
The Role of Nitroreductases in Resistance to Nitroimidazoles



1
School of Natural Sciences, Bangor University, Bangor LL57 2UW, UK
2
Department of Chemistry, Xi’an Jiaotong-Liverpool University, 111 Ren’ai Road, Suzhou Industrial Park, Suzhou 215123, China
*
Author to whom correspondence should be addressed.
Biology 2021, 10(5), 388; https://doi.org/10.3390/biology10050388
Submission received: 5 January 2021
/
Revised: 4 February 2021
/
Accepted: 10 February 2021
/
Published: 1 May 2021
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Antimicrobial resistance continues to be a major global health threat. It is estimated by the WHO that 700,000 people die each year because of drug resistance, and this is predicted to rise to 10 million by 2050. As well as the increased cost, which is forecast to exceed $100 trillion, as more expensive drugs have to be deployed, illnesses often last longer and require hospital treatment. This, in turn, increases the strain on often-inadequate healthcare systems. As resistances continue to grow, finding alternatives is crucial. This review showed that nitroreductases play a role in drug activation but are also associated with resistance mechanisms. These mechanisms require further investigation to fully understand them before they can be utilised against multidrug-resistant organisms. This will depend on committed collaborations between the private and public sector to translate academic research into the clinic.
Abstract
Antimicrobial resistance is a major challenge facing modern medicine, with an estimated 700,000 people dying annually and a global cost in excess of $100 trillion. This has led to an increased need to develop new, effective treatments. This review focuses on nitroimidazoles, which have seen a resurgence in interest due to their broad spectrum of activity against anaerobic Gram-negative and Gram-positive bacteria. The role of nitroreductases is to activate the antimicrobial by reducing the nitro group. A decrease in the activity of nitroreductases is associated with resistance. This review will discuss the resistance mechanisms of different disease organisms, including Mycobacterium tuberculosis, Helicobacter pylori and Staphylococcus aureus, and how these impact the effectiveness of specific nitroimidazoles. Perspectives in the field of nitroimidazole drug development are also summarised.
1. Introduction
Nitroreductases (NTR) are a family of proteins involved in the reduction of nitro-containing compounds [1]. The flexibility of these enzymes comprises their usefulness in a variety of biological and medical applications. These include environmental decontamination using bioremediation [2]; various cancer therapies such as gene and viral-directed prodrugs [3,4,5,6,7] and probes for detecting hypoxia in tumours [8,9,10,11], antiparasitics [12,13], herbicides [14] and for the detection of explosives [15]. For a more detailed review of the many uses of nitroreductases, see the papers by Kumari et al. [11], Zhang et al. [4] and Nepali et al. [16].
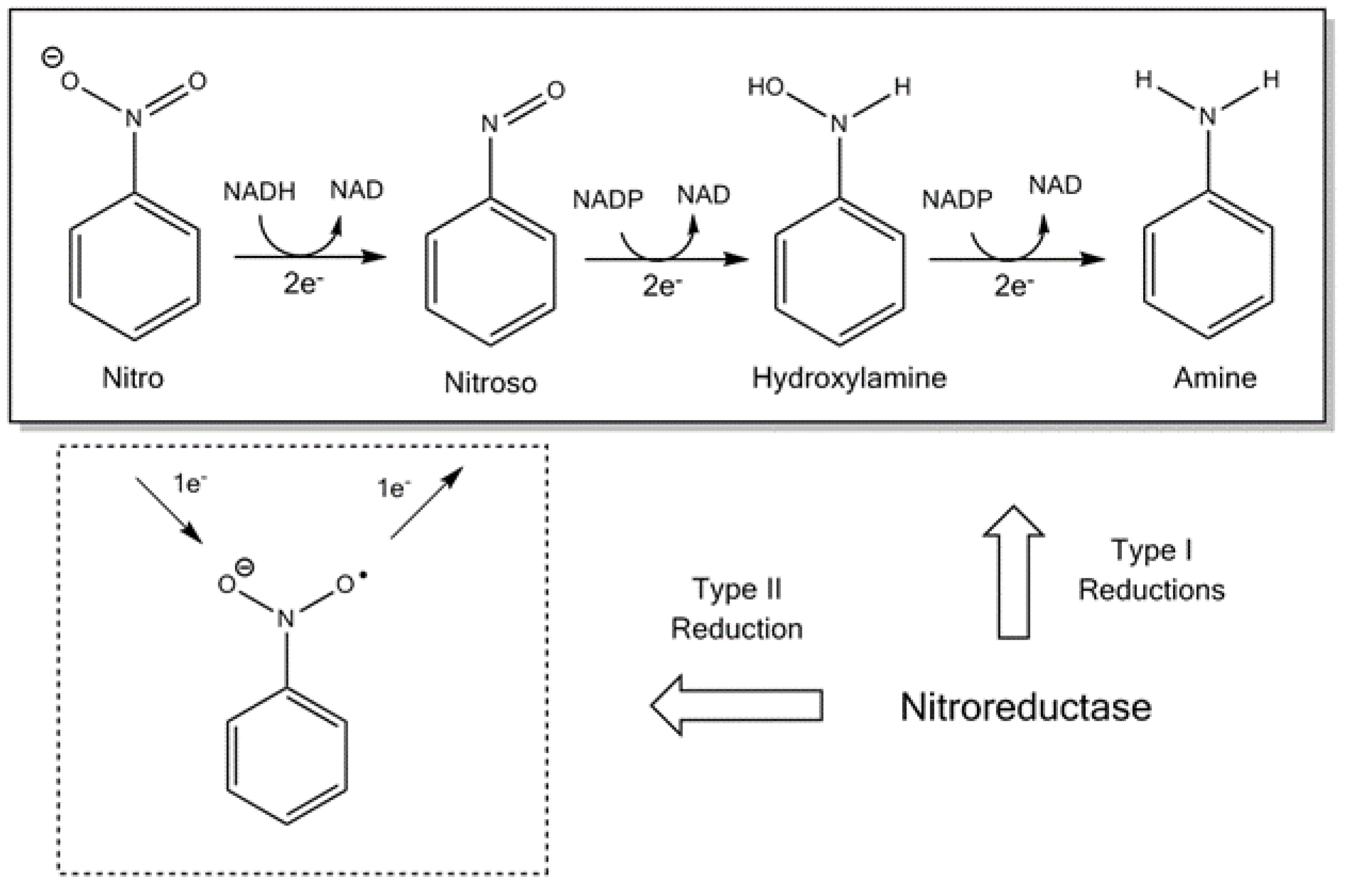
Nitroreductases can be divided into two groups: flavin reductases [17] and those from enteric bacteria [18]. Typical NTRs share similar biochemical properties; they usually occur as a homodimer, contain flavin mononucleotide (FMN) as a cofactor and catalyse using the ping-pong bi-bi kinetic mechanism [1,19]. The bacterial NTRs can be further split into Type I, oxygen-insensitive and type II, which are oxygen-sensitive, as illustrated in Figure 1 [20]. Bacteria can contain both type I and II, although the most studied are those enzymes that belong to type I [19]. The oxygen-insensitive NTRs can be further divided into major and minor protein groups. The minor group can utilise nicotinamide adenine dinucleotide (NAPH) or nicotinamide adenine dinucleotide phosphate (NADPH), while the major group can utilise NADPH only as the electron donor [21]. The most studied examples of the major and minor groups are the Escherichia coli NfsA and NfsB enzymes [21].
The physiological functions of NTRs are not completely understood, but several have been proposed for oxygen-insensitive bacterial NTRs [19]. It has been assumed that they have a role to play in detoxification, as they reduce a broad range of compounds [19]. Some are involved in specific degradation pathways, such as nitrobenzene reductase and nitrophenol reductase [22,23]. NTRs may also be involved in the response to oxidative stress, as the enzyme NfsA is regulated by the SoxRS system, which is involved in the prevention of oxidative damage [24,25]. The range and adaptability of NTRs have enabled some of these enzymes to specialise in different metabolic functions without necessarily losing the other reductase activities [19]. Therefore, while some NTRs are associated with specific metabolic pathways or the reduction of different nitroaromatic compounds, others may be active in processes such as oxidative stress response or bioluminescence [26]. Due to the ability of NTRs to affect the toxic, mutagenic and carcinogenic characteristics of many nitroaromatics, nitrofuran derivatives have been used to develop a group of antimicrobials commonly knowns as nitroimidazoles [27].
Nitroimidazoles have seen a resurgence in interest due to their broad spectrum of activity against anaerobic Gram-negative and Gram-positive bacteria. They are particularly being seen as important in the battle against antibiotic resistance. Resistance to antibiotics is a major challenge facing modern medicine, and its importance was highlighted in 2016 when it was a topic for discussion at the United Nations General Assembly [28]. It is estimated that 700,000 people die each year because of infections that are resistant to current antibiotics and that this figure is predicted to rise to 10 million annually by 2050, with the global cost forecast to exceed $100 trillion over the next few decades [29]. The overuse of antibiotics in both clinical and agricultural settings has accelerated the process of resistance [29,30]. Perhaps the most widely known antibiotic is penicillin, which was discovered by Fleming in 1928, although it was not produced on any great scale until 1940 [30]. Most current antimicrobials were discovered between the 1940s and 1970s, including nitroimidazoles in the early 1950s, when azomycin was isolated from a crude extract of Streptomyces bacteria [31]. This review will look at the role played by NTRs in the resistance mechanisms of nitroimidazoles.
2. Nitroimidazoles
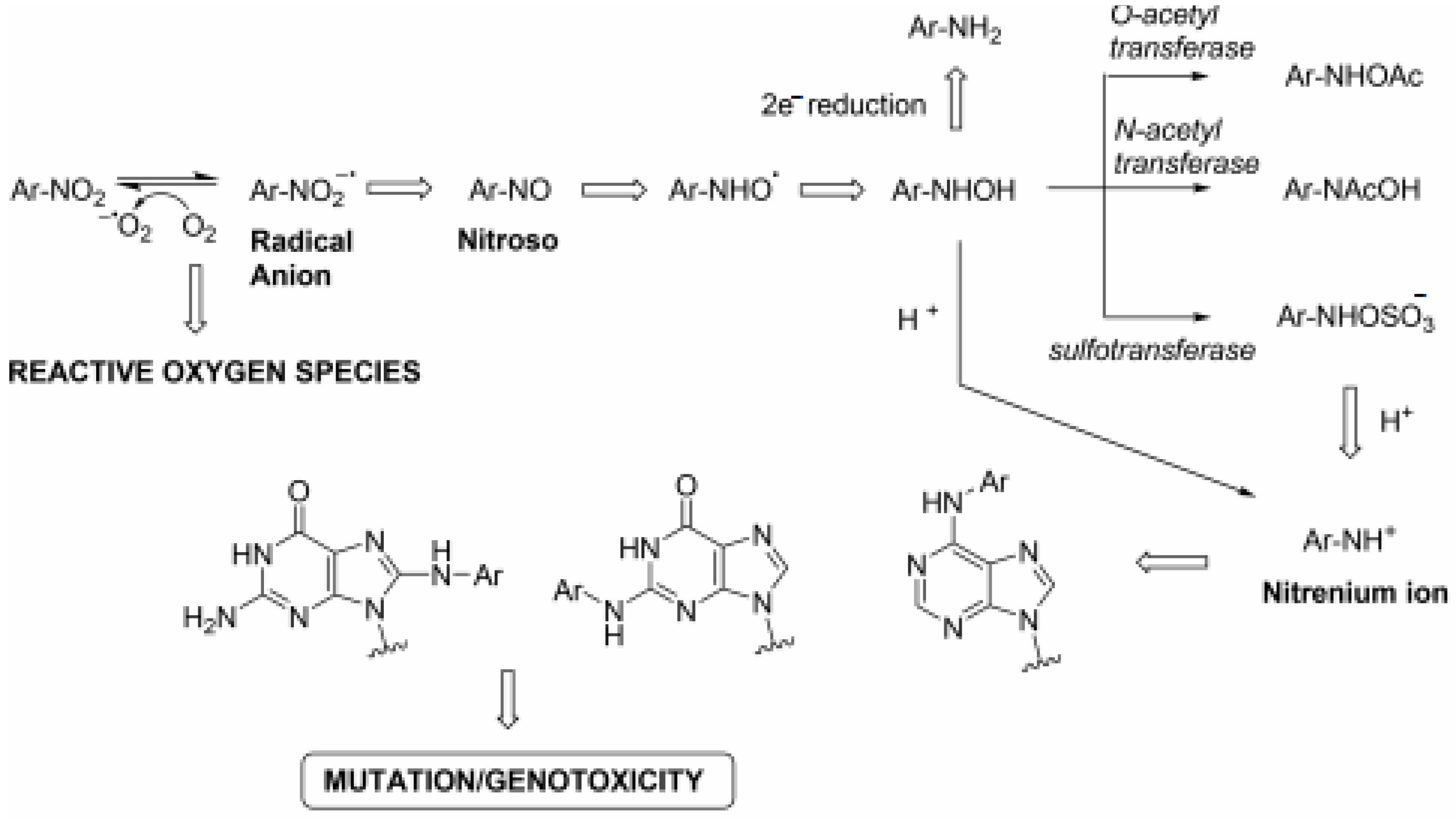
Nitroimidazoles are a class of antimicrobial drugs that have a broad spectrum of activity against anaerobic Gram-positive and Gram-negative bacteria, as well as parasites and mycobacteria, since their discovery drugs such as metronidazole, pretomanid and delamanid have gone on to form a large part of the treatment for Helicobacter pylori and Mycobacterium tuberculosis, respectively. The mode of action of nitroimidazoles can help to explain why they have such a broad spectrum of activity. They are prodrugs that require the reduction of the nitro group before they display any antimicrobial effects. This is normally achieved by NTRs using flavin mononucleotide (FMN) or flavin adenine dinucleotide (FAD) as prosthetic groups and either NADH or NADPH as reducing agents [27]. The mechanism is understood to have the following steps: (i) molecules enter the cells through passive diffusion, (ii) the nitro group is reduced to reactive radical species and (iii) the radicals react with the DNA or protein within the cell [27]. The reduction of products within a cell depends on the redox potential of the compound and the number of electrons being transferred [32]. The system operates more efficiently under anaerobic conditions, meaning the bactericidal effects are increased compared to when oxygen is present [33]. The complete pathway can be seen in Figure 2. The diverse mode of action, which is often inadequately defined, can lead to problems when trying to maximise their bactericidal potential. It is also a factor that must be taken into consideration when looking at the important issue of resistance.
Resistance to nitroimidazoles usually occurs because there is a decrease in the activity of the enzymes responsible for the reduction of the nitro group [34]. These resistance mechanisms differ depending on the target organism and will be described in more depth in the sections below. To develop new drugs that will aid the management of infectious diseases, these resistance mechanisms must be better understood.
3. Metronidazole
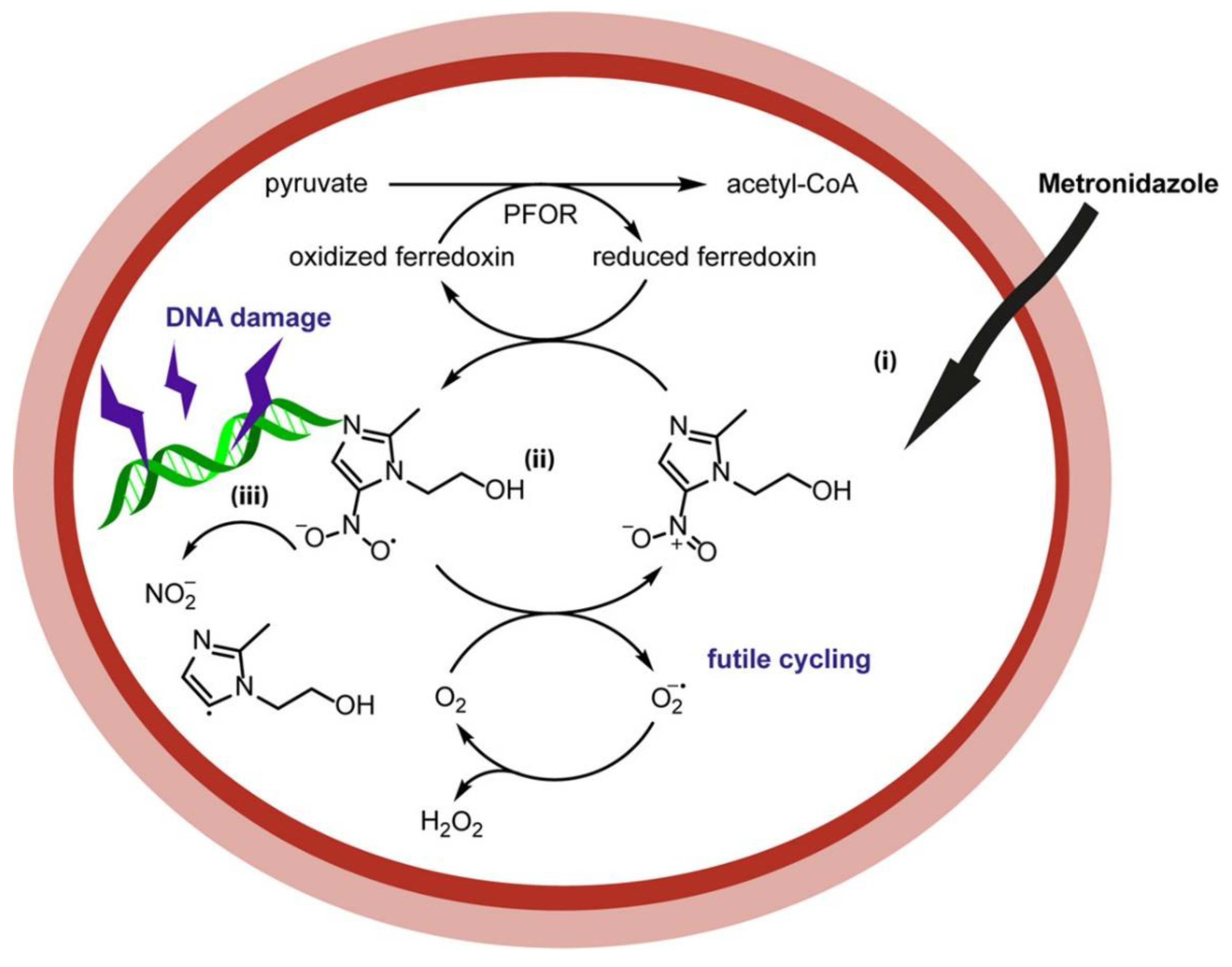
Metronidazole (MTZ) is a nitroimidazole prodrug derived from azomycin, which has been used as an antimicrobial since the early 1960s [35]. The drug enters the cells via passive diffusion but is inactive until the nitro group is reduced. This can occur via two routes (Figure 3): reductive activation, which results in toxicity, or reductive inactivation, where the nitro group is reduced to a nontoxic amino derivative [19,36]. MTZ is one of the main drugs used to treat Helicobacter pylori, which is a Gram-negative microaerophile bacterium that can be found in the stomach of almost 50% of the world’s population. The symptoms include chronic gastritis and peptic ulcers. It is also classed as a type I carcinogen by WHO, as it is a risk factor in gastric adenocarcinoma and mucosa-associated lymphoid tissue lymphoma (MALT) [37,38,39].
Resistance to MTZ in cases of H. pylori is more common than with other antimicrobials and can be as high as 80%, depending on the patient group and geographic region [40]. As the mechanism of action requires enzymatic reduction, anything which affects this can cause resistance. Mutations in the genes that encode certain electron transport proteins are one such example; inactivation of the rdxA (encodes oxygen-insensitive NADPH nitroreductase) and frxA (encodes NADPH flavin oxidoreductase) genes is linked to MTZ resistance in H. pylori [41,42]. Chua et al. [43] demonstrated a strong correlation between mutations that inactivate RdxA and its resistance. They also reported that a significant number of MTZ-resistant strains of H. pylori had a mutation of the Arg-16 residue of RdxA. This amino acid residue is responsible for binding between RdxA and the FMN phosphoryl group, and therefore, any mutation may adversely affect the reduction of MTZ to its cytotoxic form [43]. However, this cannot be responsible on its own for conferring MTZ resistance, as it has also been found in H. pylori strains that were susceptible to MTZ [43]. Chua et al. [43] proposed that high levels of frxA would counteract the effect of Arg-16 and make H. pylori susceptible to MTZ, despite any mutation of rdxA. If frxA is inactivated, it does not appear to play a major role in MTZ-resistance but may work together with other mutations to increase the resistance [43].
As resistance to MTZ can occur without either rdxA or frxA inactivation, other mechanisms must be considered [44,45]. One such mechanism suggested by Lee et al. [46] is a gene, hefA, associated with the efflux pump HefABC. The expression of this gene is increased when exposed to MTZ. In MTZ-resistant strains of H. pylori with intact frxA and rdxA, the levels of hefA were higher than in susceptible strains. If hefA was knocked out, the strains became sensitive to MTZ, and this was reversed by the addition of hefA, showing that hefA was directly involved in MTZ-resistance [46]. While this could help to explain cases of MTZ resistance in the presence of functioning frxA and rdxA genes, the overexpression of hefA alone does not result in resistance [46]. Clearly, the resistance of H. pylori to MTZ is a complex mechanism that requires further study to fully understand.
Nitroimidazole resistance at low levels is often associated with the nim genes [47]. The Nim genes were first described in 1994 [48], and their nitroreductase activity was observed two years later when a nimA-positive strain of Bacteroides fragilis reduced a nitroimidazole drug to its noncytotoxic amine derivative [49]. Currently, eleven nim genes have been identified: nimA–nimK [50].
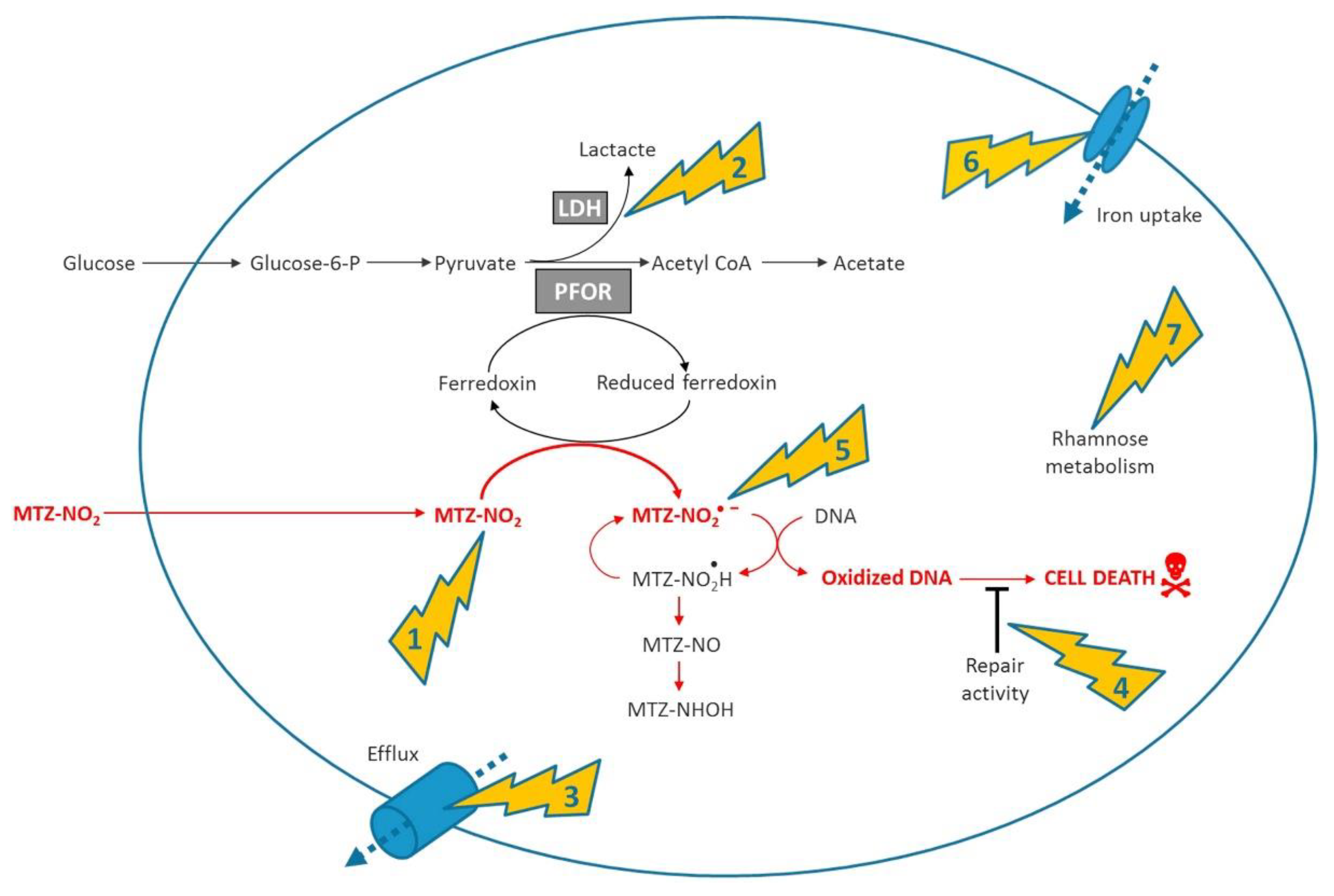
The exact mechanism of nim resistance is not fully understood, and there is conflicting evidence for the role of nim genes in metronidazole (MTZ) resistance [50,51]. Figure 4 below shows the involvement of nim genes in the mechanism and resistance of metronidazole. The overexpression of nimA, nimE and nimJ induces an increase in MTZ resistance in E. coli and B. fragilis, respectively [52,53].
However, several Bacteroides spp. and genera within the Clostridia class are resistant to MTZ without containing the nim gene, while other nim-positive species are MTZ-susceptible [54,55,56,57,58]. Thus, the presence of a nim gene does not automatically equal resistance. Gal et al. [54] found that only about half of the 50 Bacteroides strains positive for the nim gene presented minimum inhibitory concentrations (MICs) ranging from 16 to >32 µg/mL—therefore, above the resistance breakpoint. Sethi et al. [59] looked at the MTZ resistance rates in India and the relationship of isolates positive for a nim gene with resistance. They found a relatively high rate of resistance (31% compared to <1% in Europe), with a 53% rate of positivity for the nim gene. Out of 20 samples, 12 that were positive for the nim gene also showed resistance to MTZ, a significant correlation [59]. Leitsch et al. [60] ascertained the levels of expression of the nim genes using 2D gel electrophoresis (2DE). The study looked at whether adaptation to an increased concentration of MTZ led to increased levels of the nim protein. As nim proteins are thought to act as nitroreductases that reduce MTZ to its noncytotoxic form, their profusion should correlate with high levels of MTZ resistance. To test this theory, a high-level MTZ resistance was induced in three strains of B. fragilis, but when the nim levels were measured, there was found to be no upregulation [60]. Such studies reinforce the complexity of MTZ resistance mechanisms and emphasise that nim genes alone are not sufficient to give high-level MTZ resistance [53,54,56]. Nim genes have been found in multidrug resistant strains of B. fragilis, together with other resistance genes [61]. A cluster of multidrug-resistant (MDR) B. fragilis isolates containing nimB, cfiA, ermF and tetQ genes was detected using whole-genome sequencing (WGS) [61]. Nim genes have also been found in an MDR strain of B. thetaiotaomicron together with two β-lactamase genes, two tetX genes—tetQ and ermF—two cat genes and several genes encoding efflux pumps [62]. Despite this current inability to definitively demonstrate the cause and effect of nim genes and MTZ resistance, it is still worthy of further investigation as important risk factors, such as the location of the nim genes on mobile genetic materials plus the high usage of MTZ as a frontline drug in multiple infectious diseases could lead to an increase in resistance [50]. This is underlined by the presence of nim genes in several MDR anaerobes [63].
4. Delamanid and Pretomanid
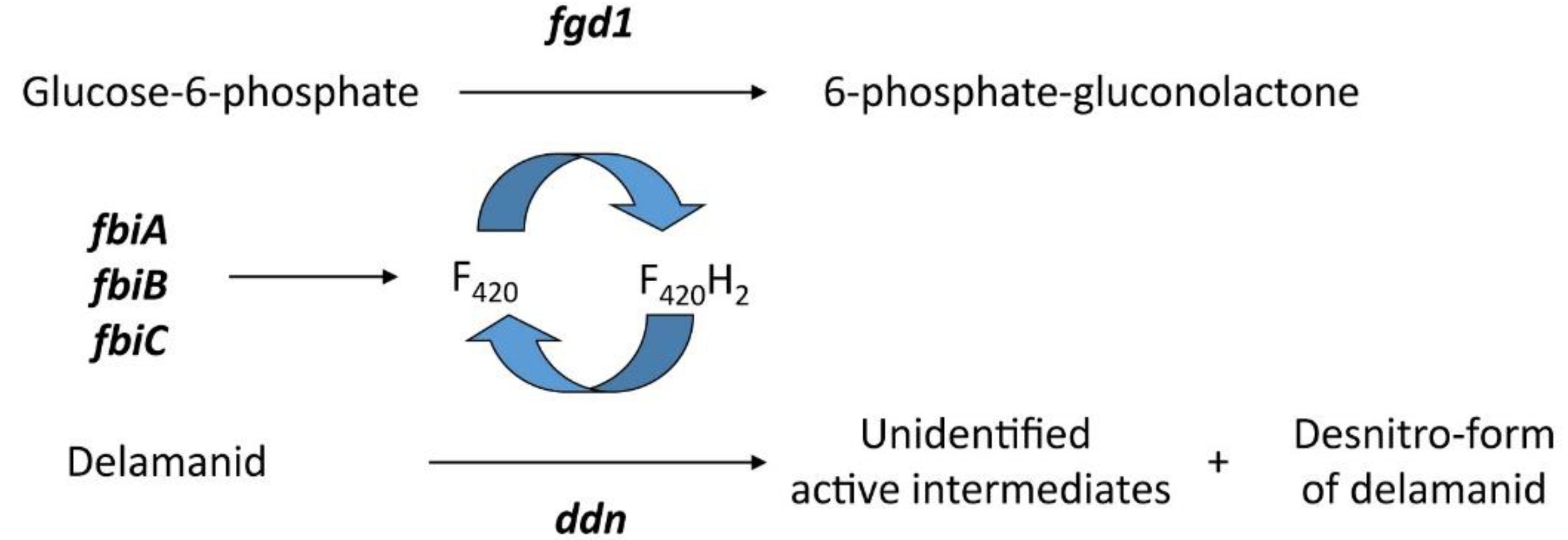
Delamanid and pretomanid are both bicyclic 4-nitroimidazoles, and phase II clinical trials have shown them to be effective against both replicating and hypoxic nonreplicating mycobacteria [64]. Tuberculosis (TB) is the leading cause of death from infectious disease. In 2018, 10 million people became ill with TB, with 1.2 million deaths [65]. Under aerobic conditions, the mechanism of action is to inhibit the formation of mycolic acids [66], while under anaerobic conditions, the mechanism induces respiratory poisoning [67]. Pretomanid, in particular, donates nitric oxide (NO), which can create toxic conditions within the bacilli in nonreplicating MTB [67]. Both delamanid and pretomanid are prodrugs that require bioactivation of the nitro group to become effective. They are activated in an F420H2-dependant reaction in MTB by deazaflavin-dependant nitroreductase (Ddn) [67,68]. Therefore, mutations that affect the activity of Ddn or the biosynthesis or reduction of F420 may result in resistance [64].
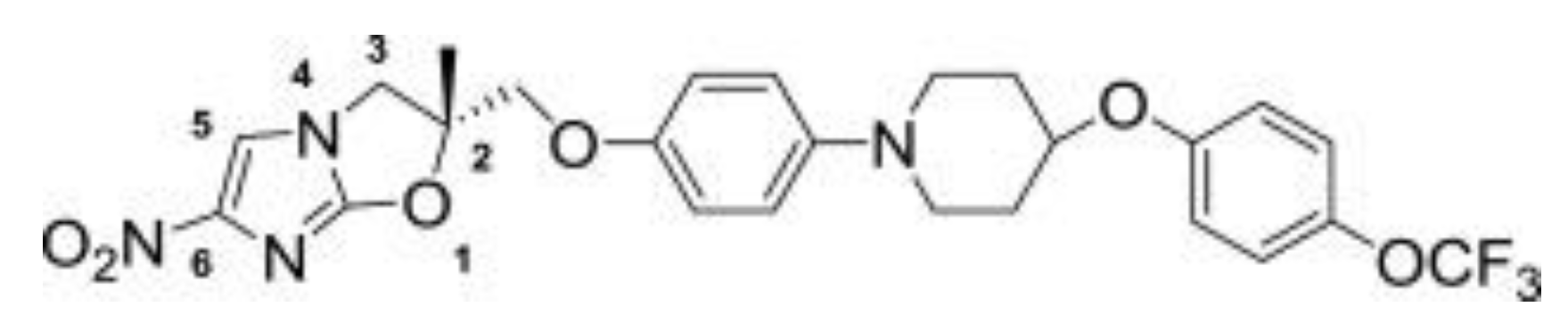
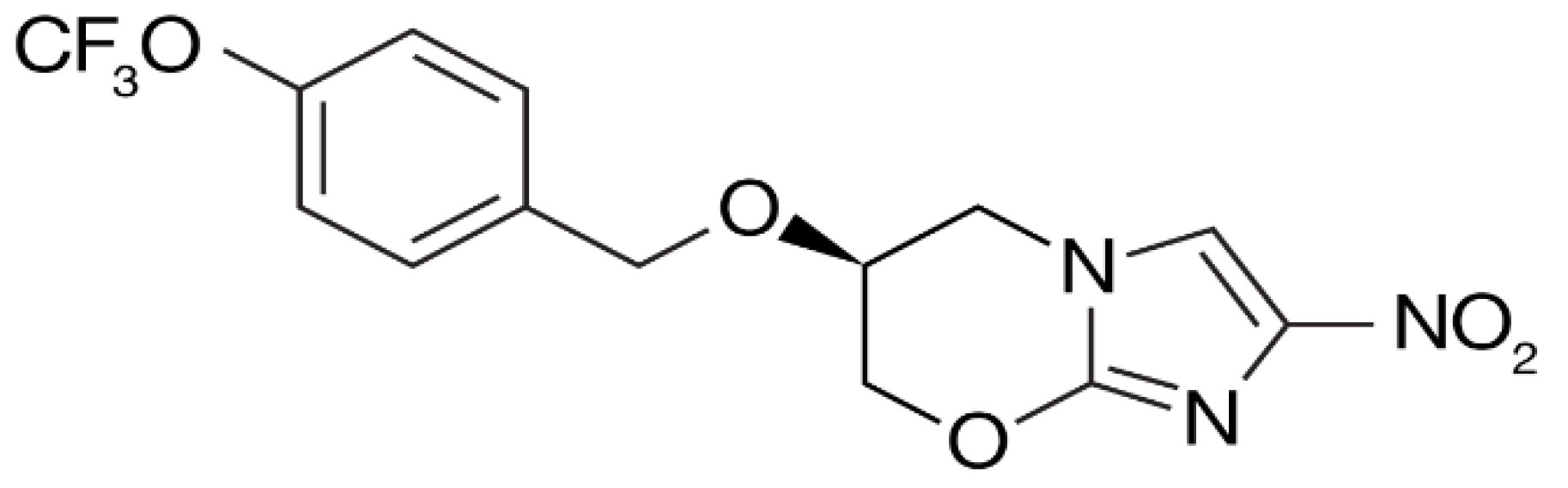
Delamanid (Figure 5) was discovered by Otsuka Pharmaceuticals [69] and was granted conditional approval in 2014 by the European Medicines Agency (EMA) for the treatment of pulmonary MDR-TB in adults in combination with other frontline drugs [70].
The World Health Organisation (WHO) issued a policy guidance, which stated that delamanid could be added to the WHO-recommended regimen [71]. It has a potent minimum inhibitory concentration (MIC) range of 6–240 ng/mL, which is the lowest among the current TB drugs [69,72]; plus, it showed no signs of toxicity in the Ames test [73]. In a phase IIa trial, delamanid showed good activity in drug-susceptible TB patients, but due to poor adsorption at high doses, twice-daily dosing was required (2 × 50-mg tablets per dose) [74,75]. The compound works by inhibiting the synthesis of cell wall components and is active against replicating and nonreplicating persister bacteria, both extra and intracellular bacilli [69]. It does not show a cross-resistance with other TB drugs and can therefore be used in combinations [76]. A phase II trial was completed in 2012 in combination with an optimised background regime (OBR) in 17 centres across nine countries [77]. Phase III trials have since been completed, and further investigations into its use in children are underway [77].
Dormant bacteria are often resistant to treatment, and therefore, new drugs with the ability to kill dormant bacilli are more likely to be effective against MDR-TB. The Wayne model is being used to test the efficacy of TB drugs under low oxygen concentrations [78,79]. Various studies have used a modified version of this model to investigate the bactericidal effects of delamanid in aerobic versus anaerobic settings, with Upton et al. [80,81] showing that it killed 99% of MTB bacilli at 4.4 µg/mL. Delamanid was also as effective as the frontline drug rifampicin against intracellular mycobacteria [82].
A resistance to delamanid has been reported in clinical isolates of TB. Bacilli resistant to delamanid have mutations in one of the five genes (Figure 6) associated with the F420-dependant nitroreduction pathway: fgd1, ddn, fbiA, fbiB and fbiC [83]. FbiA, fbiB and fbiC are coded for proteins FbiA, FbiB and FbiC, which are essential for the biosynthesis of F420, with each gene affecting a different stage [84,85]; fgd1 (gloucose-6-phospahte dehydrogenase) play a role in the F420 redox recycling mechanism and ddn is the reductase required for activation [86].
Fujiwara et al. [86] showed that, from 30 randomly selected resistant colonies, the frequency of mutations across the five genes varied considerably: ddn (20%), fgd1 (30%), fbiA (16.7%), fbiB (6.7%) and fbiC (26.7%). The fact that resistant bacilli lack the ability to activate delamanid is shown by the following; when drug-susceptible MTB bacilli were incubated with delamanid, the concentration decreased as it was converted to the des-nitroimidazole form, but when resistant bacilli were used, no change was seen [82,86].
Pretomanid (Figure 7) was identified by PathoGenesis and was approved by the FDA in 2019 for the treatment of multidrug-resistant TB in combination with bedaquilne and linezolid [87]. The MIC values range from 150 to 250 ng/mL, and no cross-resistance with other TB drugs has yet been seen, making it a good candidate for treating MDR-TB [88]. Studies have shown that pretomanid, in combination with moxifloxacin and pyrazinamide, had better bactericidal activity than the current standard regimen [89,90]. The primary mechanism of action is the inhibition of synthesis of cell wall lipids and proteins [91].
Resistant mutants have shown that both Ddn and FGD1 are essential for the drug to become activated; however, only Ddn is involved in reducing the nitro group, producing three primary metabolites and reactive nitrogen species [67]. A study by Haver et al. [88] looked at 183 spontaneous pretomanid-resistant mutations of which 29% of the lesions were seen in ddn, 26% in fbiC, 19% in fbiA, 7% in fgd1 and 2% in fbiB; the remaining 17% did not show mutations in any of the five genes. Eighty-three percent had single mutations in one of the genes. As 17% had no mutations within the five genes, it can be hypothesised that other targets are involved in either the mechanism of action or activation pathway of pretomanid [88].
As stated previously, mutations that knock out the activity of Ddn completely may result in resistance in both delamanid and pretomanid. However, this could affect the fitness of MTB, as the cofactor F420 has been shown to be essential for the survival of MTB, used by at least 28 enzymes [92] that play important roles in the hypoxic recovery and evasion of the hosts immune system [93,94]. Therefore, in order for nitroimidazole resistance to spread, the native activity of Ddn must either be retained or compensated for. The activation of delamanid and pretomanid by Ddn is a promiscuous activity that is more prone to mutation than native functions [95], meaning that the prodrug activation could be lost without loss of the native function. Lee et al. [96] conducted studies to better understand the fitness costs of the loss of Ddn activity. They were able to demonstrate, through studying 75 mutants, that a number were able to prevent the activation of pretomanid without the loss of all native functions. Therefore, these mutants would be fit enough to spread to new patients [96]. These mutations are likely to be one of the main routes through which resistance to these two new TB drugs will spread. Of the 75 mutants studied, 25 did not reduce pretomanid, but only 10 lost the ability to reduce delamanid, despite their similarities, suggesting that they interact with Ddn in different ways [96]. Clearly, the results from this study should be considered when looking at the continued development and clinical use of nitroimidazoles. Both delamanid and pretomanid are undergoing phase II/III trials, in combination with other drugs, against drug-susceptible TB and MDR-TB. Mutations of Ddn should be closely monitored to ensure that the best drug combination is used, and that will ultimately reduce the spread of resistance. The indication that delamanid binds to Ddn differently than pretomanid means that it could be used to treat some TB strains that are resistant to pretomanid [96]. A combination of the two could also be used to reduce the spread of resistance. Further testing of a range of nitroimidazoles against a variety of Ddn variants could help to identify drugs which are less likely to develop resistance.
To fully understand the resistance mechanism, Fujiwara et al. [86] suggested that a rapid drug susceptibility test should be developed. Presently, only phenotypic tests are available for delamanid based on cultures, with results taking several weeks [97]. However, the development of a molecular based test has been problematic due to the distribution of mutations across five genes [86].
One reason why drugs containing a nitro group have not been more widely used is because of the associations with toxicity, including bone marrow suppression, hepatotoxicity and carcinogenicity [16]. Both types of nitroreduction (I and II) produce toxicity; in type I, this is as a result of the generation of hydroxylamine, and during type II, the generation of reactive oxygen species (ROS) via redox cycling causes pharmacological activity [98]. Therefore, if they are going to play any part in the fight against drug resistance, any new developments must aim to reduce these issues. Drugs such as delamanid and pretomanid have been shown to not be mutagenic or genotoxic, but their use is hampered by issues with solubility. This is an area that could be further investigated by formulation chemists [16].
5. Chloramphenicol
The ever-increasing resistance to modern antibiotics has led to the re-evaluation of drugs that had either limited use or were discarded due to toxicity or efficacy issues. One such antimicrobial that is now being reassessed is chloramphenicol [99,100]. Chloramphenicol can cross the blood–brain barrier and other sites that can be hard to target, which makes it a powerful drug in the treatment of bacterial meningitis [99]. However, chloramphenicol has also been linked with haematological toxicity [101], and its use is carefully controlled in developed countries [101]. As a possible consequence of this, it can still be effective against multidrug-resistant organisms, including MRSA [102,103].
The main mechanisms of chloramphenicol resistance occur through functions such as enzymatic inactivation via chloramphenicol acetyltransferance, efflux pump removal and ribosome protection [104]. The bacterial modification of chloramphenicol was first reported only two years after it was discovered. Several species of bacteria were reported as being able to reduce the nitro group, with the resulting compound having no antimicrobial effect [104]. Crofts et al. [104] set out to identify the bacterial genes that could be involved in this reduction mechanism. They chose to focus on type I oxygen-insensitive nitroreductases—in particular, NfsB from Haemophilus influenzae. They determined the ability of H. influenzae NfsB to reduce the nitro group of chloramphenicol completely to an amine. The reduction is enough to confer resistance to chloramphenicol, but Crofts et al. observed that the H. influenzae NfsB enzyme also uses metronidazole (MTZ) as a substrate but does not completely reduce the nitro group. They hypothesised that, while NfsB protects the cell against chloramphenicol, it activates MTZ to its cytotoxic state, therefore offsetting any survival benefits. They concluded that the two drugs could be used together to combat bacteria resistant to chloramphenicol on its own [104].
One of the reasons that chloramphenicol has not been more widely used is the concern that its use could result in the development of aplastic anaemia [105]. The mechanism through which this happens is not fully understood, although it is thought that nitro reduction is involved. The identification of a bacterial enzyme that has been shown to reduce chloramphenicol means that this theory can now be tested and hopefully identify analogues of chloramphenicol that can be used in the clinic [104].
6. MT02
The antimicrobial drug MT02 has been identified as a new candidate that is active against Gram-positive bacteria such as Staphylococcus aureus [106]. S. aureus is a common pathogen associated with a wide range of infections, both superficial and invasive [107]. Together with S. epidermidis, it accounts for more than 20% of all infections associated with hospitalisation, affecting over 250,000 patients in the USA and Europe annually [107]. The first antibiotic used to treat S. aureus was penicillin; prior to this, the infection was normally fatal [108]. Resistant strains soon appeared, which, in turn, stimulated the search for new antibiotics in the 1950s, such as streptomycin and tetracycline, but resistance soon developed once they were being routinely used in the clinic [109,110]. The development of methicillin was a result of the search for an antibiotic that would be active against penicillin-resistant S. aureus. Methicillin-resistant S. aureus strains (MRSA) emerged in the 1960s, rendering the drug ineffective [111]. MT02 is a nitro-active compound, and the work carried out by Menzel et al. [106] in 2011 showed that it is a DNA-binding compound leading to the inhibition of DNA replication. It shows high levels of antimicrobial activity against S. aureus and other Gram-positive bacteria [106]. El-Hossary et al. [112] carried out further work to identify the potential of S. aureus to develop resistance to MT02 [112]. In order to select MT02-resistant clones, S. aureus strain MA12 was cultivated with increasing concentrations of MT02. Once the concentration reached 10 µM, a red colour began to appear, which then intensified with increasing concentrations. Work to identify the structure of this red compound showed the stepwise reduction of the aromatic nitro groups of MT02 to amino groups. The conclusion was that this reduction was induced by an enzyme found in the resistant S. aureus strain [112]. Four previously identified enzymes with nitroreductase activity (SAUSA300_0788, SAUSA300_0381, SAUSA300_1986 and SAUSA300_2462) were found not to be responsible for this reduction of MT02 [112]. The group carried out total RNA sequencing (RNA-seq), which identified the overexpression of an assumed nitroreductase SAUSA300_0859. This was confirmed by RT-PCR, which showed a 160-fold overexpression of SAUSA300_0859 in the MT02-resistant strain. The importance of SAUSA300_0859 in the development of resistance to MT02 was confirmed when sensitivity was restored by a transposon insertion mutant. A biochemical analysis showed that the SAUSA300_0859-encoded protein produced an intense red colour when incubated with MT02, demonstrating that it is the functional enzyme giving resistance in S. aureus [112]. While the molecular mechanism that leads to the overexpression of SAUSA300_0859 is not known, this is thought to be the first example of a nitroreductase-based antibiotic resistance mechanism in S. aureus.
7. Future Perspectives
As resistances to antimicrobials continue to grow, finding alternatives is crucial. As this review demonstrated, nitroimidazoles have a broad range of activity against many different organisms, and it would therefore be prudent to re-examine their potential. Several strategies that merit further investigation are briefly discussed below.
As well as looking for ways to combat the issue of toxicity, the mechanism of nitroimidazole resistance must be fully understood if effective drugs are to be developed. What is clear from the discussions above is that the role played by nitroreductases needs further investigation. This is of particular importance, as many bacteria have developed resistance to multiple drugs [113]. Strategies that aim to directly combat different resistance mechanisms are more likely to be effective in finding ways to treat drug-resistant organisms. The repurposing of existing antimicrobials and their use in novel combinations is one such strategy. Drug repurposing is defined as “an approved drug in one disease area is found to be active in another disease whereas drug repositioning is the uses of a drug active in one disease as a template for the synthesis of derivatives active in another disease” [114]. Drug rescue is defined as “developing new uses of a drug that failed to progress through clinical studies or which was removed from the market” [114]. All these strategies have been applied to nitroimidazoles.
Collaboration is important in drug development and repurposing. Open databases provide cost-effective access to the sharing of resources and data. Examples include PubChem and Chemspider, which contain chemical structural information, while DrugBank and SuperTarget provide profiles of drugs and target diseases [115]. As NTRs have such a broad spectrum of activity, the use of these databases presents a useful tool. For example, entering “nitroimidazole” as a search in PubChem (December 8th, 2020) identified 3479 molecules in the compound database and 1754 records in the Bioassay database. Research groups can use these databases to profile nitroimidazoles against a much greater number of microorganisms than would have previously been possible. This could potentially identify those with untapped potential. Along similar lines, the open innovation concepts, such as the Community for Open Antimicrobial Drug Discovery (CO-ADD) funded by the Wellcome Trust and The University of Queensland, encourage the open sharing of data and ideas. These initiatives aim to remove any barriers that could limit an individual group’s ability to develop suitable compounds by making resources widely available. This is particularly relevant to NTRs, as their complex modes of action require extensive investigation to fully understand [27].
As previously discussed, NTRs can be used as prodrug activators. Celik et al., [116] investigated the potential for NTRs to be used in antibiotic activation. They aimed to identify possibilities for redesigning existing drugs as precursor prodrugs that could then be activated by bacterial NTRs. They redesigned sulfamethoxazole as a prodrug, and when this was activated by a type I NTR, the results showed an improved antimicrobial action, thus demonstrating that the use of existing approved antibiotics in the form of pre-antibiotics could be used against disease-causing bacteria [116]. The use of antibiotic adjuvants is another strategy that has been shown to work [117]. These are small molecules that, although they have no antimicrobial actions, can increase the efficiency of antibiotics [114].
The development of more robust vaccines for TB would see the need for aggressive treatments with antibiotics reduced. One such area of research is the use of latency-associated antigens, particularly those encoded by the dormancy survival regulator (DosR) regulon [118]. The current BCG vaccine offers poor protection against adult TB [119], as it does not protect against the bacteria when they are in the dormant phase. As most TB patients have a latent infection before any clinical signs are present, latency-associated antigens that are expressed during the dormant phase are of interest as potential vaccine candidates. Kwon et al. [120] selected Rv3131, which is a hypothetical NTR with two DosR-binding sites, as a vaccine candidate. They aimed to investigate whether Rv3131 had any potential as a vaccine against TB. They concluded that it showed promise, as it met the following criteria: it was recognised by the immune system in in vivo experiments, could induce Ag-specific Th-1 T cells and showed activity against highly virulent TB strains [120].
8. Conclusions
This review has shown that nitroreductases play an important role in drug activation but are also associated with resistance mechanisms. This review highlighted the urgent need for further investigations to fully understand these mechanisms before they can be utilised against multidrug-resistant organisms for the development of new drugs. This will depend on the committed collaborations between the private and public sectors to translate academic research into the clinic.
Author Contributions
All authors contributed equally to conception C.T. drafted the review and C.D.G. provided the funding a proof read. Both authors approved the version to be published, and agree to be accountable for all aspects of the work in ensuring that questions related to the accuracy or integrity of any part of the work are appropriately investigated and resolved. All authors have read and agreed to the published version of the manuscript.
Funding
The authors would like to acknowledge the Celtic Advanced Life Sciences Network (CALIN) which is supported by the European Regional Development Fund through the Ireland Wales Cooperation programme.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gwenin, C.D.; Kalaji, M.; Williams, P.A.; Kay, C.M. A kinetic analysis of three modified novel nitroreductases. Biodegradation 2010, 22, 463–474. [Google Scholar] [CrossRef]
- Van Dillewijn, P.; Couselo, J.L.; Corredoira, E.; Delgado, A.; Wittich, R.-M.; Ballester, A.; Ramos, J.L. Bioremediation of 2,4,6-Trinitrotoluene by Bacterial Nitroreductase Expressing Transgenic Aspen. Environ. Sci. Technol. 2008, 42, 7405–7410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Güngör, T.; Önder, F.C.; Tokay, E.; Gülhan, Ü.G.; Hacıoğlu, N.; Tok, T.T.; Çelik, A.; Köçkar, F.; Ay, M. Prodrugs for nitroreductase based cancer therapy-2: Novel amide/Ntr combinations targeting PC3 cancer cells. Eur. J. Med. Chem. 2019, 171, 383–400. [Google Scholar] [CrossRef]
- Zhang, X.; Li, Q.; You, X.; Zhang, X. Prodrug strategy for cancer cell-specific targeting: A recent overview. Eur. J. Med. Chem. 2017, 139, 542–563. [Google Scholar] [CrossRef]
- Chan-Hyams, J.V.E.; Copp, J.N.; Smaill, J.B.; Patterson, A.V.; Ackerley, D.F. Evaluating the abilities of diverse ni-troaromatic prodrug metabolites to exit a model Gram negative vector for bacterial-directed enzyme-prodrug therapy. Biochem. Pharmacol. 2018, 158, 192–200. [Google Scholar] [CrossRef]
- Ball, P.; Thompson, E.; Anderson, S.; Gwenin, V.; Gwenin, C. Time dependent HPLC analysis of the product ratio of enzymatically reduced prodrug CB1954 by a modified and immobilised nitroreductase. Eur. J. Pharm. Sci. 2019, 127, 217–224. [Google Scholar] [CrossRef] [Green Version]
- Copp, J.N.; Mowday, A.M.; Williams, E.M.; Guise, C.P.; Ashoorzadeh, A.; Sharrock, A.V.; Flanagan, J.U.; Smaill, J.B.; Patterson, A.V.; Ackerley, D.F. Engineering a multifunctional nitroreductase for improved activation of prodrugs and PET probes for cancer gene therapy. Cell Chem. Biol. 2017, 24, 391–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, L.; Hu, F.; Huang, J.; Li, N.; Gu, Y.; Wang, P. A fluorescent turn-on probe for nitroreductase imaging in living cells and tissues under hypoxia conditions. Sens. Actuators B Chem. 2018, 268, 70–76. [Google Scholar] [CrossRef]
- Zhu, K.; Qin, T.; Zhao, C.; Luo, Z.; Huang, Y.; Liu, B.; Wang, L. A novel fluorescent turn-on probe for highly selective detection of nitroreductase in tumor cells. Sens. Actuators B Chem. 2018, 276, 397–403. [Google Scholar] [CrossRef]
- Li, Y.; Deng, Y.; Liu, J.; Fu, J.; Sun, Y.; Ouyang, R.; Miao, Y. A near-infrared frequency upconversion probe for nitroreductase detection and hypoxia tumor in vivo imaging. Sens. Actuators B Chem. 2019, 286, 337–345. [Google Scholar] [CrossRef]
- Kumari, R.; Sunil, D.; Ningthoujam, R.S. Naphthalimides in fluorescent imaging of tumor hypoxia—An up-to-date review. Bioorg. Chem. 2019, 88, 102979. [Google Scholar] [CrossRef]
- Wilkinson, S.R.; Bot, C.; Kelly, J.M.; Hall, B.S. Trypanocidal activity of nitroaromatic prodrugs: Current treatments and future perspectives. Curr. Top. Med. Chem. 2011, 11, 2072–2084. [Google Scholar] [CrossRef] [PubMed]
- Davies, C.; Dey, N.; Negrette, O.S.; Parada, L.A.; Basombrio, M.A.; Garg, N.J. Hepatotoxicity in mice of a novel an-ti-parasite drug candidate hydroxymethylnitrofurazone: A comparison with Benznidazole. PLoS Negl. Trop. Dis. 2014, 8, e3231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De la Calle, M.E.; Cabrera, G.; Cantero, D.; Valle, A.; Bolivar, J. A genetically engineered Escherichia coli strain overexpressing the ni-troreductase NfsB is capable of producing the herbicide D-DIBOA with 100% molar yield. Microb. Cell Fact. 2019, 18, 86. [Google Scholar] [CrossRef] [PubMed]
- Gwenin, C.; Kalaji, M.; Kay, C.M.; Williams, P.A.; Tito, D.N. An in situ amperometric biosensor for the detection of vapours from explosive compounds. Analyst 2008, 133, 621–625. [Google Scholar] [CrossRef]
- Nepali, K.; Lee, H.Y.; Liou, J.P. Nitro-Group-Containing Drugs. J. Med. Chem. 2019, 62, 2851–2893. [Google Scholar] [CrossRef]
- Zenno, S.; Saigo, K.; Kanoh, H.; Inouye, S. Identification of the gene encoding the major NAD(P)H-flavin oxidoreductase of the bioluminescent bacterium Vibrio fischeri ATCC 7744. J. Bacteriol. 1994, 176, 3536–3543. [Google Scholar] [CrossRef] [Green Version]
- Bryant, D.W.; McCalla, D.R.; Leeksma, M.; Laneuville, P. Type I nitroreductases of Escherichia coli. Can. J. Microbiol. 1981, 27, 81–86. [Google Scholar] [CrossRef]
- Roldán, M.D.; Pérez-Reinado, E.; Castillo, F.; Moreno-Vivián, C. Reduction of polynitroaromatic compounds: The bacterial nitroreductases. FEMS Microbiol. Rev. 2008, 32, 474–500. [Google Scholar] [CrossRef] [Green Version]
- Powell, N. Nitroreductase in Cancer Therapy. Available online: https://nitroreductaseincancertreatment.wordpress.com/nitroreductase/ (accessed on 17 December 2020).
- Whiteway, J.; Koziarz, P.; Veall, J.; Sandhu, N.; Kumar, P.; Hoecher, B.; Lambert, I.B. Oxygen-Insensitive Nitroreductases: Analysis of the Roles of nfsA and nfsB in Development of Resistance to 5-Nitrofuran Derivatives in Escherichia coli. J. Bacteriol. 1998, 180, 5529–5539. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Y.; Wu, J.; Liu, H.; Wang, S.; Liu, S.; Zhou, N. Characterization of genes involved in the initial reactions of 4-chloronitrobenzene degradation in Pseudomonasputida ZWL73. Appl. Microbiol. Biotechnol. 2006, 73, 166–171. [Google Scholar] [CrossRef]
- Somerville, C.C.; Nishino, S.F.; Spain, J.C. Isolation and characterization of nitrobenzene nitroreductase from Pseudomonas pseudoalcaligenes JS45. J. Bacteriol. 1995, 177, 3837–3842. [Google Scholar] [CrossRef] [Green Version]
- Liochev, S.I.; Hausladen, A.; Beyer, W.F.; Fridovich, I. NADPH: Ferredoxin oxidoreductase acts as a paraquat diaphorase and is a member of the soxRS regulon. Proc. Natl. Acad. Sci. USA 1994, 91, 1328–1331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liochev, S.I.; Hausladen, A.; Fridovich, I. Nitroreductase A is regulated as a member of the soxRS regulon of Escherichia coli. Proc. Natl. Acad. Sci. USA 1999, 96, 3537–3539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lei, B.; Liu, M.; Huang, S.; Tu, S.C. Vibrio harveyi NADPH-flavin oxidoreductase: Cloning, sequencing and overexpression of the gene and purification and characterization of the cloned enzyme. J. Bacteriol. 1994, 176, 3552–3558. [Google Scholar] [CrossRef] [Green Version]
- Ang, C.W.; Jarrad, A.M.; Cooper, M.; Blaskovich, M.A.T. Nitroimidazoles: Molecular Fireworks That Combat a Broad Spectrum of Infectious Diseases. J. Med. Chem. 2017, 60, 7636–7657. [Google Scholar] [CrossRef] [PubMed]
- WHO Factsheet. Available online: https://www.who.int/en/news-room/fact-sheets/detail/antimicrobial-resistance (accessed on 17 December 2020).
- Crofts, T.S.; Gasparrini, A.J.; Dantas, G. Next-generation approaches to understand and combat the antibiotic resistome. Nat. Rev. Microbiol. 2017, 15, 422–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durand, G.A.; Raoult, D.; Dubourg, G. Antibiotic discovery: History, methods and perspectives. Int. J. Antimicrob. Agents 2019, 53, 371–382. [Google Scholar] [CrossRef]
- Nakamura, S. Structure of Azomycin, a New Antibiotic. Pharm. Bull. 1955, 3, 379–383. [Google Scholar] [CrossRef]
- Edwards, D.I. Nitroimidazole drugs-action and resistance mechanisms I. Mechanism of action. J. Antimicrob. Chemother. 1993, 31, 9–20. [Google Scholar] [CrossRef]
- Patterson, S.; Wyllie, S. Nitro drugs for the treatment of trypanosomatid diseases: Past, present, and future prospects. Trends Parasitol. 2014, 30, 289–298. [Google Scholar] [CrossRef] [Green Version]
- Edwards, D.I. Nitroimidazole drugs—Action and resistance mechanisms II. Mechanisms of resistance. J. Antimicrob. Chemother. 1993, 31, 201–210. [Google Scholar] [CrossRef]
- Samuelson, J. Why Metronidazole is Active against both Bacteria and Parasites. Antimicrob. Agents Chemother. 1999, 43, 1533–1541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bryant, C.; DeLuca, M. Purification and characterization of an oxygen-insensitive NAD(P)H nitroreductase from Enterobacter cloacae. J. Biol. Chem. 1991, 266, 4119–4125. [Google Scholar] [CrossRef]
- Thaker, Y.; Moon, A.; Afzali, A. Helicobacter pylori: A Review of Epidemiology, Treatment, and Management. J. Clin. Gastroenterol. Treat. 2016, 2, 19. [Google Scholar] [CrossRef]
- Fischbach, W.; Goebeler-Kolve, M.E.; Dragosics, B.; Greiner, A.; Stolte, M. Long term outcome of patients with gastric marginal zone B cell lymphoma of mucosa associated lymphoid tissue (MALT) following exclusive Helicobacter pylori eradication therapy: Experience from a large prospective series. Gut 2004, 53, 34–37. [Google Scholar] [CrossRef]
- IARC Working Group on the Evaluation of Carcinogenic Risk to Humans. Schistosomes, Liver Flukes and Helicobacter pylori. In IARC Monographs on the Evaluation of Carcinogenic Risks to Humans; International Agency for Research on Cancer: Lyon, France, 1994; p. 61. [Google Scholar]
- Thung, I.; Aramin, H.; Vavinskaya, V.; Gupta, S.; Park, J.Y.; Crowe, S.E.; Valasek, M.A. Review article: The global emergence of Helicobacter pylori antibiotic resistance. Aliment. Pharmacol. Ther. 2016, 43, 514–533. [Google Scholar] [CrossRef] [Green Version]
- Jeong, J.-Y.; Mukhopadhyay, A.K.; Dailidiene, D.; Wang, Y.; Velapatiño, B.; Gilman, R.H.; Parkinson, A.J.; Nair, G.B.; Wong, B.C.Y.; Lam, S.K.; et al. Sequential Inactivation of rdxA (HP0954) and frxA (HP0642) Nitroreductase Genes Causes Moderate and High-Level Metronidazole Resistance in Helicobacter pylori. J. Bacteriol. 2000, 182, 5082–5090. [Google Scholar] [CrossRef] [Green Version]
- Kwon, D.H.; Hulten, K.; Kato, M.; Kim, J.J.; Lee, M.; El-Zaatari, F.A.K.; Osato, M.S.; Graham, D.Y. DNA Sequence Analysis of rdxA andfrxA from 12 Pairs of Metronidazole-Sensitive and -Resistant Clinical Helicobacter pylori Isolates. Antimicrob. Agents Chemother. 2001, 45, 2609–2615. [Google Scholar] [CrossRef] [Green Version]
- Chua, E.-G.; Debowski, A.W.; Webberley, K.M.; Peters, F.; Lamichhane, B.; Loke, M.-F.; Vadivelu, J.; Tay, C.-Y.; Marshall, B.J.; Wise, M.J. Analysis of core protein clusters identifies candidate variable sites conferring metronidazole resistance in Helicobacter pylori. Gastroenterol. Rep. 2019, 7, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Kwon, D.-H.; El-Zaatari, F.A.K.; Kato, M.; Osato, M.S.; Reddy, R.; Yamaoka, Y.; Graham, D.Y. Analysis of rdxA and Involvement of Additional Genes Encoding NAD(P)H Flavin Oxidoreductase (FrxA) and Ferredoxin-Like Protein (FdxB) in Metronidazole Resistance of Helicobacter pylori. Antimicrob. Agents Chemother. 2000, 44, 2133–2142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Francesco, V.D.; Zullo, A.; Hassan, C.; Giorgio, F.; Rosania, R.; Ierardi, E. Mechanisms of Helicobacter pylori antibiotic resistance: An updated appraisal. World J. Gastrointest. Pathophysiol. 2011, 2, 35–41. [Google Scholar] [CrossRef]
- Lee, S.M.; Kim, N.; Kwon, Y.H.; Nam, R.H.; Kim, J.M.; Park, J.Y.; Lee, Y.S.; Lee, D.H. rdxA, frxA, and efflux pump in metronidazole-resistantHelicobacter pylori: Their relation to clinical outcomes. J. Gastroenterol. Hepatol. 2018, 33, 681–688. [Google Scholar] [CrossRef] [PubMed]
- Reysset, G. Genetics of 5-Nitroimidazole Resistance inBacteroidesSpecies. Anaerobe 1996, 2, 59–69. [Google Scholar] [CrossRef]
- Haggoud, A.; Reysset, G.; Azeddoug, H.; Sebald, M. Nucleotide sequence analysis of two 5-nitroimidazole resistance determinants from Bacteroides strains and of a new insertion sequence upstream of the two genes. Antimicrob. Agents Chemother. 1994, 38, 1047–1051. [Google Scholar] [CrossRef] [Green Version]
- Carlier, J.P.; Sellier, N.; Rager, M.N.; Reysset, G. Metabolism of a 5-nitroimidazole in susceptible and resistant isogenic strains of Bacteroides fragilis. Antimicrob. Agents Chemother. 1997, 41, 1495–1499. [Google Scholar] [CrossRef] [Green Version]
- Alauzet, C.; Lozniewski, A.; Marchandin, H. Metronidazole resistance and nim genes in anaerobes: A review. Anaerobe 2019, 55, 40–53. [Google Scholar] [CrossRef] [PubMed]
- Dingsdag, S.A.; Hunter, N. Metronidazole: An update on metabolism, structure–cytotoxicity and resistance mechanisms. J. Antimicrob. Chemother. 2018, 73, 265–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meggersee, R.; Abratt, V. The occurrence of antibiotic resistance genes in drug resistant Bacteroides fragilis isolates from Groote Schuur Hospital, South Africa. Anaerobe 2015, 32, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Husain, F.; Veeranagouda, Y.; Hsi, J.; Meggersee, R.; Abratt, V.; Wexler, H.M. Two Multidrug-Resistant Clinical Isolates of Bacteroides fragilis Carry a Novel Metronidazole ResistancenimGene (nimJ). Antimicrob. Agents Chemother. 2013, 57, 3767–3774. [Google Scholar] [CrossRef] [Green Version]
- Gal, M.; Brazier, J.S. Metronidazole resistance in Bacteroides spp. carrying nim genes and the selection of slow-growing metronidazole-resistant mutants. J. Antimicrob. Chemother. 2004, 54, 109–116. [Google Scholar] [CrossRef] [Green Version]
- Stanko, A.P.; Sóki, J.; Brkić, D.V.; Plečko, V. Lactate dehydrogenase activity in Bacteroides fragilis group strains with induced resistance to metronidazole. J. Glob. Antimicrob. Resist. 2016, 5, 11–14. [Google Scholar] [CrossRef]
- Theron, M.M.; Van Rensburg, M.N.J.; Chalkley, L.J. Nitroimidazole resistance genes (nimB) in anaerobic Gram-positive cocci (previously Peptostreptococcus spp.). J. Antimicrob. Chemother. 2004, 54, 240–242. [Google Scholar] [CrossRef] [Green Version]
- Kullin, B.; Brock, T.; Rajabally, N.; Anwar, F.; Vedantam, G.; Reid, S.; Abratt, V. Characterisation of Clostridium difficile strains isolated from Groote Schuur Hospital, Cape Town, South Africa. Eur. J. Clin. Microbiol. Infect. Dis. 2016, 35, 1709–1718. [Google Scholar] [CrossRef]
- Sóki, J.; Hedberg, M.; Patrick, S.; Bálint, B.; Herczeg, R.; Nagy, I.; Hecht, D.W.; Nagy, E.; Urbán, E. Emergence and evolution of an international cluster of MDR Bacteroides fragilis isolates. J. Antimicrob. Chemother. 2016, 71, 2441–2448. [Google Scholar] [CrossRef] [Green Version]
- Sethi, S.; Shukla, R.; Bala, K.; Gautam, V.; Angrup, A.; Ray, P. Emerging metronidazole resistance in Bacteroides spp. and its association with the nim gene: A study from North India. J. Glob. Antimicrob. Resist. 2019, 16, 210–214. [Google Scholar] [CrossRef]
- Leitsch, D.; Sóki, J.; Kolarich, D.; Urbán, E.; Nagy, E. A study on Nim expression in Bacteroides fragilis. Microbiology 2014, 160, 616–622. [Google Scholar] [CrossRef] [Green Version]
- Ank, N.; Sydenham, T.V.; Iversen, L.H.; Justesen, U.S.; Wang, M. Characterisation of a multidrug-resistant Bacteroides fragilis isolate recovered from blood of a patient in Denmark using whole-genome sequencing. Int. J. Antimicrob. Agents 2015, 46, 117–120. [Google Scholar] [CrossRef] [PubMed]
- Sadarangani, S.P.; Cunningham, S.A.; Jeraldo, P.R.; Wilson, J.W.; Khare, R.; Patel, R. Metronidazole- and car-bapenem-resistant bacteroides thetaiotaomicron isolated in Rochester, Minnesota, in 2014. Antimicrob. Agents Chemother. 2015, 59, 4157–4161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gajdács, M.; Spengler, G.; Urbán, E. Identification and Antimicrobial Susceptibility Testing of Anaerobic Bacteria: Ru-bik’s Cube of Clinical Microbiology? Antibiotics 2017, 6, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukherjee, T.; Boshoff, H. Nitroimidazoles for the treatment of TB: Past, present and future. Future Med. Chem. 2011, 3, 1427–1454. [Google Scholar] [CrossRef] [Green Version]
- WHO Global Tuberculosis Report. 2019. Available online: https://apps.who.int/iris/bitstream/handle/10665/329368/9789241565714-eng.pdf?ua=1 (accessed on 17 December 2020).
- Boshoff, H.I.; Barry, C.E. Is the mycobacterial cell wall a hopeless drug target for latent tuberculosis? Drug Discov. Today Dis. Mech. 2006, 3, 237–245. [Google Scholar] [CrossRef]
- Singh, R.; Manjunatha, U.; Boshoff, H.I.M.; Ha, Y.H.; Niyomrattanakit, P.; Ledwidge, R.; Dowd, C.S.; Lee, I.Y.; Kim, P.; Zhang, L.; et al. PA-824 Kills Nonreplicating Mycobacterium tuberculosis by Intracellular NO Release. Science 2008, 322, 1392–1395. [Google Scholar] [CrossRef] [Green Version]
- Manjunatha, U.H.; Boshoff, H.; Dowd, C.S.; Zhang, L.; Albert, T.J.; Norton, J.E.; Daniels, L.; Dick, T.; Pang, S.S.; Barry, C.E. Identification of a nitroimidazo-oxazine-specific protein involved in PA-824 resistance in Mycobacterium tuberculosis. Proc. Natl. Acad. Sci. USA 2005, 103, 431–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Matsumoto, M.; Ishida, H.; Ohguro, K.; Yoshitake, M.; Gupta, R.; Geiter, L.; Hafkin, J. Delamanid: From discovery to its use for pulmonary multidrug-resistant tuber-culosis (MDR-TB). Tuberculosis (Edinb.) 2018, 111, 20–30. [Google Scholar] [CrossRef]
- Xavier, A.S.; Lakshmanan, M. Delamanid: A new armor in combating drug-resistant tuberculosis. J. Pharmacol. Pharmacother. 2014, 5, 222–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO. Position Statement on the Use of Delamanid for Multidrug-Resistant Tuberculosis. Available online: https://www.who.int/tb/publications/2018/Position_Paper_Delamanid/en/ (accessed on 17 December 2020).
- Stinson, K.; Kurepina, N.; Venter, A.; Fujiwara, M.; Kawasaki, M.; Timm, J.; Shashkina, E.; Kreiswirth, B.N.; Liu, Y.; Matsumoto, M.; et al. MIC of Delamanid (OPC-67683) against Mycobacterium tuberculosis Clinical Isolates and a Proposed Critical Concentration. Antimicrob. Agents Chemother. 2016, 60, 3316–3322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanaki, E.; Hayashi, M.; Matsumoto, M. Delamanid is not metabolized by Salmonella or human nitroreductases: A pos-sible mechanism for the lack of mutagenicity. Regul. Toxicol. Pharmacol. 2017, 84, 1–8. [Google Scholar] [CrossRef]
- Diacon, A.H.; Dawson, R.; Hanekom, M.; Narunsky, K.; Venter, A.; Hittel, N.; Geiter, L.J.; Wells, C.D.; Paccaly, A.J.; Donald, P.R. Early bactericidal activity of delamanid (OPC-67683) in smear-positive pulmonary tuberculosis patients. Int. J. Tuberc. Lung Dis. 2011, 15, 949–954. [Google Scholar] [CrossRef] [PubMed]
- Sloan, D.J.; Lewis, J.M. The role of delamanid in the treatment of drug-resistant tuberculosis. Ther. Clin. Risk Manag. 2015, 11, 779–791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skripconoka, V.; Danilovits, M.; Pehme, L.; Tomson, T.; Skenders, G.; Kummik, T.; Cirule, A.; Leimane, V.; Kurve, A.; Levina, K.; et al. Delamanid improves outcomes and reduces mortality in multi-drug-resistant tuberculosis. Eur. Respir. J. 2013, 41, 1393–1400. [Google Scholar] [CrossRef] [Green Version]
- Esposito, S.; Bosis, S.; Tadolini, M.; Bianchini, S.; Migliori, G.B.; Principi, N. Efficacy, safety, and tolerability of a 24-month treatment regimen including delamanid in a child with extensively drug-resistant tuberculosis. Medicine (Baltimore) 2016, 95, e5347. [Google Scholar] [CrossRef]
- Wayne, L.G.; Hayes, L.G. An in vitro model for sequential study of shiftdown of Mycobacterium tuberculosis through two stages of nonreplicating persistence. Infect. Immun. 1996, 64, 2062–2069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wayne, L.G. In Vitro Model of Hypoxically Induced Nonreplicating Persistence of Mycobacterium tuberculosis. Methods Mol. Med. 2001, 54, 247–269. [Google Scholar] [CrossRef]
- Upton, A.M.; Cho, S.; Yang, T.J.; Kim, Y.; Wang, Y.; Lu, Y.; Wang, B.; Xu, J.; Mdluli, K.; Ma, Z.; et al. In vitro and in vivo activities of the nitroimidazole TBA-354 against Mycobacterium tuberculosis. Antimicrob. Agents Chemother. 2015, 59, 136–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Hashizume, H.; Tomishige, T.; Nakamura, I.; Matsuba, M.; Fujiwara, M.; Kitamoto, R.; Hanaki, E.; Ohba, Y.; Matsumoto, M. Delamanid kills dormant myco-bacteria in vitro and in a Guinea pig model of tuberculosis. Antimicrob. Agents Chemother. 2017, 61, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumoto, M.; Hashizume, H.; Tomishige, T.; Kawasaki, M.; Tsubouchi, H.; Sasaki, H.; Shimokawa, Y.; Komatsu, M. OPC-67683, a nitro-dihydro-imidazooxazole derivative with prom-ising action against tuberculosis in vitro and in mice. PLoS Med. 2006, 3, e466. [Google Scholar] [CrossRef]
- Zhang, Y.; Yew, W.-W. Mechanisms of drug resistance in Mycobacterium tuberculosis: Update 2015. Int. J. Tuberc. Lung Dis. 2015, 19, 1276–1289. [Google Scholar] [CrossRef]
- Bashiri, G.; Rehan, A.M.; Greenwood, D.R.; Dickson, J.M.J.; Baker, E.N. Metabolic Engineering of Cofactor F420 Production in Mycobacterium smegmatis. PLoS ONE 2010, 5, e15803. [Google Scholar] [CrossRef] [Green Version]
- Choi, K.-P.; Kendrick, N.; Daniels, L. Demonstration that fbiC Is Required by Mycobacterium bovis BCG for Coenzyme F420 and FO Biosynthesis. J. Bacteriol. 2002, 184, 2420–2428. [Google Scholar] [CrossRef] [Green Version]
- Fujiwara, M.; Kawasaki, M.; Hariguchi, N.; Liu, Y.; Matsumoto, M. Mechanisms of resistance to delamanid, a drug for Mycobacterium tuberculosis. Tuberculosis 2018, 108, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Masto, B. Not-for-profit to launch antibiotic against drug-resistant tuberculosis. Nat. Biotechnol. 2019. [Google Scholar] [CrossRef]
- Haver, H.L.; Chua, A.; Ghode, P.; Lakshminarayana, S.B.; Singhal, A.; Mathema, B.; Wintjens, R.; Bifani, P. Mutations in genes for the F420 biosynthetic pathway and a nitroreductase enzyme are the primary resistance determinants in spontaneous in vitro-selected PA-824-resistant mutants of Mycobacterium tuberculosis. Antimicrob. Agents Chemother. 2015, 59, 5316–5323. [Google Scholar] [CrossRef] [Green Version]
- Li, S.-Y.; Tasneen, R.; Tyagi, S.; Soni, H.; Converse, P.J.; Mdluli, K.; Nuermberger, E.L. Bactericidal and Sterilizing Activity of a Novel Regimen with Bedaquiline, Pretomanid, Moxifloxacin, and Pyrazinamide in a Murine Model of Tuberculosis. Antimicrob. Agents Chemother. 2017, 61, e00913-17. [Google Scholar] [CrossRef] [Green Version]
- Dawson, R.; Diacon, A.H.; Everitt, D.; Van Niekerk, C.; Donald, P.R.; Burger, D.A.; Schall, R.; Spigelman, M.; Conradie, A.; Eisenach, K.; et al. Efficiency and safety of the combination of moxifloxacin, pretomanid (PA-824), and pyrazinamide during the first 8 weeks of antituberculosis treatment: A phase 2b, open-label, partly randomised trial in patients with drug-susceptible or drug-resistant pulmonary tuberculosis. Lancet 2015, 385, 1738–1747. [Google Scholar] [CrossRef] [PubMed]
- Stover, C.K.; Warrener, P.; Van Devanter, D.R.; Sherman, D.R.; Arain, T.M.; Langhorne, M.H.; Anderson, S.W.; Towell, J.A.; Yuan, Y.; McMurray, D.N.; et al. A small-molecule nitroimidazopyran drug candidate for the treatment of tuberculosis. Nat. Cell Biol. 2000, 405, 962–966. [Google Scholar] [CrossRef]
- Greening, C.; Ahmed, F.H.; Mohamed, A.E.; Lee, B.M.; Pandey, G.; Warden, A.C.; Scott, C.; Oakeshott, J.G.; Taylor, M.C.; Jackson, C.J. Physiology, Biochemistry, and Applications of F420- and Fo-Dependent Redox Reactions. Microbiol. Mol. Biol. Rev. 2016, 80, 451–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Purwantini, E.; Mukhopadhyay, B. Conversion of NO2 to NO by reduced coenzyme F420 protects mycobacteria from nitrosative damage. Proc. Natl. Acad. Sci. USA 2009, 106, 6333–6338. [Google Scholar] [CrossRef] [Green Version]
- Gurumurthy, M.; Rao, M.; Mukherjee, T.; Rao, S.P.S.; Boshoff, H.I.; Dick, T.; Barry, C.E., 3rd; Manjunatha, U.H. A novel F(420)-dependent anti-oxidant mechanism protects Mycobacterium tuberculosis against oxidative stress and bactericidal agents. Mol. Microbiol. 2013, 87, 744–755. [Google Scholar] [CrossRef]
- Aharoni, A.; Gaidukov, L.; Khersonsky, O.; Gould, S.M.; Roodveldt, C.; Tawfik, D.S. The ’evolvability’ of promiscuous protein functions. Nat. Genet. 2004, 37, 73–76. [Google Scholar] [CrossRef]
- Lee, B.M.; Harold, L.K.; Almeida, D.V.; Afriat-Jurnou, L.; Aung, H.L.; Forde, B.M.; Hards, K.; Pidot, S.J.; Ahmed, F.H.; Mohamed, A.E.; et al. Predicting nitroimidazole antibiotic resistance mutations in Mycobacterium tuberculosis with protein engineering. PLoS Pathog. 2020, 16, e1008287. [Google Scholar] [CrossRef] [Green Version]
- Schena, E.; Nedialkova, L.; Borroni, E.; Battaglia, S.; Cabibbe, A.M.; Niemann, S.; Utpatel, C.; Merker, M.; Trovato, A.; Hofmann-Thiel, S.; et al. Delamanid susceptibility testing of Mycobacterium tuberculosis using the resazurin microtitre assay and the BACTEC™ MGIT™ 960 system. J. Antimicrob. Chemother. 2016, 71, 1532–1539. [Google Scholar] [CrossRef] [Green Version]
- Ryan, A. Azoreductases in drug metabolism. Br. J. Pharmacol. 2017, 174, 2161–2173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dinos, G.P.; Athanassopoulos, C.M.; Missiri, D.A.; Giannopoulou, P.C.; Vlachogiannis, I.A.; Papadopoulos, G.E.; Papaioannou, D.; Kalpaxis, D.L. Chloramphenicol Derivatives as Antibacterial and Anticancer Agents: Historic Problems and Current Solutions. Antibiotics 2016, 5, 20. [Google Scholar] [CrossRef]
- Rahim, N.A.; Cheah, S.; Johnson, M.D.; Yu, H.; Sidjabat, H.E.; Boyce, J.; Butler, M.S.; Cooper, M.A.; Fu, J.; Paterson, D.L.; et al. Synergistic killing of NDM-producing MDR Klebsiella pneumoniae by two ’old’ antibiotics-polymyxin B and chloramphenicol. J. Antimicrob. Chemother. 2015, 70, 2589–2597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shu, X.; Linet, M.; Gao, R.; Gao, Y.; Brinton, L.; Jin, F.; Fraumeni, J. Chloramphenicol use and childhood leukaemia in Shanghai. Lancet 1987, 330, 934–937. [Google Scholar] [CrossRef]
- Lim, C.; Takahashi, E.; Hongsuwan, M.; Wuthiekanun, V.; Thamlikitkul, V.; Hinjoy, S.; Day, N.P.; Peacock, S.J.; Limmathurotsakul, D. Epidemiology and burden of multidrug-resistant bacterial infection in a developing country. eLife 2016, 5, e18082. [Google Scholar] [CrossRef] [PubMed]
- Sood, S. Chloramphenicol—A Potent Armament Against Multi-Drug Resistant (MDR) Gram Negative Bacilli? J. Clin. Diagn. Res. 2016, 10, DC01-3. [Google Scholar] [CrossRef] [PubMed]
- Crofts, T.S.; Sontha, P.; King, A.O.; Wang, B.; Biddy, B.A.; Zanolli, N.; Gaumnitz, J.; Dantas, G. Discovery and character-isation of a nitroreductase capable of conferring bacterial resistance to chloramphenicol. Cell Chem. Biol. 2019, 26, 559–570. [Google Scholar] [CrossRef]
- Eliakim-Raz, N.; Lador, A.; Leibovici-Weissman, Y.; Elbaz, M.; Paul, M.; Leibovici, L. Efficacy and safety of chloramphenicol: Joining the revival of old antibiotics? Systematic review and meta-analysis of randomized controlled trials. J. Antimicrob. Chemother. 2015, 70, 979–996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menzel, T.M.; Tischer, M.; François, P.; Nickel, J.; Schrenzel, J.; Bruhn, H.; Albrecht, A.; Lehmann, L.; Holzgrabe, U.; Ohlsen, K. Mode-of-action studies of the novel bisquaternary bisnaphthalimide MT02 against Staphylococcus aureus. Antimicrob. Agents Chemother. 2011, 55, 311–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatterjee, S.S.; Otto, M. Improved understanding of factors driving methicillin-resistant Staphylococcus aureus epidemic waves. Clin. Epidemiol. 2013, 5, 205–217. [Google Scholar]
- Lowy, F.D. Staphylococcus aureus Infections. N. Engl. J. Med. 1998, 339, 520–532. [Google Scholar] [CrossRef]
- Shanson, D. Antibiotic-resistant Staphylococcus aureus. J. Hosp. Infect. 1981, 2, 11–36. [Google Scholar] [CrossRef]
- Smith, J.T.; Amyes, S.G.B. Bacterial resistance to antifolate chemotherapeutic agents mediated by Plasmids. Br. Med. Bull. 1984, 40, 42–46. [Google Scholar] [CrossRef] [PubMed]
- Jevons, M.P. “Celbenin”—Resistant Staphylococci. BMJ 1961, 1, 124–125. [Google Scholar] [CrossRef]
- El-Hossary, E.M.; Förstner, K.U.; François, P.; Baud, D.; Streker, K.; Schrenzel, J.; Ohlsen, K.; Holzgrabe, U. A Novel Mechanism of Inactivating Antibacterial Nitro Compounds in the Human Pathogen Staphylococcus aureus by Overexpression of a NADH-Dependent Flavin Nitroreductase. Antimicrob. Agents Chemother. 2018, 62, e01510-17. [Google Scholar] [CrossRef] [Green Version]
- Tejman-Yarden, N.; Millman, M.; Lauwaet, T.; Davids, B.J.; Gillin, F.D.; Dunn, L.; Upcroft, J.A.; Miyamoto, Y.; Eckmann, L. Impaired parasite attachment as fitness cost of metronidazole re-sistance in Giardia lamblia. Antimicrob. Agents Chemother. 2011, 55, 4643–4651. [Google Scholar] [CrossRef] [Green Version]
- Njoroge, M.; Njuguna, N.M.; Mutai, P.; Ongarora, D.S.B.; Smith, P.W.; Chibale, K. Recent Approaches to Chemical Discovery and Development Against Malaria and the Neglected Tropical Diseases Human African Trypanosomiasis and Schistosomiasis. Chem. Rev. 2014, 114, 11138–11163. [Google Scholar] [CrossRef]
- Allarakhia, M. Open-source approaches for the repurposing of existing or failed candidate drugs: Learning from and applying the lessons across diseases. Drug Des. Dev. Ther. 2013, 7, 753–766. [Google Scholar] [CrossRef] [Green Version]
- Çelik, A.; Yetiş, G.; Ay, M.; Güngör, T. Modification of existing antibiotics in the form of precursor prodrugs that can be subsequently activated by nitroreductases of the target pathogen. Bioorg. Med. Chem. Lett. 2016, 26, 4057–4060. [Google Scholar] [CrossRef]
- Drawz, S.M.; Papp-Wallace, K.M.; Bonomo, R.A. New β-Lactamase Inhibitors: A Therapeutic Renaissance in an MDR World. Antimicrob. Agents Chemother. 2014, 58, 1835–1846. [Google Scholar] [CrossRef] [Green Version]
- Sander, C.; McShane, H. Translational Mini-Review Series on Vaccines: Development and evaluation of improved vaccines against tuberculosis. Clin. Exp. Immunol. 2007, 147, 401–411. [Google Scholar] [CrossRef] [PubMed]
- Kaufmann, S.H.; Gengenbacher, M. Recombinant live vaccine candidates against tuberculosis. Curr. Opin. Biotechnol. 2012, 23, 900–907. [Google Scholar] [CrossRef]
- Kwon, K.W.; Kim, W.S.; Kim, H.; Han, S.J.; Hahn, M.-Y.; Lee, J.S.; Nam, K.T.; Cho, S.-N.; Shin, S.J. Novel vaccine potential of Rv3131, a DosR regulon-encoded putative nitroreductase, against hyper-virulent Mycobacterium tuberculosis strain K. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The reduction scheme of a nitro group and the respective electron transfers required [20]. NAD: nicotinamide adenine dinucleotide and NADPH: nicotinamide adenine dinucleotide phosphate.
Figure 1.
The reduction scheme of a nitro group and the respective electron transfers required [20]. NAD: nicotinamide adenine dinucleotide and NADPH: nicotinamide adenine dinucleotide phosphate.

Figure 2.
Mutagenic pathway of nitroarenes [33].
Figure 2.
Mutagenic pathway of nitroarenes [33].

Figure 3.
Mechanism of metronidazole [27] involving bioreduction of the nitro group by ferredoxin.
Figure 3.
Mechanism of metronidazole [27] involving bioreduction of the nitro group by ferredoxin.

Figure 4.
Summarised mode of action and main mechanisms involved in resistance. (1) Shows nitroimidazole reductase activity encoded by the nim genes, (2) metabolic shift away to the pathway related to conversion of pyruvate to lactate via lactate dehydrogenase (3) increased efflux of the antibiotic (4) increased DNA repair capacity (5) activation of antioxidant defense systems (6) deficiency of the ferrous iron transporter FeoAB, (7) overexpression of the rhamnose catabolism regulatory protein RhaR [50].
Figure 4.
Summarised mode of action and main mechanisms involved in resistance. (1) Shows nitroimidazole reductase activity encoded by the nim genes, (2) metabolic shift away to the pathway related to conversion of pyruvate to lactate via lactate dehydrogenase (3) increased efflux of the antibiotic (4) increased DNA repair capacity (5) activation of antioxidant defense systems (6) deficiency of the ferrous iron transporter FeoAB, (7) overexpression of the rhamnose catabolism regulatory protein RhaR [50].

Figure 5.
Structure of delamanid [69].
Figure 5.
Structure of delamanid [69].

Figure 6.
Gene products involved in the bioactivation of delamanid [86].
Figure 6.
Gene products involved in the bioactivation of delamanid [86].

Figure 7.
Structure of pretomanid [91].
Figure 7.
Structure of pretomanid [91].

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Thomas, C.; Gwenin, C.D. The Role of Nitroreductases in Resistance to Nitroimidazoles. Biology 2021, 10, 388. https://doi.org/10.3390/biology10050388
AMA Style
Thomas C, Gwenin CD. The Role of Nitroreductases in Resistance to Nitroimidazoles. Biology. 2021; 10(5):388. https://doi.org/10.3390/biology10050388
Chicago/Turabian StyleThomas, Carol, and Christopher D. Gwenin. 2021. "The Role of Nitroreductases in Resistance to Nitroimidazoles" Biology 10, no. 5: 388. https://doi.org/10.3390/biology10050388
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.