nieman et al. (2020) [15] | 0 | 2 | 3 | 2 | 2 | 9 | Excellent |
| Garcia-Flores et al. (2018) [18] | 2 | 2 | 3 | 0 | 1 | 8 | Good |
| Markworth et al. (2013) [16] | 0 | 2 | 3 | 1 | 2 | 8 | Good |
| Vella et al. (2019) [17] | 0 | 1 | 3 | 0 | 2 | 6 | Good |
| Gollach et al. (2019) [19] | 0 | 1 | 3 | 0 | 1 | 5 | Fair |
| Nieman et al.(2014) [22] | 0 | 1 | 1 | 0 | 1 | 3 | Poor |
| Giordano et al. (2011) [20] | 0 | 1 | 1 | 0 | 1 | 3 | Poor |
| Medina et al. (2012) [21] | 0 | 0 | 1 | 0 | 1 | 2 | Poor |
Table 2.
Study characteristics.
| Investigators, Year Published | Study Population | Research Design | Exercise Intensity and Duration | Enzymatic Pathway | Analytical Platform | Matrix | Key Findings, Exercise Effect |
|---|---|---|---|---|---|---|---|
| Garcia-Flores et al. (2018) [18] | 16 triathletes (10 men 19.0 ± 1.7 years, and 6 women 21.1 ± 3.0 years of age) | Randomized, double-blinded, placebo-controlled, and crossover design. Triathlon training at different conditions: control baseline (15 days), control training (15 days), placebo, and supplement drink crossover (100 days) and washout between these conditions (10 days), and control post-training (15 days). Training was based on objective load scale (ECOs). Urine samples timepoints: pre- and post-each condition (24 h). | Training, high-intensity, long-duration | COX, LOX, and non- enzymatic pathways | UHPLC-MS/MS | Urine | 37 oxylipins detected, with small decreases in F2-IsoPs and PGF1α, and small increases in PGDM, 11-β-PGF2α and PGE1 |
| Medina et al. (2012) [21] | 15 triathletes (10 men 19.0 ± 1.7 years and 5 women 21.8 ± 3.0 years of age) | Intense triathlon training for two weeks (cycling, swimming, running). No control group. Urine samples timepoints: pre-training (24 h) and post-training (24 h). | Training, high-intensity, long-duration | COX and non- enzymatic pathways | UPLC–QqQ-MS/MS | Urine | 13 oxylipins detected, with small decreases in F2-IsoPs, tetranor-PGEM and 11-β-PGF2α, and an increase in 6-keto-PGF1α |
| Giordano et al. (2011) [20] | 14 light to moderately active healthy subjects (6 men and 8 women, 36.9 ± 8.4 years of age) | In three visits, participants randomly performed submaximal bicycle tests for 20 min at 30%, 60%, and 80% of their maximal workload. In an additional visit, a test was performed at 60% of maximal work capacity for 40 min. Blood samples timepoints: pre-exercise, during exercise (20th min), and post-exercise (2 min) in the three visits, and pre-exercise and during exercise (39th min) in the additional visit | Low, moderate, and high-intensity, short-duration | CYP | UPLC-MS/MS | Plasma | 6 oxylipins detected, with small increases after 80% exercise in 8,9-DiHETrE, 11,12-DiHETrE, 14,15-DiHETrE, and after 40 min 60% exercise for 14,15-EpETrE and 14,15-DiHETrE |
| Gollasch et al. (2019) [19] | Six healthy subjects (5 men and 1 woman 38.0 ± 15.0 years of age) | Subjects performed a maximal graded treadmill test. Blood samples timepoints: pre-exercise test (10 min before), during exercise (when heart rate reached 150 bpm), post-exercise (0 min, 10 min) | High-intensity, short-duration | LOX and CYP | HPLC-MS/MS | Plasma | 56 oxylipins detected, with small increases in 5,6-DiHETrE, 12,13-EpOME, 5,6-DiHETE, 17,18-DiHETE. Most of them returned to close pre-exercise values 10 min after the end exercise |
| Markworth et al. (2013) [16] | 16 healthy men (2 groups of 8 each, 23 ± 1.3 years and 23.0 ± 0.5 years of age) | Parallel group design randomized with two groups: placebo and ibuprofen. Exercise included a 10 min warmup; 3 sets of 8 –10 repetitions of squatting with bar; leg press 45° and knee extension at 80% 1 RM. Circuit with 1 min of recovery between sets, 3 min recovery between stations. Blood samples timepoints: after 10 h fasting; pre-exercise (15 min), post-exercise protocol (0 h, 0.5 h, 1 h, 1.5 h, 2 h, 2.5 h, 3 h, 24 h) | High-intensity resistance, short-duration | COX, LOX, and CYP | HPLC-MRM-MS/MS | Serum | 87 oxylipins detected with small to moderate increases in 29 oxylipins, especially between 1 and 3 h post-exercise. Most of them returned to close pre-exercise values between 3 and 24 h after the end exercise |
| Nieman et al. (2020) [15] | 59 healthy cyclists (38.6 ± 1.5 years of age) | Parallel group design, randomized, double blind and placebo-controlled intervention. Two-week supplementation period (freeze-dried blueberry powder or placebo) followed by a 75 km cycling time trial (while consuming water only or water with bananas). Four groups (blueberry-water trial / blueberry–banana trial / placebo-banana trial / placebo-water trial). Blood samples timepoints: pre-exercise, post-exercise (0 h, 1.5 h, 3 h, 5 h, 24 h, and 48 h) | High-intensity, long-duration | COX, LOX, and CYP | LC-MRM-MS | Plasma | Large increases in plasma concentrations of 64 of 67 oxylipins detected, with most near pre-exercise levels within 5 h post-exercise |
| Nieman et al. (2019) [14] | 20 healthy cyclists (39.1 ± 2.4 years of age) | Randomized, crossover, counterbalanced approach. Four sessions of 75-km cycling time trial, 2-weekwashout. Four groups (Cavendish banana trial / mini-yellow banana trial / sugar beverage trial / water trial). Blood samples timepoints: pre-exercise, post-exercise (0 h, 0.75 h, 1.5 h, 3 h, 4.5 h, 21 h and 45 h) | High-intensity, long-duration | COX, LOX, and CYP | LC-MRM-MS/UHPLC/MS | Plasma | Large increases in plasma concentrations of 43 of 45 oxylipins detected, with most near pre-exercise levels within 4.5 h post-exercise |
| Nieman et al., (2014) [22] | 19 male cyclists (38.1 ± 1.6 years of age) | Subjects performed a 75 km cycling time trial without any beverage or food containing energy or nutrients. Blood samples timepoints: pre-exercise and post-exercise (0 h, 1.5 h, 21 h) | High-intensity, long-duration | LOX and CYP | UHPLC- MS/MS and GC-MS | Plasma | Large increases in 9-HODE, 13-HODE, 9,10-DiHOME, and 12,13-DiHOME, with most near pre-exercise levels between 1.5 and 21 h post-exercise |
| Vella et al. (2019) [17] | 12 recreationally active men (22.1 ± 0.6 years of age) | Acute bout of maximal concentric and eccentric isokinetic unilateral knee extension exercise, three sets of 12 maximal repetitions, 2 min of rest between sets. Muscle biopsy timepoints: pre-protocol, post-protocol (2 h, 4 h, 24 h) | High-intensity, resistance, short-duration | COX, LOX, and CYP | HPLC-MRM-MS/MS | Muscle Biopsy | 84 oxylipins detected. Modest to small increases in 22 oxylipins at 2 h post-exercise including TXB2, PGE2, PGF2α, 15d-D12,14-PGJ3), 12-oxo-LTB4, 20-COOH-LTB4, 5-HETE, 12-HETE, tetranor 12-HETE, 15-HETE, 12-HEPE, 4HDoHE, 7-HDoHE, 14-HDoHE, 5,6-EpETrE, 11,12-DiHETrE, and 14,15-DiHETrE. Most of them returned to close pre-exercise values 4 h after the end exercise |
20-COOH-LTB4: 20-carboxy-leukotriene B4; COX: cyclooxygenase; CYP: cytochrome P450; 11-dihydro-TXB2: 11-dehydro Thromboxane B2; DiHETE: dihydroxy-eicosatetraenoic acid; DiHETrE: dihydroxy-eicosatrienoic acid; DiHOME: dihydroxy-octadecenoic acid; 15d-D12,14-PGJ3: 15-Deoxy-D12,14-prostaglandin J3; EpETrE: epoxy-eicosatrienoic acid; EpOME: epoxy-octadecenoic acid; GC-MS: gas chromatography mass spectrometry; HDoHE: hydroxy-docosahexaenoic acid; HEPE: hydroxy-eicosapentaenoic acid; HETE: hydroxy-eicosatetraenoic acid; HODE: hydroxy-octadecadienoic acid; HPLC-MRM-MS/MS: high-performance liquid chromatography-multiple reaction monitoring-mass spectrometry-tandem mass spectrometer; HPLC-MS/MS: high-performance liquid chromatography-tandem mass spectrometer; IsoPs: isoprostanes; 6-keto-PGF1α: 6-keto Prostaglandin F1α; LC-MS/MS: liquid chromatography coupled to mass spectrometry; LC-MRM-MS/UHPLC: liquid chromatography-multiple reaction monitoring-mass spectrometry with ultra-high performance liquid chromatography; LOX: lipoxygenase; LT: leukotriene; 12-oxo-LTB4: 12-oxo Leukotriene B4; PG: prostaglandin; PGDM: prostaglandin D metabolite; 11-β-PGF2α: 11β-Prostaglandin F2α; tetranor-PGDM: tetranor-prostaglandin D metabolite; tetranor-PGEM: tetranor-prostaglandin E metabolite; TX: thromboxane; UHPLC-MS/MS: ultra-high performance liquid chromatography coupled to mass spectrometry; UPLC–QqQ-MS/MS: ultra-performance liquid chromatography coupled to triple quadrupole mass spectrometry; UPLC-MS/MS: ultra-performance liquid chromatography -tandem mass spectrometer.
Table 3.
Exercise-related oxylipins measured and magnitude of changes reported in each study.
| Pathway | Studies | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Giordano et al. (2011) [20] | Medina et al. (2012) [21] | Markworth et al. (2013) [16] | Nieman et al. (2014) [22] | Garcia-Flores et al. (2018) [18] | Nieman et al. (2019) [14] | Gollach et al. (2019) [19] | Vella et al. (2019) [17] | Nieman et al. (2020) [15] | |
| Acute Effect (Short Duration) | Chronic Effect (Long Duration) | Acute Effect (Short Duration) | Acute Effect (Long Duration) | Chronic Effect (long Duration) | Acute Effect (Long Duration) | Acute Effect (Short Duration) | Acute Effect (Short Duration) | Acute Effect (long Duration) | |
| COX | - | ↓ Tetranor- PGEM ↓ 11-β-PGF2α ↑ 6-keto-PGF1α | ↑↑ TXB2 ↑↑ 12-HHTrE ↑ PGD2 ↑ PGE2 ↑ 15-keto-PGE2 ↑ 15-keto- PGF2α ↑ 6-keto-PGF1α ↑ 13,14- dihydro-15-keto-PGE2 ↑ RvE1 | - | ↑ PGDM ↑ 11-β-PGF2α ↑ PGE1 ↓↓ PGF1α |
↑↑ TXB2 ↑↑ 12-HHTrE ↑ PGFM ↑↑ 18-HEPE | - | ↑ TXB2 ↑ 12-HHTrE ↑ PGE2 ↑ PGF2α ↑ 15d- D12,14-PGJ3 | ↑↑TXB2 ↑↑ 12-HHTrE ↑ PGFM ↑↑ PGE2 ↑↑ dh-PGE2 ↑↑ TXB1 ↑↑ TXB3 ↑ PGE3 ↑ 18-HEPE ↑ RvE1 |
| LOX | - | - | ↑ 12-HETE ↑ 5,12- DiHETE ↑ Tetranor- 12-HETE ↑ 15-HETE ↑ 15-oxo-ETE ↑ LTB4 ↑ LXA4 ↑ LXB4 ↑ 9-oxo-ODE ↑ 13-HODE ↑ 13-oxo-ODE ↑ 10(S),17(S)- DiHDoHE ↑ RvD1 | ↑ 9-HODE ↑ 13-HODE | ↑ 5-HETE ↑ 8-HETE ↑↑ 9-HETE ↑↑ 11-HETE ↑↑ 12-HETE ↑↑ Tetranor 12-HETE ↑↑ 15-HETE ↑↑ 9-HODE ↑↑ 9-oxo-ODE ↑↑ 13-HODE ↑ 13-oxo-ODE ↑↑ 5-HETrE ↑↑ 8-HETrE ↑↑ 15-HETrE ↑↑ 4-HdoHE ↑↑ 8-HdoHE ↑↑ 10-HdoHE ↑ 13-HdoHE ↑↑ 14-HdoHE ↑ 16-HdoHE ↑↑ 5-HEPE ↑↑ 12-HEPE ↑ 15-HEPE | - | ↑ 5-HETE ↑ 12-HETE ↑↑ Tetranor 12-HETE ↑ 12-oxo- LTB4 ↑ 20-COOH- LTB4 ↑ 4-HdoHE ↑ 7-HdoHE ↑ 14-HdoHE ↑↑ 12-HEPE | ↑ 5-HETE ↑ 5-oxo-ETE ↑↑ 5,15- DiHETE ↑↑ 8-HETE ↑↑ 9-HETE ↑↑ 11-HETE ↑↑ 12-HETE ↑ 12-oxo-ETE ↑↑ Tetranor 12-HETE ↑ 15-HETE ↑ 15R-LXA4 ↑↑ 9-HODE ↑↑ 9-oxo-ODE ↑↑ 13-HODE ↑↑ 13- oxo-ODE ↑↑ 5-HETrE ↑↑ 8-HETrE ↑↑ 15-HETrE ↑ 4-HdoHE ↑↑ 7-HdoHE ↑↑ 8-HdoHE ↑↑ 10-HdoHE ↑↑ 11-HDoHE | |
| LOX | - | - | ↑↑ 9-HOTrE ↑↑ 13-HOTrE | - | ↑↑ 13-HDoHE ↑↑ 14-HDoHE ↑ 16-HDoHE ↑↑ 17-HDoHE ↑↑ PD1 ↑↑ 5-HEPE ↑ 8-HEPE ↑↑ 9-HEPE ↑ 11-HEPE ↑↑ 12-HEPE ↑↑ 15-HEPE ↑↑ 9-HOTrE ↑↑ 13-HOTrE | ||||
| CYP | ↑ 8,9- DiHETrE ↑ 11,12- DiHETrE ↓ 14,15- EpETrE ↑ 14,15- DiHETrE | - | ↑ 11,12- DiHETrE ↑ 14,15- DiHETrE ↑ 9,10-EpOME ↑ 9,10- DiHOME | ↑ 9,10- DiHOME ↑ 12,13- DiHOME | - | ↑↑ 8,9- DiHETrE ↑↑ 11,12- DiHETrE ↑ 14,15- DiHETrE ↑↑ 17-HETE ↑ 18-HETE ↑ 19-HETE ↑↑ 20-HETE ↑↑ 20- COOH-AA ↑ 9,10- EpOME ↑↑ 9,10- DiHOME ↑↑ 12,13- DiHOME ↑↑ 19,20- DiHDPE ↑↑ 20- HDoHE | ↑ 5,6- DiHETrE ↑ 12,13- EpOME ↑ 5,6- DiHETE ↑ 17,18- DiHETE | ↑ 5,6-EpETrE ↑ 11,12- DiHETrE ↑ 14,15- DiHETrE | ↑↑ 5,6- EpETrE ↑↑ 5,6- DiHETrE ↑↑ 8,9- DiHETrE ↑↑ 11,12- EpETrE ↑↑ 11,12- DiHETrE ↑↑ 14,15- DiHETrE ↑ 16-HETE ↑↑ 17-HETE ↑↑ 18-HETE ↑↑ 19-HETE ↑↑ 20-HETE ↑ 9,10- EpOME ↑ 9,10- DiHOME ↑ 12,13- DiHOME ↑↑ 19,20- DiHDPE ↑↑ 20-COOH- AA ↑↑ 20-HDoHE |
| Non- Enzymatic | - | ↓ 8-iso- PGF2α | - | - | ↓ 15-keto- 15-F2t-iso ↓ 9-epi- 15-F2t-iso ↓ 5-epi-5F2t-iso | ↑↑ 5-iso- PGF2α-VI | - | - | ↑ 8,12-iso- Isoprostane- F2α-VI ↑ 13,14- Dihydro-15-keto-PGF2α |
↑↑ > 8-fold changes; ↑↑ 4 to 8-fold changes; ↑ 2 to 4-fold changes; ↑ < 2-fold changes. Late responses (after 5 h) were reported with arrow in gray scale. The arrows represent the fold changes between pre- and post-exercise condition (relative to greater difference value obtained for each metabolite). The background colors represent the metabolic pathways: COX (red), LOX (blue), CYP (green), and non-enzymatic pathway (yellow). The metabolites screened are those that had a statistical difference between the pre- and post-exercise condition. 20-COOH-AA: 20-carboxy arachidonic acid; 20-COOH-LTB4: 20-carboxy-leukotriene B4; COX: cyclooxygenase; CYP: cytochrome P450; 15d-D12,14-PGJ3: 15-deoxy-D12,14-prostaglandin J3; dh-PGE2: dihomo-prostaglandin E2; 13,14-dihydro-15-keto-PGE2: 13,14-dihydro-15-keto prostaglandin E2; 11-dihydro-TXB2: 11-dihydro thromboxane B2; 2,3-dinoβ-11-PGF2α: 2,3-dinor-11β-prostaglandin F2α; DiHDoHE: dihydroxy-docosahexaenoic acid; DiHDPE: dihydroxy-docosapentaenoic acid; DiHETE: dihydroxy-eicosatetraenoic acid; DiHETrE: dihydroxy-eicosatrienoic acid; DiHOME: dihydroxy-octadecenoic acid; EpETrE: epoxy-eicosatrienoic acid; EpOME: epoxy-octadecenoic acid; HDoHE: hydroxy-docosahexaenoic acid; HEPE: hydroxy-eicosapentaenoic acid; HETE: hydroxy-eicosatetraenoic acid; HETrE: hydroxy-eicosatrienoic acid; HHTrE: hydroxyheptadecatrienoic acid; HODE: hydroxy-octadecadienoic acid; HOTrE: hydroxy-octadecatrienoic acid; Iso: isoprostane; 15-keto-PGE2: 15-keto prostaglandin E2; 6-keto-PGF1α: 6-keto prostaglandin F1α; 15-keto-PGF2α: 15-keto prostaglandin F2α; LOX: lipoxygenase; LT: leukotriene; LX: lipoxin; oxo-ETE: oxo-eicosatetraenoic acid; 12-oxo-LTB4: 12-oxo leukotriene B4; oxo-ODE: oxo-octadecadienoic acid; PD1: protectin D1; PG: prostaglandin; PGDM: prostaglandin D metabolite; 11-β-PGF2α: 11β-prostaglandin F2α; PGFM: prostaglandin F metabolite; 15R-LXA4: 15R-lipoxin A4; Rv: resolvin; Tetranor-PGDM: tetranor-prostaglandin D metabolite; tetranor-PGEM: tetranor-prostaglandin E metabolite; TX: thromboxane.
Table 4.
PICOS search strategy.
| PICOS | |
|---|---|
| Population | Healthy adult subjects |
| Intervention | Physical Exercise influence |
| Comparator | Non-exercise/intervention condition |
| Outcome | Exercise-related oxylipins |
| Study Design | Analytical studies |
Table 5.
Score setting adjusted for lipidomics studies quality assessment.
| Score Setting | |||
|---|---|---|---|
| Section | Maximum Score | Aspects | Score Attribution |
| Research Design | 2 | Number of Participants | Parallel Studies 0—n < 20 2—n > 20 Crossover Studies 0—n < 13 2—n > 13 |
| 2 | Study Characteristics | —Randomized control group —Proper matrix —> 2 timepoints data collection —Duration ≥ 3wk (chronic studies only) 0—None of the previous items 1—At least 2 of the first 3 criteria listed 2—All 3 of the first 3 criteria listed | |
| Methodology | 3 | Analysis Methods | 1—< 10 oxylipins measured using global metabolomics 2—40–10 oxylipins measured using LC-MS/MS targeted oxylipins panel 3—> 40 oxylipins measured using LC-MS/MS targeted oxylipins panel |
| 2 | Statistical Support | 0—simple univariate statistics 1—Univariate statistics + additional analyses to sort and group the data, and to control for confounding factors 2—univariate statistics + PCA, OPLS-DA, PLS-DA, or similar advanced bioinformatics procedures | |
| Novelty | 2 | 0–2—New information in the literature | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Signini, É.F.; Nieman, D.C.; Silva, C.D.; Sakaguchi, C.A.; Catai, A.M. Oxylipin Response to Acute and Chronic Exercise: A Systematic Review. Metabolites 2020, 10, 264. https://doi.org/10.3390/metabo10060264
AMA Style
Signini ÉF, Nieman DC, Silva CD, Sakaguchi CA, Catai AM. Oxylipin Response to Acute and Chronic Exercise: A Systematic Review. Metabolites. 2020; 10(6):264. https://doi.org/10.3390/metabo10060264
Chicago/Turabian StyleSignini, Étore F., David C. Nieman, Claudio D. Silva, Camila A. Sakaguchi, and Aparecida M. Catai. 2020. "Oxylipin Response to Acute and Chronic Exercise: A Systematic Review" Metabolites 10, no. 6: 264. https://doi.org/10.3390/metabo10060264
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.