

Role of a 49 kDa Trypanosoma cruzi Mucin-Associated Surface Protein (MASP49) during the Infection Process and Identification of a Mammalian Cell Surface Receptor
 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Parasites
2.2. Peritoneal Macrophages
2.3. Rat Cardiomyocytes
2.4. MASP Peptide and Antiserum Production
2.5. Total Protein Extract
2.6. MASP49 Immune Detection
2.7. Indirect Immunofluorescence (IFI) Assay
2.8. Membrane Enriched Fraction
2.9. MASP49 Purification
2.10. Phosphatidylinositol-Specific Phospholipase C (PLC) Treatment
2.11. In Vitro Inhibition Infection Assay
2.12. Binding Assay
2.13. Glycoprotein Detection
2.14. MASP Binding Inhibition via Carbohydrates
2.15. The Colocalization of MASP49 and mMGL
2.16. Animal Ethical Management
3. Results
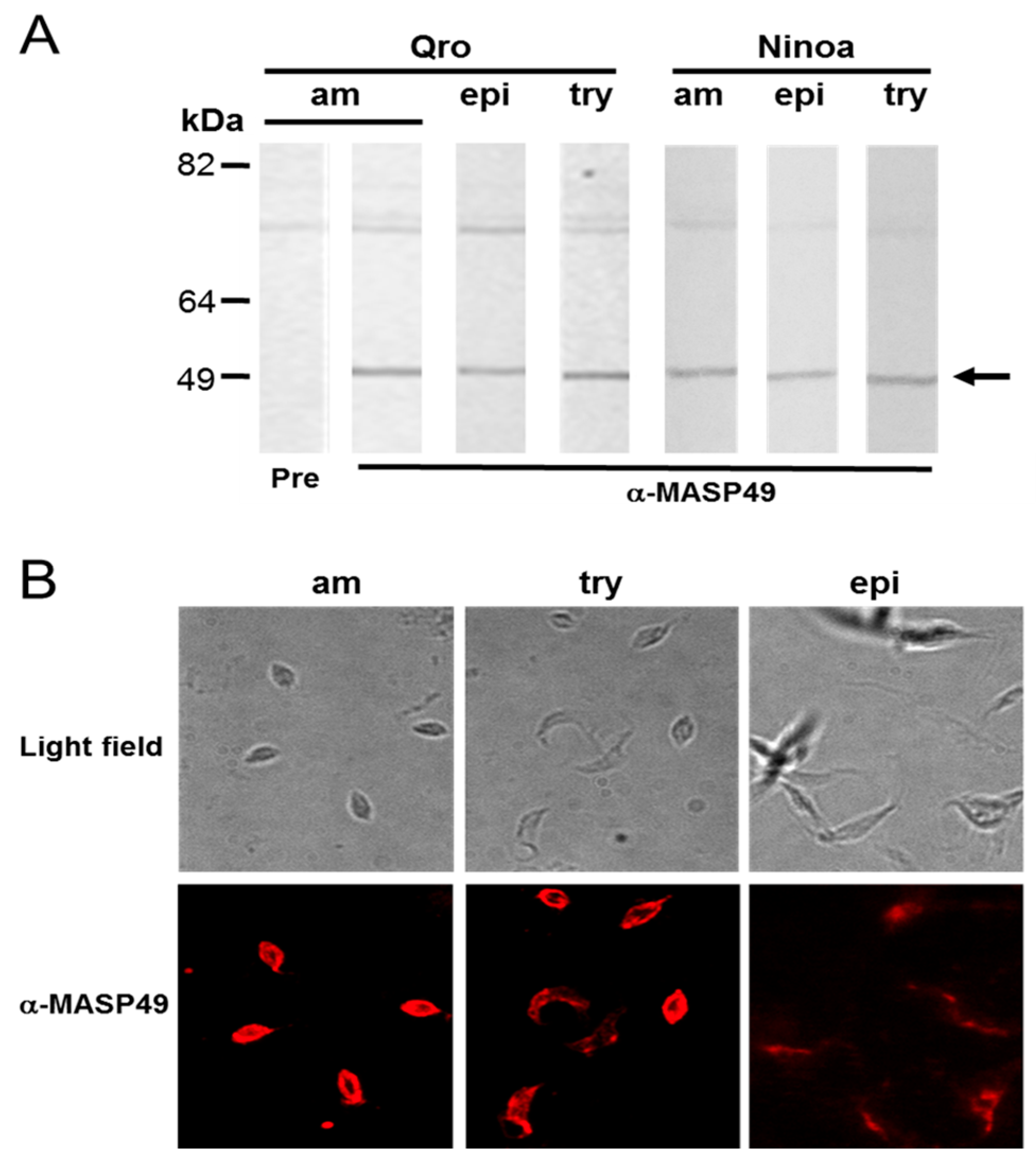
3.1. MASP49 Is Expressed in All Stages of T. cruzi
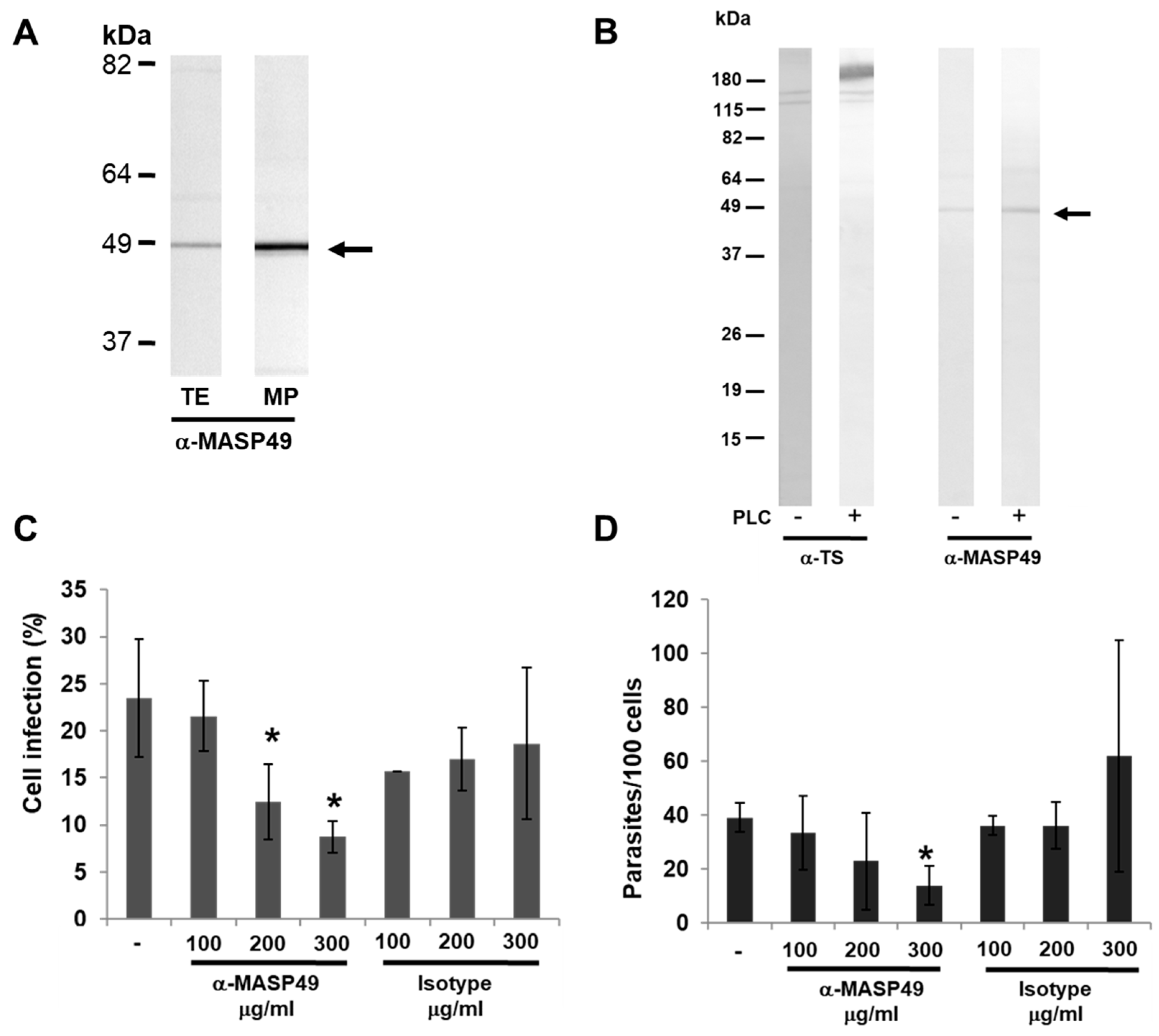
3.2. MASP49 Is GPI-Anchored to the Membrane of the Parasite
3.3. MASP49 in Mammalian Infection
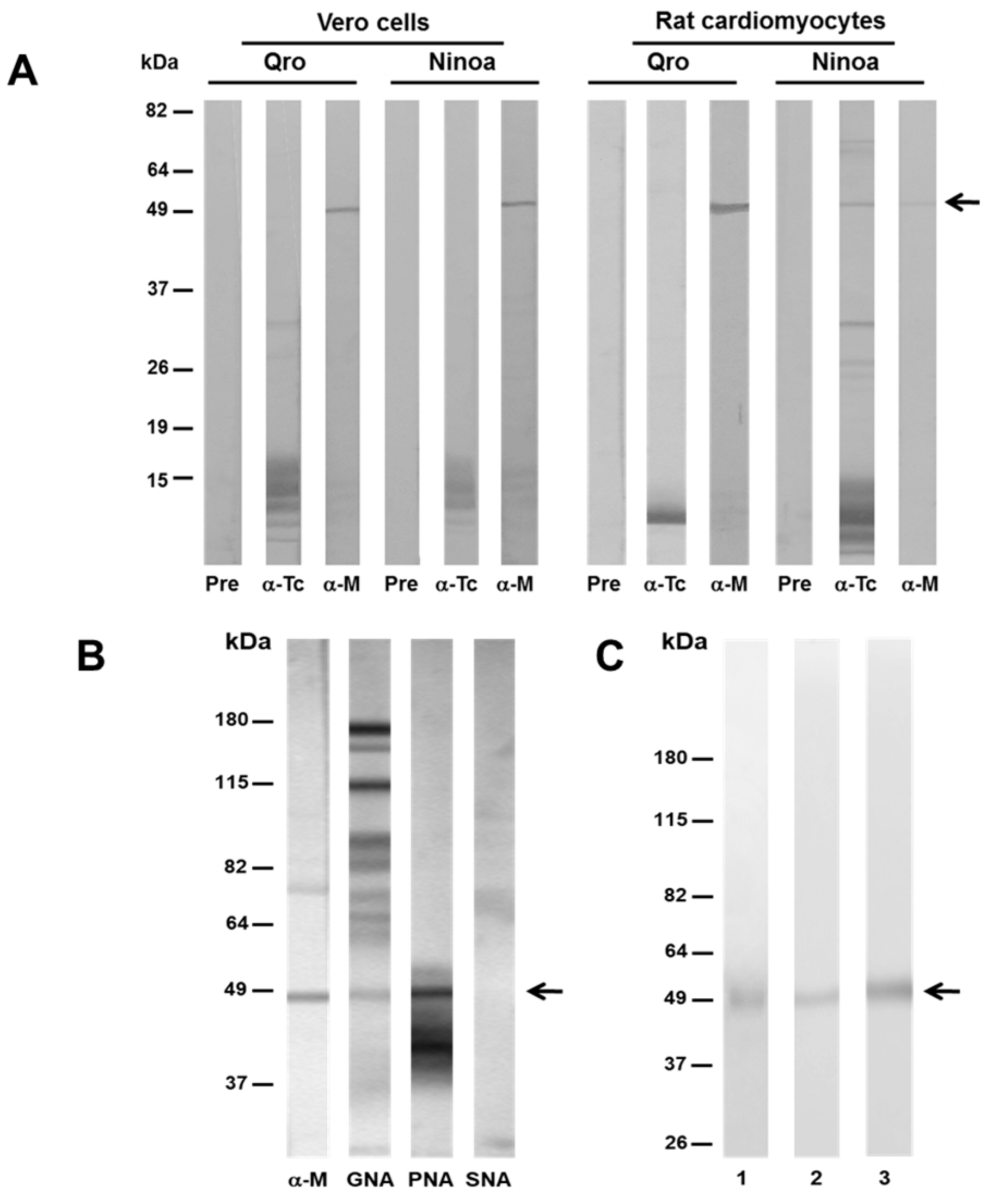
3.4. MASP49 Binds to Different Host Cells
3.5. MASP49 Is Glycosylated with Galactose (1-3) N-Acetylgalactosamine
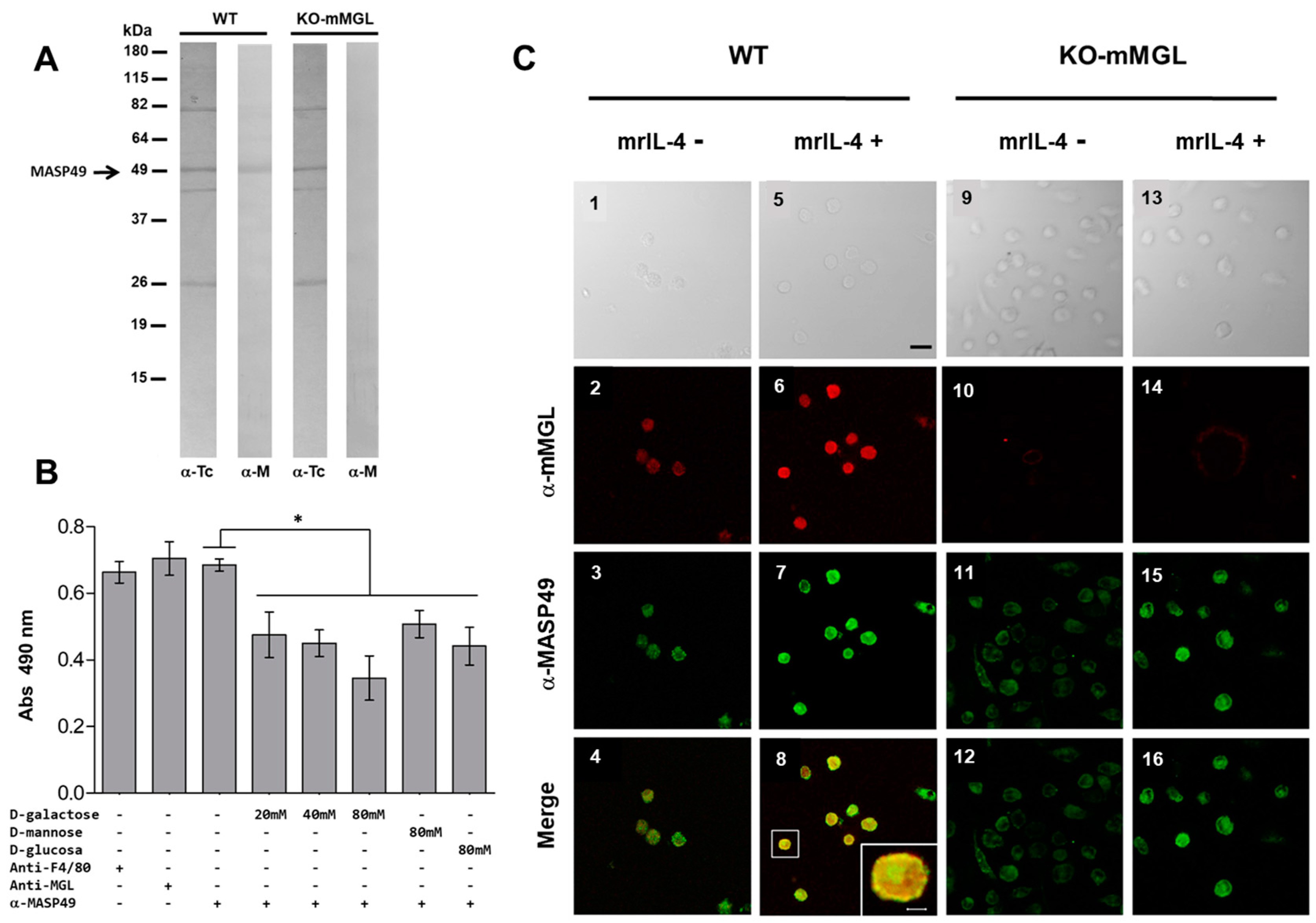
3.6. MASP49 Binds to mMGL, and Galactose Reduces This Binding
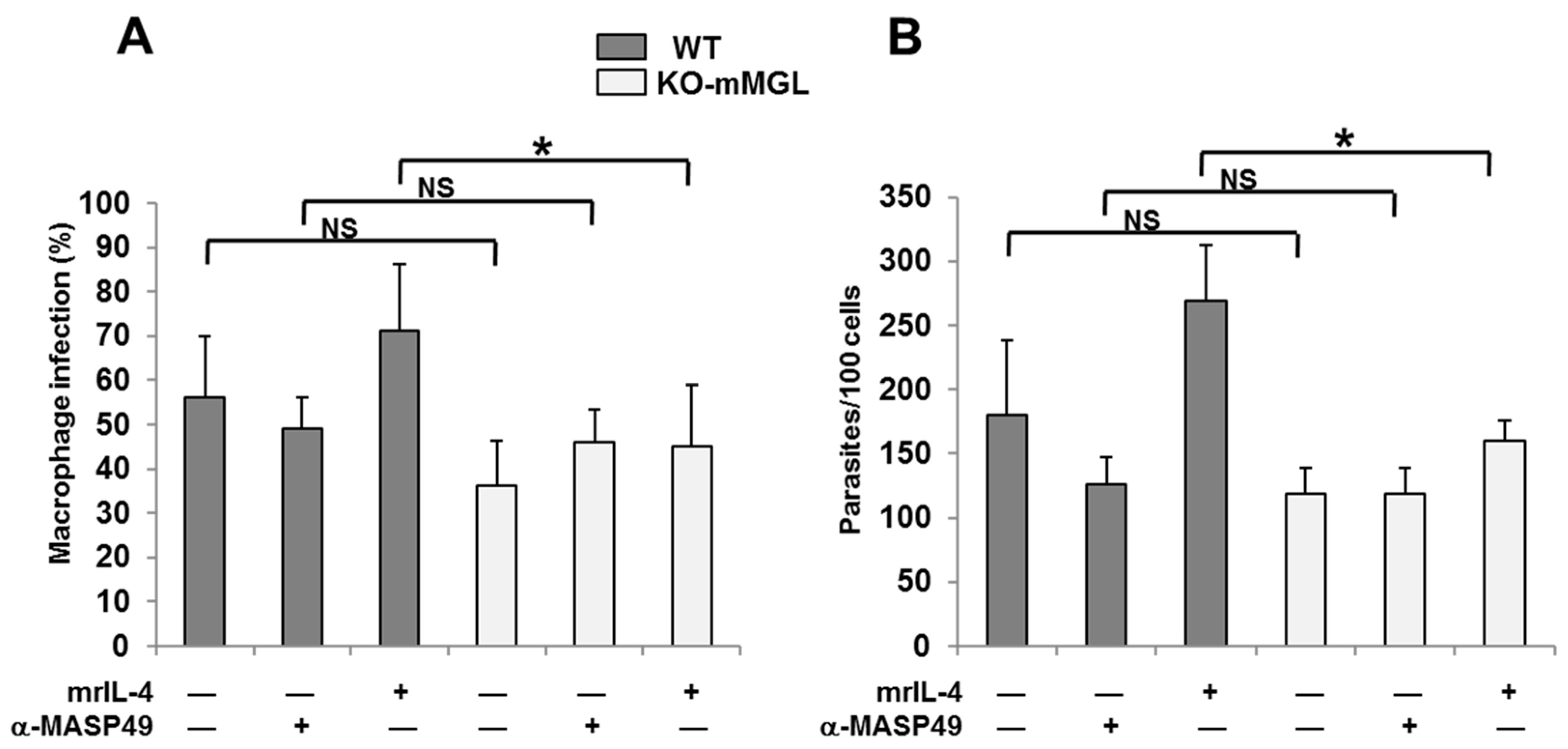
3.7. The Absence of mMGL Reduces Peritoneal Macrophage Infection
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. Chagas disease (also known as American trypanosomiasis). Fact Sheet, World Health Organization. Available online: https://www.who.int/news-room/fact-sheets/detail/chagas-disease-(american-trypanosomiasis) (accessed on 2 October 2022).
- Torhorst, C.W.; White, Z.S.; Bhosale, C.R.; Beatty, N.L.; Wisely, S.M. Identification of the parasite, Trypanosoma cruzi, in multiple tissues of epidemiological significance in the Virginia opossum (Didelphis virginiana): Implications for environmental and vertical transmission routes. PLoS Negl. Trop. Dis. 2022, 16, e0010974. [Google Scholar] [CrossRef] [PubMed]
- Onofre, T.S.; Loch, L.; Ferreira Rodrigues, J.P.; Macedo, S.; Yoshida, N. Gp35/50 mucin molecules of Trypanosoma cruzi metacyclic forms that mediate host cell invasion interact with annexin A2. PLoS Negl. Trop. Dis. 2022, 16, e0010788. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, N.M.; Myler, P.J.; Bartholomeu, D.C.; Nilsson, D.; Aggarwal, G.; Tran, A.N.; Ghedin, E.; Worthey, E.A.; Delcher, A.L.; Blandin, G.; et al. The genome sequence of Trypanosoma cruzi, etiologic agent of Chagas disease. Science 2005, 309, 409–415. [Google Scholar] [CrossRef] [Green Version]
- Franzén, O.; Ochaya, S.; Sherwood, E.; Lewis, M.D.; Llewellyn, M.S.; Miles, M.A.; Andersson, B. Shotgun sequencing analysis of Trypanosoma cruzi I Sylvio X10/1 and comparison with T. cruzi VI CL Brener. PLoS Negl. Trop. Dis. 2011, 5, e984. [Google Scholar] [CrossRef] [Green Version]
- Leão, A.C.; Viana, L.A.; Fortes de Araujo, F.; de Lourdes Almeida, R.; Freitas, L.M.; Coqueiro-Dos-Santos, A.; da Silveira-Lemos, D.; Cardoso, M.S.; Reis-Cunha, J.L.; Teixeira-Carvalho, A.; et al. Antigenic diversity of MASP gene family of Trypanosoma cruzi. Microbes Infect. 2022, 24, 104982. [Google Scholar] [CrossRef] [PubMed]
- Atwood, J.A., 3rd; Minning, T.; Ludolf, F.; Nuccio, A.; Weatherly, D.B.; Alvarez-Manilla, G.; Tarleton, R.; Orlando, R. Glycoproteomics of Trypanosoma cruzi trypomastigotes using subcellular fractionation, lectin affinity, and stable isotope labeling. J. Proteome Res. 2006, 5, 3376–3384. [Google Scholar] [CrossRef] [PubMed]
- Queiroz, R.M.; Ricart, C.A.; Machado, M.O.; Bastos, I.M.; de Santana, J.M.; de Sousa, M.V.; Roepstorff, P.; Charneau, S. Insight into the exoproteome of the tissue-Derived trypomastigote form of Trypanosoma cruzi. Front. Chem. 2016, 4, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandacaru, S.C.; Queiroz, R.M.L.; Alborghetti, M.R.; de Oliveira, L.S.; de Lima, C.M.R.; Bastos, I.M.D.; Santana, J.M.; Roepstorff, P.; Ricart, C.A.O.; Charneau, S. Exoproteome profiling of Trypanosoma cruzi during amastigogenesis early stages. PLoS ONE 2019, 14, e0225386. [Google Scholar] [CrossRef]
- De Pablos, L.M.; Díaz Lozano, I.M.; Jercic, M.I.; Quinzada, M.; Giménez, M.J.; Calabuig, E.; Espino, A.M.; Schijman, A.G.; Zulantay, I.; Apt, W.; et al. The C-terminal region of Trypanosoma cruzi MASPs is antigenic and secreted via exovesicles. Sci. Rep. 2016, 6, 27293. [Google Scholar] [CrossRef] [Green Version]
- Díaz Lozano, I.M.; De Pablos, L.M.; Longhi, S.A.; Zago, M.P.; Schijman, A.G.; Osuna, A. Immune complexes in chronic Chagas disease patients are formed by exovesicles from Trypanosoma cruzi carrying the conserved MASP N-terminal region. Sci. Rep. 2017, 7, 44451. [Google Scholar] [CrossRef]
- Bartholomeu, D.C.; Cerqueira, G.C.; Leão, A.C.; daRocha, W.D.; Pais, F.S.; Macedo, C.; Djikeng, A.; Teixeira, S.M.; El-Sayed, N.M. Genomic organization and expression profile of the mucin-associated surface protein (masp) family of the human pathogen Trypanosoma cruzi. Nucleic Acids Res. 2009, 37, 3407–3417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Pablos, L.M.; González, G.G.; Solano Parada, J.; Seco Hidalgo, V.; Díaz Lozano, I.M.; Gómez Samblás, M.M.; Cruz Bustos, T.; Osuna, A. Differential expression and characterization of a member of the mucin-associated surface protein family secreted by Trypanosoma cruzi. Infect. Immun. 2011, 79, 3993–4001. [Google Scholar] [CrossRef] [Green Version]
- De Pablos, L.M.; Osuna, A. Conserved regions as markers of different patterns of expression and distribution of the mucin-associated surface proteins of Trypanosoma cruzi. Infect. Immun. 2012, 80, 169–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- dos Santos, S.L.; Freitas, L.M.; Lobo, F.P.; Rodrigues-Luiz, G.F.; Mendes, T.A.; Oliveira, A.C.; Andrade, L.O.; Chiari, E.; Gazzinelli, R.T.; Teixeira, S.M.; et al. The MASP family of Trypanosoma cruzi: Changes in gene expression and antigenic profile during the acute phase of experimental infection. PLoS Negl. Trop. Dis. 2012, 6, e1779. [Google Scholar] [CrossRef] [PubMed]
- Belew, A.T.; Junqueira, C.; Rodrigues-Luiz, G.F.; Valente, B.M.; Oliveira, A.E.R.; Polidoro, R.B.; Zuccherato, L.W.; Bartholomeu, D.C.; Schenkman, S.; Gazzinelli, R.T.; et al. Comparative transcriptome profiling of virulent and non-virulent Trypanosoma cruzi underlines the role of surface proteins during infection. PLoS Pathog. 2017, 13, e1006767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serna, C.; Lara, J.A.; Rodrigues, S.P.; Marques, A.F.; Almeida, I.C.; Maldonado, R.A. A synthetic peptide from Trypanosoma cruzi mucin-like associated surface protein as candidate for a vaccine against Chagas disease. Vaccine 2014, 32, 3525–3532. [Google Scholar] [CrossRef] [Green Version]
- Durante, I.M.; La Spina, P.E.; Carmona, S.J.; Agüero, F.; Buscaglia, C.A. High-resolution profiling of linear B-cell epitopes from mucin-associated surface proteins (MASPs) of Trypanosoma cruzi during human infections. PLoS Negl. Trop. Dis. 2017, 11, e0005986. [Google Scholar] [CrossRef] [Green Version]
- Elisei, R.M.T.; Matos, C.S.; Carvalho, A.; Chaves, A.T.; Medeiros, F.A.C.; Barbosa, R.; Marcelino, A.P.; Dos Santos Emidio, K.; Coelho, E.A.F.; Duarte, M.C.; et al. Immunogenomic screening approach to identify new antigens for the serological diagnosis of chronic Chagas’ disease. Appl. Microbiol. Biotechnol. 2018, 102, 6069–6080. [Google Scholar] [CrossRef]
- López-Olmos, V.; Pérez-Nasser, N.; Piñero, D.; Ortega, E.; Hernandez, R.; Espinoza, B. Biological characterization and genetic diversity of Mexican isolates of Trypanosoma cruzi. Acta Trop. 1998, 69, 239–254. [Google Scholar] [CrossRef]
- Bosseno, M.F.; Barnabé, C.; Magallón Gastélum, E.; Lozano Kasten, F.; Ramsey, J.; Espinoza, B.; Brenière, S.F. Predominance of Trypanosoma cruzi lineage I in Mexico. J. Clin. Microbiol. 2002, 40, 627–632. [Google Scholar] [CrossRef]
- Espinoza, B.; Rico, T.; Sosa, S.; Oaxaca, E.; Vizcaino-Castillo, A.; Caballero, M.L.; Martínez, I. Mexican Trypanosoma cruzi T. cruzi I strains with different degrees of virulence induce diverse humoral and cellular immune responses in a murine experimental infection model. J. Biomed. Biotechnol. 2010, 2010, 890672. [Google Scholar] [CrossRef] [Green Version]
- Espinoza, B.; Solorzano-Domínguez, N.; Vizcaino-Castillo, A.; Martínez, I.; Elias-López, A.L.; Rodríguez-Martínez, J.A. Gastrointestinal infection with Mexican TcI Trypanosoma cruzi strains: Different degrees of colonization and diverse immune responses. Int. J. Biol. Sci. 2011, 7, 1357–1370. [Google Scholar] [CrossRef] [Green Version]
- Martínez, I.; Nogueda, B.; Martínez-Hernández, F.; Espinoza, B. Microsatellite and mini-exon analysis of Mexican human DTU I Trypanosoma cruzi strains and their susceptibility to nifurtimox and benznidazole. Vector Borne Zoonotic Dis. 2013, 13, 181–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernández-Osorio, L.A.; Márquez-Dueñas, C.; Florencio-Martínez, L.E.; Ballesteros-Rodea, G.; Martínez-Calvillo, S.; Manning-Cela, R.G. Improved method for in vitro secondary amastigogenesis of Trypanosoma cruzi: Morphometrical and molecular analysis of intermediate developmental forms. J. Biomed. Biotechnol. 2010, 2010, 283842. [Google Scholar] [CrossRef] [Green Version]
- Onami, T.M.; Lin, M.Y.; Page, D.M.; Reynolds, S.A.; Katayama, C.D.; Marth, J.D.; Irimura, T.; Varki, A.; Varki, N.; Hedrick, S.M. Generation of mice deficient for macrophage galactose- and N-acetylgalactosamine-specific lectin: Limited role in lymphoid and erythroid homeostasis and evidence for multiple lectins. Mol. Cell. Biol. 2002, 22, 5173–5181. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Velasco, M.; Ruiz-Hurtado, G.; Rueda, A.; Neco, P.; Mercado-Morales, M.; Delgado, C.; Napolitano, C.; Priori, S.G.; Richard, S.; Gómez, A.M.; et al. RyRCa2+ leak limits cardiac Ca2+ window current overcoming the tonic effect of calmodulinin mice. PLoS ONE 2011, 6, e20863. [Google Scholar] [CrossRef] [Green Version]
- Grodzki, A.C.; Berenstein, E. Antibody purification: Affinity chromatography - protein A and protein G Sepharose. Methods Mol. Biol. 2010, 588, 33–41. [Google Scholar] [CrossRef]
- Weist, S.; Eravci, M.; Broedel, O.; Fuxius, S.; Eravci, S.; Baumgartner, A. Results and reliability of protein quantification for two-dimensional gel electrophoresis strongly depend on the type of protein sample and the method employed. Proteomics 2008, 8, 3389–3396. [Google Scholar] [CrossRef]
- Sánchez, B.; Monteón, V.; Reyes, P.A.; Espinoza, B. Standardization of micro-enzyme-linked immunosorbent assay (ELISA) and Western blot for detection of Trypanosoma cruzi antibodies using extracts from Mexican strains as antigens. Arch. Med. Res. 2001, 32, 382–388. [Google Scholar] [CrossRef]
- Jimenez, V.; Henriquez, M.; Galanti, N.; Riquelme, G. Electrophysiological characterization of potassium conductive pathways in Trypanosoma cruzi. J. Cell Biochem. 2011, 112, 1093–1102. [Google Scholar] [CrossRef]
- Schägger, H.; von Jagow, G. Tricine-sodium dodecyl sulfate-polyacrylamide gel electrophoresis for the separation of proteins in the range from 1 to 100 kDa. Anal. Biochem. 1987, 166, 368–379. [Google Scholar] [CrossRef] [PubMed]
- Espinoza, B.; Tarrab-Hazdai, R.; Silman, I.; Arnon, R. Acetylcholinesterase in Schistosoma mansoni is anchored to the membrane via covalently attached phosphatidylinositol. Mol. Biochem. Parasitol. 1988, 29, 171–179. [Google Scholar] [CrossRef]
- Rodríguez-Hernández, K.D.; Martínez, I.; Agredano-Moreno, L.T.; Jiménez-García, L.F.; Reyes-Chilpa, R.; Espinoza, B. Coumarins isolated from Calophyllum brasiliense produce ultrastructural alterations and affect in vitro infectivity of Trypanosoma cruzi. Phytomedicine 2019, 61, 152827. [Google Scholar] [CrossRef] [PubMed]
- Arroyo, R.; Alderete, J.F. Two Trichomonas vaginalis surface proteinases bind to host epithelial cells and are related to levels of cytoadherence and cytotoxicity. Arch. Med. Res. 1995, 26, 279–285. [Google Scholar]
- Upham, J.P.; Pickett, D.; Irimura, T.; Anders, E.M.; Reading, P.C. Macrophage receptors for influenza A virus: Role of the macrophage galactose-type lectin and mannose receptor in viral entry. J. Virol. 2010, 84, 3730–3737. [Google Scholar] [CrossRef] [Green Version]
- Borges, A.R.; Link, F.; Engstler, M.; Jones, N.G. The Glycosylphosphatidylinositol anchor: A linchpin for cell surface versatility of trypanosomatids. Front. Cell. Dev. Biol. 2021, 9, 720536. [Google Scholar] [CrossRef]
- Buscaglia, C.A.; Campo, V.A.; Di Noia, J.M.; Torrecilhas, A.C.; De Marchi, C.R.; Ferguson, M.A.; Frasch, A.C.; Almeida, I.C. The surface coat of the mammal-dwelling infective trypomastigote stage of Trypanosoma cruzi is formed by highly diverse immunogenic mucins. J. Biol. Chem. 2004, 279, 15860–15869. [Google Scholar] [CrossRef] [Green Version]
- Giorgi, M.E.; de Lederkremer, R.M. Trans-sialidase and mucins of Trypanosoma cruzi: An important interplay for the parasite. Carbohydr. Res. 2011, 346, 1389–1393. [Google Scholar] [CrossRef]
- Piazza, R.M.; Borges, M.M.; Kloetzel, J.K.; Stolf, A.M. Reactivity of Trypanosoma cruzi strains with peanut agglutinin (PNA) correlates with number of in vitro infected host cells. Acta Trop. 1996, 61, 41–50. [Google Scholar] [CrossRef]
- Freire, T.; Robello, C.; Soulé, S.; Ferreira, F.; Osinaga, E. Sialyl-Tn antigen expression and O-linked GalNAc-Thr synthesis by Trypanosoma cruzi. Biochem. Biophys. Res. Commun. 2003, 312, 1309–1316. [Google Scholar] [CrossRef]
- de Araújo-Jorge, T.C.; de Souza, W. Interaction of Trypanosoma cruzi with macrophages: Effect of previous incubation of the parasites or the host cells with lectins. Z. Parasitenkd. 1986, 72, 153–171. [Google Scholar] [CrossRef]
- Araújo-Jorge, T.C.; De Souza, W. Interaction of Trypanosoma cruzi with macrophages. Involvement of surface galactose and N-acetyl-D-galactosamine residues on the recognition process. Acta Trop. 1988, 45, 127–136. [Google Scholar]
- Raes, G.; Brys, L.; Dahal, B.K.; Brandt, J.; Grooten, J.; Brombacher, F.; Vanham, G.; Noël, W.; Bogaert, P.; Boonefaes, T.; et al. Macrophage galactose-type C-type lectins as novel markers for alternatively activated macrophages elicited by parasitic infections and allergic airway inflammation. J. Leukoc. Biol. 2005, 77, 321–327. [Google Scholar] [CrossRef]
- Soeiro Mde, N.; Paiva, M.M.; Barbosa, H.S.; Meirelles Mde, N.; Araújo-Jorge, T.C. A cardiomyocyte mannose receptor system is involved in Trypanosoma cruzi invasion and is down-modulated after infection. Cell Struct. Funct. 1999, 24, 139–149. [Google Scholar] [CrossRef] [Green Version]
- Vázquez, A.; Ruiz-Rosado Jde, D.; Terrazas, L.I.; Juárez, I.; Gomez-Garcia, L.; Calleja, E.; Camacho, G.; Chávez, A.; Romero, M.; Rodriguez, T.; et al. Mouse macrophage galactose-type lectin (mMGL) is critical for host resistance against Trypanosoma cruzi infection. Int. J. Biol. Sci. 2014, 10, 909–920. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, T.; Pacheco-Fernández, T.; Vázquez-Mendoza, A.; Nieto-Yañez, O.; Juárez-Avelar, I.; Reyes, J.L.; Terrazas, L.I.; Rodriguez-Sosa, M. MGL1 receptor plays a key role in the control of T. cruzi infection by increasing macrophage activation through modulation of ERK1/2, c-Jun, NF-κB and NLRP3 pathways. Cells 2020, 9, 108. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Espinoza, B.; Martínez, I.; Martínez-Velasco, M.L.; Rodríguez-Sosa, M.; González-Canto, A.; Vázquez-Mendoza, A.; Terrazas, L.I. Role of a 49 kDa Trypanosoma cruzi Mucin-Associated Surface Protein (MASP49) during the Infection Process and Identification of a Mammalian Cell Surface Receptor. Pathogens 2023, 12, 105. https://doi.org/10.3390/pathogens12010105
Espinoza B, Martínez I, Martínez-Velasco ML, Rodríguez-Sosa M, González-Canto A, Vázquez-Mendoza A, Terrazas LI. Role of a 49 kDa Trypanosoma cruzi Mucin-Associated Surface Protein (MASP49) during the Infection Process and Identification of a Mammalian Cell Surface Receptor. Pathogens. 2023; 12(1):105. https://doi.org/10.3390/pathogens12010105
Chicago/Turabian StyleEspinoza, Bertha, Ignacio Martínez, María Luisa Martínez-Velasco, Miriam Rodríguez-Sosa, Augusto González-Canto, Alicia Vázquez-Mendoza, and Luis I. Terrazas. 2023. "Role of a 49 kDa Trypanosoma cruzi Mucin-Associated Surface Protein (MASP49) during the Infection Process and Identification of a Mammalian Cell Surface Receptor" Pathogens 12, no. 1: 105. https://doi.org/10.3390/pathogens12010105