
Identification and Quantification of Anti-Gp.Mur Antibodies in Human Serum Using an Insect-Cell-Based System

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Cultures and Viruses
2.2. Generation of Baculovirus Expression Construct
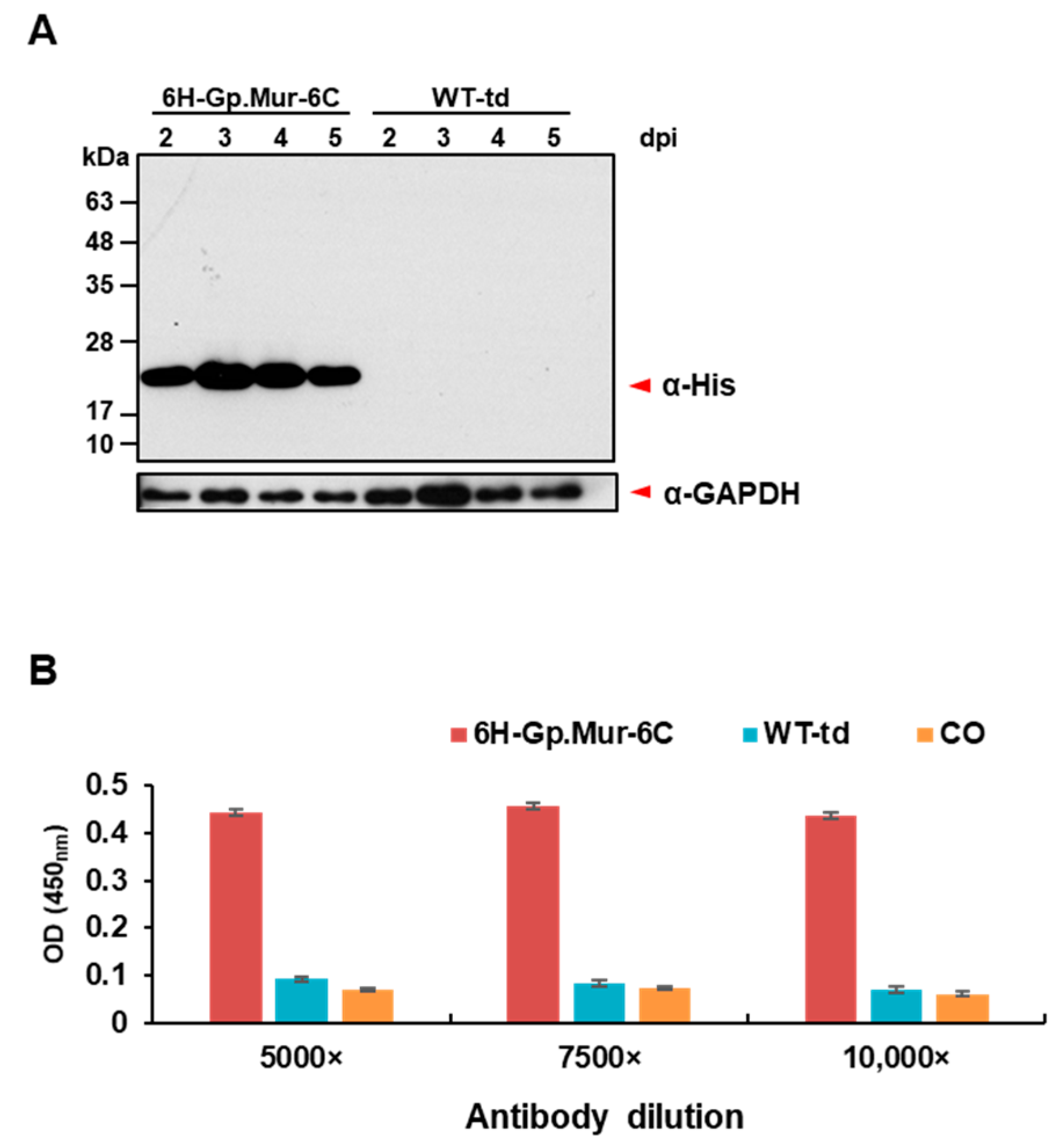
2.3. Western Blot Analysis
2.4. Cell-Based Enzyme-Linked Immunosorbent Assay (ELISA)
2.5. Human Serum Samples
2.6. Preparation of Insect Cell Suspension
2.7. Hemagglutination Inhibition Assay
2.8. Stability Assay of Cells Displaying Gp.Mur Antigen
2.9. Calculations and Statistical Analyses
3. Results
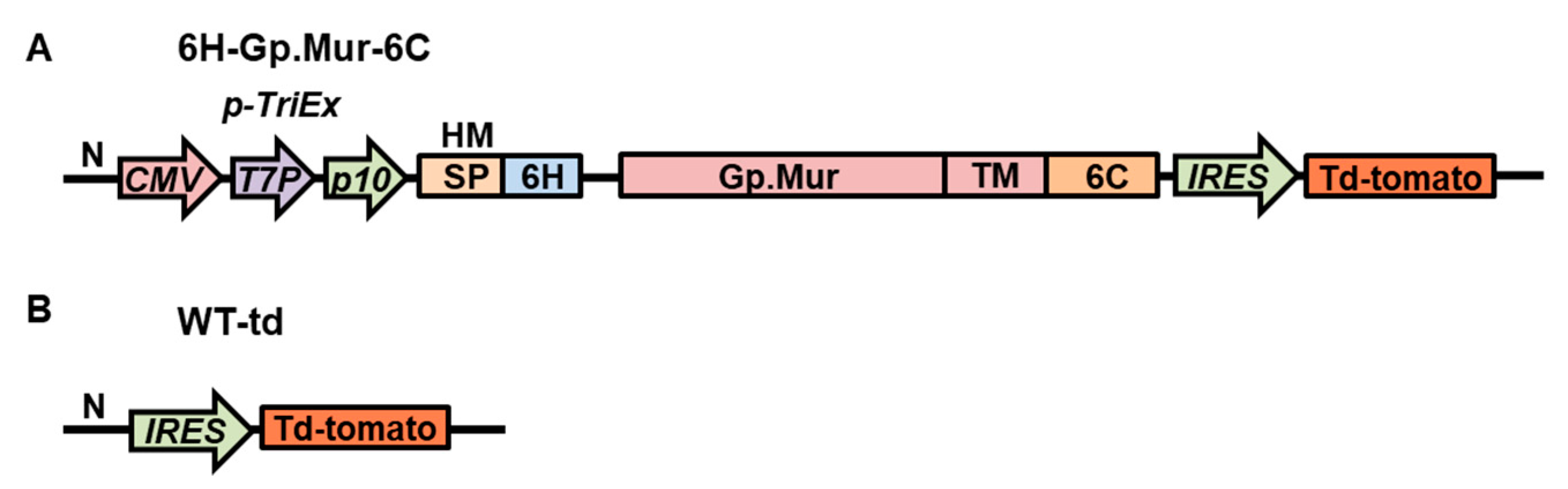
3.1. Construction of Recombinant Baculovirus That Can Display Gp.Mur Antigen on Its Cell Surface
3.2. Expression and Surface Display of Gp.Mur Antigen by Recombinant Baculovirus in Insect Cells
3.3. Hemagglutination Inhibition Assay on Human Serum Samples Harboring Anti-Gp.Mur Antibody
3.3.1. Characterization of the Serum Samples
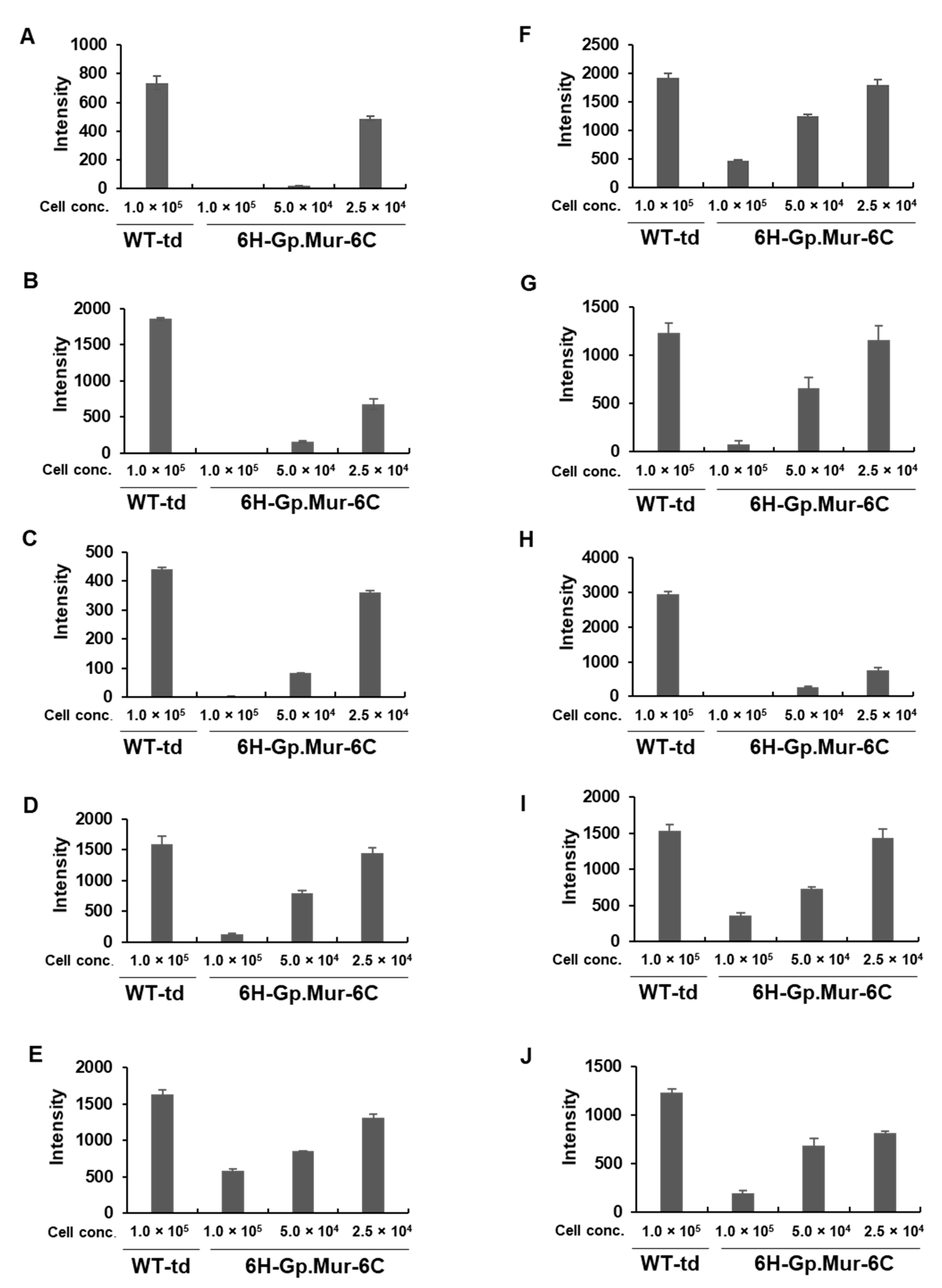
3.3.2. Adsorption Using Gp.Mur Antigen Expressed in Insect Cells
3.3.3. Grading of Hemagglutination
3.3.4. Quantitative Grading of Hemagglutination
3.3.5. Comparison of the Grading Methods
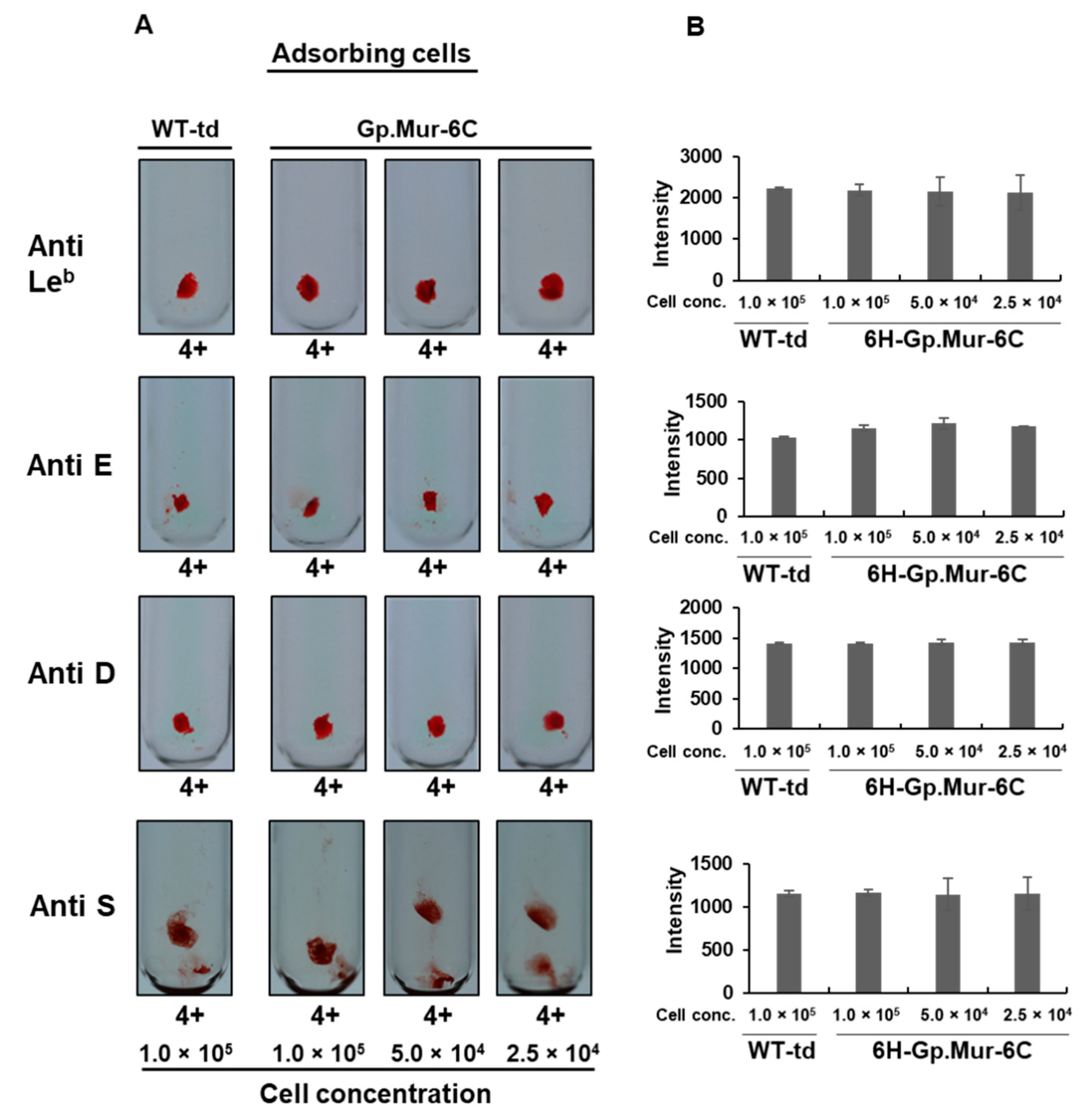
3.3.6. Determining the Specificity of Gp.Mur Antigen Expressed in Insect Cells
3.3.7. Stability of the Gp.Mur Antigen Expressed in Insect Cell
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reid, M.E. MNS blood group system: A review. Immunohematology 2009, 25, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Storry, J.R.; Castilho, L.; Chen, Q.; Daniels, G.; Denomme, G.; Flegel, W.A.; Gassner, C.; de Haas, M.; Hyland, C.; Keller, M.; et al. International society of blood transfusion working party on red cell immunogenetics and terminology: Report of the Seoul and London meetings. ISBT Sci. Ser. 2016, 11, 118–122. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.-H.; Blumenfeld, O. Molecular Genetics of Human Erythrocyte Mi111 and MiVI Glycophorins. Use of a pseudoexon in construction of two delta-alpha-delta hybrid genes resulting in antigenic diversification. J. Biol. Chem. 1991, 266, 7248–7255. [Google Scholar] [CrossRef]
- Lomas-Francis, C. Miltenberger phenotypes are glycophorin variants: A review. ISBT Sci. Ser. 2011, 6, 296–301. [Google Scholar] [CrossRef]
- Broadberry, R.E.; Lin, M. The distribution of the MiIII (Gp.Mur) phenotype among the population of Taiwan. Transfus Med. 1996, 6, 145–148. [Google Scholar] [CrossRef] [PubMed]
- Broadberry, R.E.; Lin, M. The incidence and significance of anti-“Mia” in Taiwan. Transfusion 1994, 34, 349–352. [Google Scholar] [CrossRef]
- Nadarajan, V.S.; Laing, A.A.; Saad, S.M.; Usin, M. Prevalence and specificity of red-blood-cell antibodies in a multiethnic South and East Asian patient population and influence of using novel MUT+Mur+ kodecytes on its detection. Vox Sang. 2012, 102, 65–71. [Google Scholar] [CrossRef]
- Nadarajan, V.S. The prevalence, immunogenicity, and evanescence of alloantibodies to MUT and Mur antigens of GP.Mur red blood cells in a Southeast Asian patient cohort. Transfusion 2018, 58, 1189–1198. [Google Scholar] [CrossRef]
- Lin, M.; Broadberry, R.E. Immunohematology in Taiwan. Transfus Med. Rev. 1998, 12, 56–72. [Google Scholar] [CrossRef]
- Wang, L.Y.; Liang, D.C.; Liu, H.C.; Chang, F.C.; Wang, C.L.; Chan, Y.S.; Lin, M. Alloimmunization among patients with transfusion-dependent thalassemia in Taiwan. Transfus Med. 2006, 16, 200–203. [Google Scholar] [CrossRef]
- Lamba, D.S.; Mittal, K.; Sood, T.; Bedi, R.K.; Kaur, P.; Kaur, G. Antibody screening in multitransfused patients: A prerequisite before each transfusion. Transfus Apher Sci 2014, 51, 132–133. [Google Scholar] [CrossRef]
- Hsu, K.; Kuo, M.-S.; Yao, C.-C.; Lee, T.-Y.; Chen, Y.-C.; Cheng, H.-C.; Lin, C.-H.; Yu, T.-H.; Lin, H.-J. Expedited CO2 respiration in people with Miltenberger erythrocyte phenotype GP.Mur. Sci. Rep. 2015, 5, 10327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, M. Taiwan experience suggests that RhD typing for blood transfusion is unnecessary in southeast Asian populations. Transfusion 2006, 46, 95–98. [Google Scholar] [CrossRef]
- Mallari, R.A.; Chan, A.; Powers, R.J.; Pandipati, S.; Bensing, K.M.; Biese, D.; Denomme, G.A. Fetal inheritance of Gp.Mur causing severe HDFN in an unrecognized case of maternal alloimmunization. Transfusion 2020, 60, 870–874. [Google Scholar] [CrossRef]
- Lin, C.K.; Mak, K.H.; Szeto, S.C.; Poon, K.H.; Yuen, C.M.; Chan, N.K.; Liu, H.W.; Ng, C.P. First case of haemolytic disease of the newborn due to anti-Mur in Hong Kong. Clin. Lab. Haematol 1996, 18, 19–22. [Google Scholar] [CrossRef]
- Yang, C.A.; Lin, J.A.; Chang, C.W.; Wu, K.H.; Yeh, S.P.; Ho, C.M.; Chang, J.G. Selection of GP. Mur antigen-negative RBC for blood recipients with anti-‘Mia’ records decreases transfusion reaction rates in Taiwan. Transfus Med. 2016, 26, 349–354. [Google Scholar] [CrossRef] [PubMed]
- Raman, L.; Armstrong, B.; Smart, E. Principles of laboratory techniques. ISBT Sci. Ser. 2008, 3, 33–60. [Google Scholar] [CrossRef]
- Seltsam, A.; Blasczyk, R. Recombinant blood group proteins in clinical practice—from puzzling to binary antibody testing. ISBT Sci. Ser. 2016, 11, 243–249. [Google Scholar] [CrossRef]
- Seltsam, A.; Wagner, F.; Lambert, M.; Bullock, T.; Thornton, N.; Scharberg, E.A.; Grueger, D.; Schneeweiss, C.; Blasczyk, R. Recombinant blood group proteins facilitate the detection of alloantibodies to high-prevalence antigens and reveal underlying antibodies: Results of an international study. Transfusion 2014, 54, 1823–1830. [Google Scholar] [CrossRef]
- Seltsam, A.; Grüger, D.; Blasczyk, R. Prokaryotic versus eukaryotic recombinant Lutheran blood group protein for antibody identification. Transfusion 2007, 47, 1630–1636. [Google Scholar] [CrossRef] [PubMed]
- Seltsam, A.; Grueger, D.; Blasczyk, R.; Flegel, W.A. Easy identification of antibodies to high-prevalence Scianna antigens and detection of admixed alloantibodies using soluble recombinant Scianna protein. Transfusion 2009, 49, 2090–2096. [Google Scholar] [CrossRef] [Green Version]
- Ridgwell, K.; Dixey, J.; Scott, M.L. Production of soluble recombinant proteins with Kell, Duffy and Lutheran blood group antigen activity, and their use in screening human sera for Kell, Duffy and Lutheran antibodies. Transfus Med. 2007, 17, 384–394. [Google Scholar] [CrossRef]
- Byrne, K.M.; Mercado, C.M.C.; Nnabue, T.N.; Paige, T.D.; Flegel, W.A. Inhibition of blood group antibodies by soluble substances. Immunohematology 2019, 35, 19–22. [Google Scholar] [CrossRef] [PubMed]
- Bartelsen, O.; Lansmann, S.; Nettersheim, M.; Lemm, T.; Ferlinz, K.; Sandhoff, K. Expression of recombinant human acid sphingomyelinase in insect Sf21 cells: Purification, processing and enzymatic characterization. J. Biotechnol. 1998, 63, 29–40. [Google Scholar] [CrossRef]
- Chen, W.-S.; Villaflores, O.B.; Jinn, T.-R.; Chan, M.-T.; Chang, Y.-C.; Wu, T.-Y. Expression of recombinant human interferon-γ with antiviral activity in the bi-cistronic baculovirus-insect/larval system. Biosci. Biotechnol. Biochem. 2011, 75, 1342–1348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jarvis, D.L.; Finn, E.E. Biochemical analysis of the N-glycosylation pathway in baculovirus-infected lepidopteran insect cells. Virology 1995, 212, 500–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, T.R.; Wood, H.A. Intrinsic glycosylation potentials of insect cell cultures and insect larvae. Cell Dev. Biol. Anim. 1995, 31, 659–663. [Google Scholar] [CrossRef]
- Ogonah, O.W.; Freedman, R.B.; Jenkins, N.; Patel, K.; Rooney, B.C. Isolation and Characterization of an Insect Cell Line Able to Perform Complex N-Linked Glycosylation on Recombinant Proteins. Nat. Biotechnol. 1996, 14, 197–202. [Google Scholar] [CrossRef]
- Tsai, C.-H.; Wei, S.-C.; Lo, H.-R.; Chao, Y.-C. Baculovirus as versatile vectors for protein display and biotechnological applications. Curr. Issues Mol. Biol. 2020, 34, 231–256. [Google Scholar] [CrossRef] [Green Version]
- Kost, T.A.; Condreay, J.P.; Jarvis, D.L. Baculovirus as versatile vectors for protein expression in insect and mammalian cells. Nat. Biotechnol. 2005, 23, 567–575. [Google Scholar] [CrossRef]
- Puthenveetil, R.; Lee, C.-J.; Banerjee, A. Production of Recombinant Transmembrane Proteins from Mammalian Cells for Biochemical and Structural Analyses. Curr. Protoc. Cell Biol. 2020, 87, e106. [Google Scholar] [CrossRef]
- Boivineau, J.; Haffke, M.; Jaakola, V.-P. Membrane protein expression in insect cells using the baculovirus expression vector system. Methods Mol. Biol. 2020, 2127, 63–80. [Google Scholar] [PubMed]
- Hu, Y. Baculovirus as a highly efficient expression vector in insect and mammalian cells. Acta Pharmacol. Sin. 2005, 26, 405–416. [Google Scholar] [CrossRef] [PubMed]
- De Pinheiro, C.G.M.; de Pedrosa, M.O.; Teixeira, N.C.; Ano Bom, A.P.D.; van Oers, M.M.; de Oliveira, G.G.S. Optimization of canine interleukin-12 production using a baculovirus insect cell expression system. BMC Res. Notes 2016, 9, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furuta, T.; Ogawa, T.; Katsuda, T.; Fujii, I.; Yamaji, H. Efficient production of an antibody Fab fragment using the baculovirus-insect cell system. J. Biosci. Bioeng. 2010, 110, 577–581. [Google Scholar] [CrossRef] [PubMed]
- Ikonomou, L.; Bastin, G.; Schneider, Y.J.; Agathos, S.N. Design of an efficient medium for insect cell growth and recombinant protein production. Cell Dev. Biol. Anim. 2001, 37, 549–559. [Google Scholar] [CrossRef]
- Nayyar, N.; Kaur, I.; Malhotra, P.; Bhatnagar, R.K. Quantitative proteomics of Sf21 cells during Baculovirus infection reveals progressive host proteome changes and its regulation by viral miRNA. Sci. Rep. 2017, 7, 10902. [Google Scholar] [CrossRef] [PubMed]
- Naik, N.G.; Lo, Y.-W.; Wu, T.-Y.; Lin, C.-C.; Kuo, S.-C.; Chao, Y.-C. Baculovirus as an efficient vector for gene delivery into mosquitoes. Sci. Rep. 2018, 8, 17778. [Google Scholar] [CrossRef]
- Wu, Y.; Teng, C.; Chen, Y.; Chen, S.; Chen, Y.; Lin, Y.; Wu, T. Internal ribosome entry site of Rhopalosiphum padi virus is functional in mammalian cells and has cryptic promoter activity in baculovirus-infected Sf21 cells. Acta Pharmacol. Sin. 2008, 29, 965–974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, M.; Broadberry, R.E. Modification of standard Western pretransfusion testing procedures for Taiwan. Vox Sang. 1994, 67, 199–202. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, S.; Chowdhry, M. A case report on anti-Mia antibody in a multi-transfused patient from India. Transfus. Apher. Sci. 2019, 58, 625–627. [Google Scholar] [CrossRef] [PubMed]
- Van Oers, M.M.; Pijlman, G.P.; Vlak, J.M. Thirty years of baculovirus-insect cell protein expression: From dark horse to mainstream technology. J. Gen. Virol. 2015, 96, 6–23. [Google Scholar] [CrossRef] [PubMed]
- Lo, S.C.; Chang, J.S.; Lin, S.W.S.; Lin, D.T. Immunological characterization of anti-Mia, a red blood cell alloantibody, in Taiwan. Vox Sang. 2002, 83, 162–164. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RBC Agglutination (Preadsorption) | RBC Agglutination: Conventional Tube Method | RBC Agglutination: Densitometric Method | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (Postadsorption) | (Postadsorption) | |||||||||||||||
| Adsorbing Cells | WT-td | Gp.Mur-6C | WT-td | Gp.Mur-6C | ||||||||||||
| Cell Conc. | 1.0 × 105 | 1.0 × 105 | 5.0 × 104 | 2.5 × 104 | 1.0 × 105 | 1.0 × 105 | 5.0 × 104 | 2.5 × 104 | ||||||||
| Sample | IS a | AHG b | Grade | Grade | % inh c | Grade | % inh | Grade | % inh | Int d | Int | % inh | Int | % inh | Int | % inh |
| A | 3+ | 1+ | 3+ | 0 | 100 | 1+ | 67 | 2+ | 33 | 735 | 0 | 100 | 19 | 97 | 486 | 34 |
| B | 4+ | 3+ | 4+ | 1+ | 75 | 2+ | 50 | 2+ | 50 | 1858 | 0 | 100 | 163 | 91 | 678 | 64 |
| C | 4+ | 2+ | 3+ | 0 | 100 | 0 | 100 | 1+ | 67 | 442 | 1 | 99 | 82 | 81 | 359 | 19 |
| D | 3+ | 1+ | 4+ | 1+ | 75 | 2+ | 50 | 4+ | 0 | 1584 | 124 | 92 | 796 | 50 | 1444 | 9 |
| E | 4+ | 3+ | 4+ | 1+ | 75 | 3+ | 25 | 4+ | 0 | 1628 | 583 | 64 | 844 | 48 | 1312 | 19 |
| F | 4+ | 1+ | 3+ | 1+ | 67 | 2+ | 33 | 3+ | 0 | 1919 | 460 | 76 | 1245 | 35 | 1788 | 7 |
| G | 3+ | 3+ | 4+ | 1+ | 75 | 2+ | 50 | 3+ | 25 | 1232 | 79 | 94 | 660 | 46 | 1155 | 6 |
| H | 4+ | 1+ | 4+ | 0 | 100 | 1+ | 75 | 2+ | 50 | 2953 | 0 | 100 | 264 | 91 | 747 | 75 |
| I | 4+ | 1+ | 4+ | 1+ | 75 | 2+ | 50 | 3+ | 25 | 1538 | 361 | 77 | 730 | 53 | 1434 | 7 |
| J | 2+ | 3+ | 4+ | 1+ | 75 | 2+ | 50 | 3+ | 25 | 1228 | 198 | 84 | 687 | 44 | 812 | 34 |
| Average | 82 | 55 | 28 | 89 | 64 | 27 | ||||||||||
| RBC Agglutination: Conventional Tube Method | RBC Agglutination: Densitometric Method | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Adsorbing Cells | WT-td | Gp.Mur-6C | WT-td | Gp.Mur-6C | ||||||||||
| Cell Conc. | 1.0 × 105 | 1.0 × 105 | 5.0 × 104 | 2.5 × 104 | 1.0 × 105 | 1.0 × 105 | 5.0 × 104 | 2.5 × 104 | ||||||
| Sample | Grade | Grade | % inh a | Grade | % inh | Grade | % inh | Int b | Int | % inh | OD | % inh | OD | % inh |
| Anti-Le b | 4+ | 4+ | 0 | 4+ | 0 | 4+ | 0 | 2222 | 2180 | 2 | 2147 | 3 | 2125 | 4 |
| Anti-E | 4+ | 4+ | 0 | 4+ | 0 | 4+ | 0 | 1427 | 1428 | 0 | 1395 | 2 | 1399 | 2 |
| Anti-D | 4+ | 4+ | 0 | 4+ | 0 | 4+ | 0 | 1046 | 1191 | 0 | 1144 | 0 | 1184 | 0 |
| Anti S | 4+ | 4+ | 0 | 4+ | 0 | 4+ | 0 | 1190 | 1365 | 0 | 1216 | 0 | 1181 | 0.70 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lamis, R.J.S.; Chiueh, T.-S.; Tsai, C.-H.; Lo, H.-R.; Wei, S.-C.; Chao, Y.-C. Identification and Quantification of Anti-Gp.Mur Antibodies in Human Serum Using an Insect-Cell-Based System. Diagnostics 2021, 11, 966. https://doi.org/10.3390/diagnostics11060966
Lamis RJS, Chiueh T-S, Tsai C-H, Lo H-R, Wei S-C, Chao Y-C. Identification and Quantification of Anti-Gp.Mur Antibodies in Human Serum Using an Insect-Cell-Based System. Diagnostics. 2021; 11(6):966. https://doi.org/10.3390/diagnostics11060966
Chicago/Turabian StyleLamis, Robert John S., Tsong-Shi Chiueh, Chih-Hsuan Tsai, Huei-Ru Lo, Sung-Chan Wei, and Yu-Chan Chao. 2021. "Identification and Quantification of Anti-Gp.Mur Antibodies in Human Serum Using an Insect-Cell-Based System" Diagnostics 11, no. 6: 966. https://doi.org/10.3390/diagnostics11060966