
The Deep Past of the White Shark, Carcharodon carcharias, in the Mediterranean Sea: A Synthesis of Its Palaeobiology and Palaeoecology


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction and Rationale
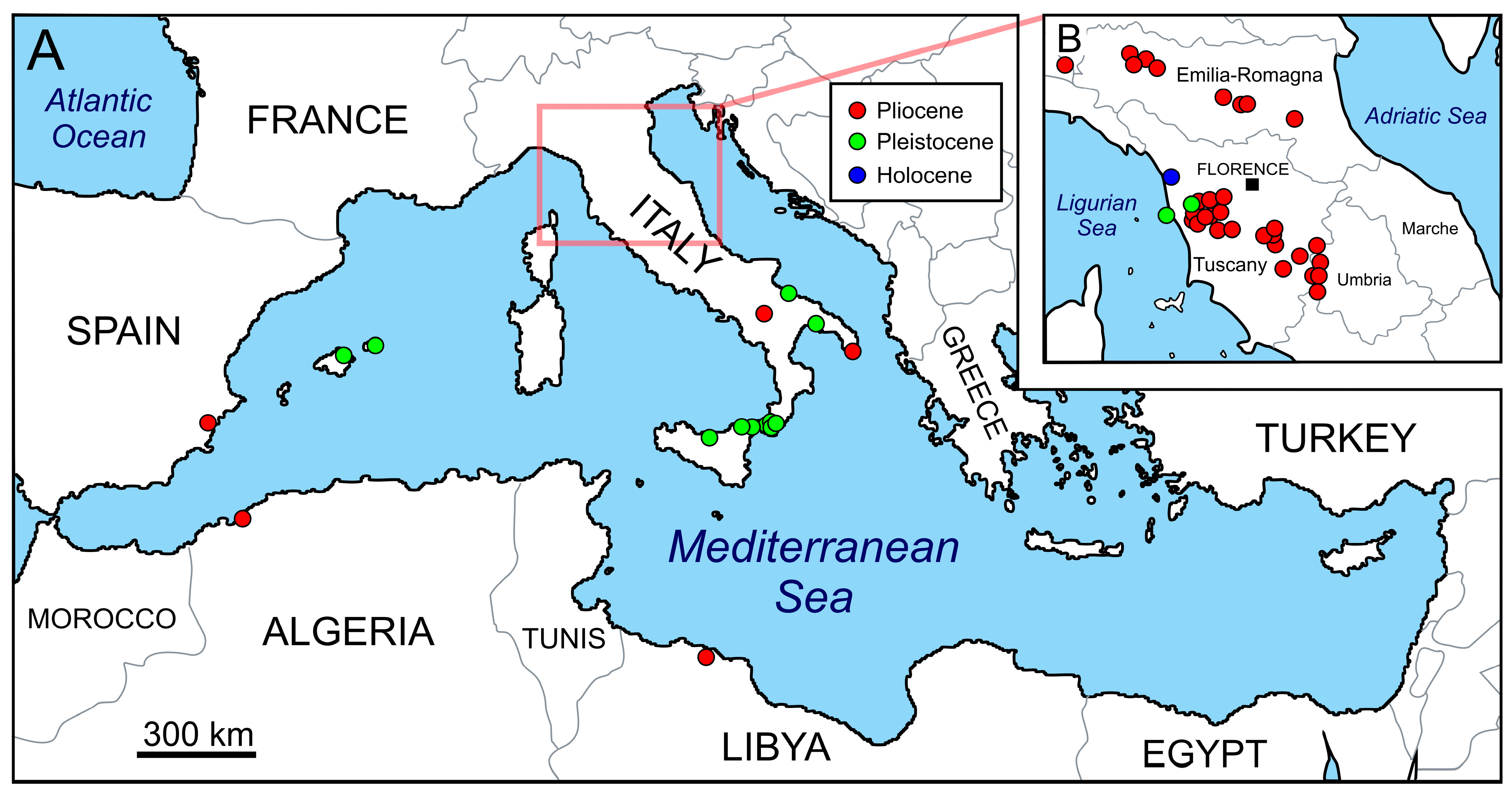
2. Origin of Carcharodon carcharias and Its Early History in the Mediterranean Sea
3. On the Maximum Body Size of the White Shark: Hints from the Mediterranean Fossils
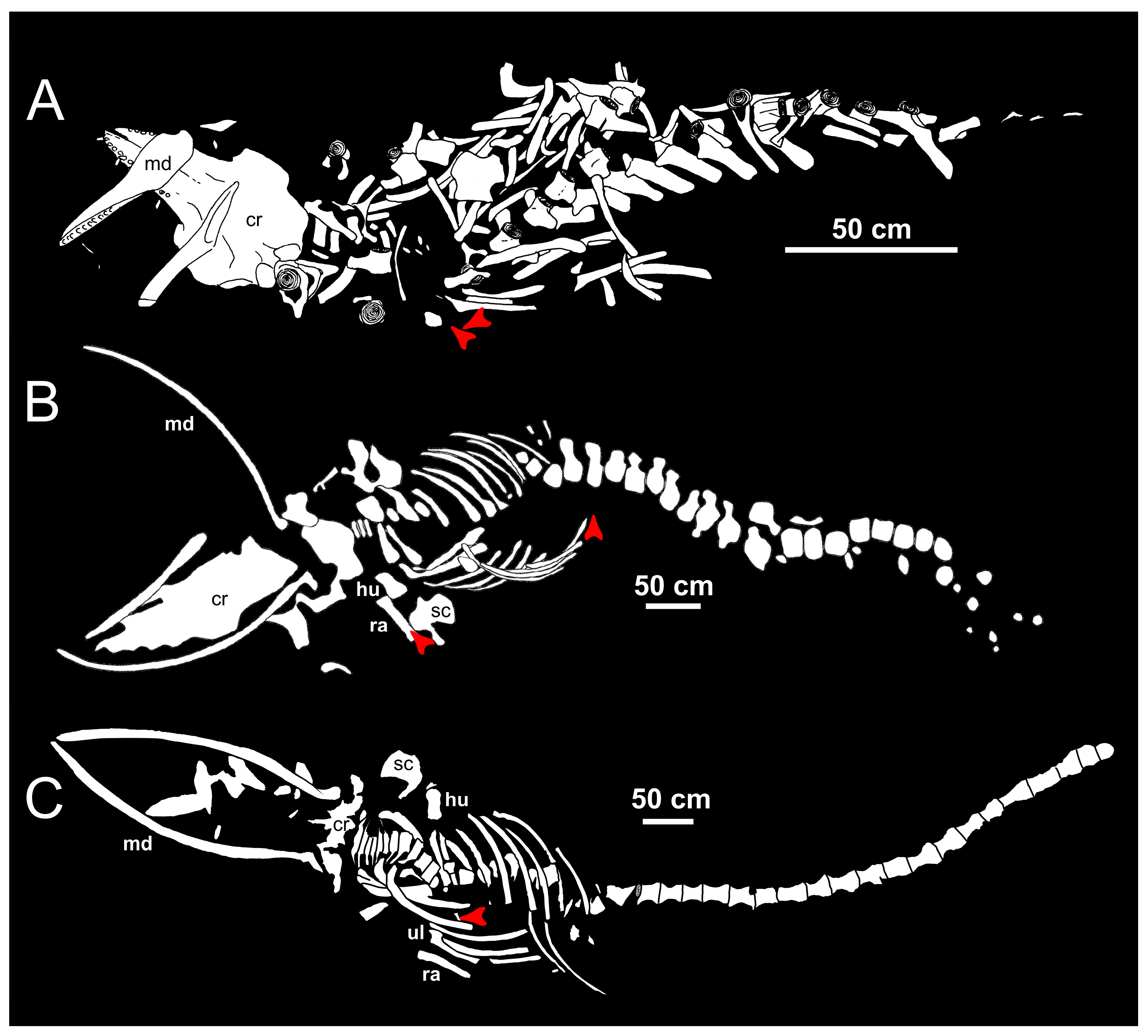
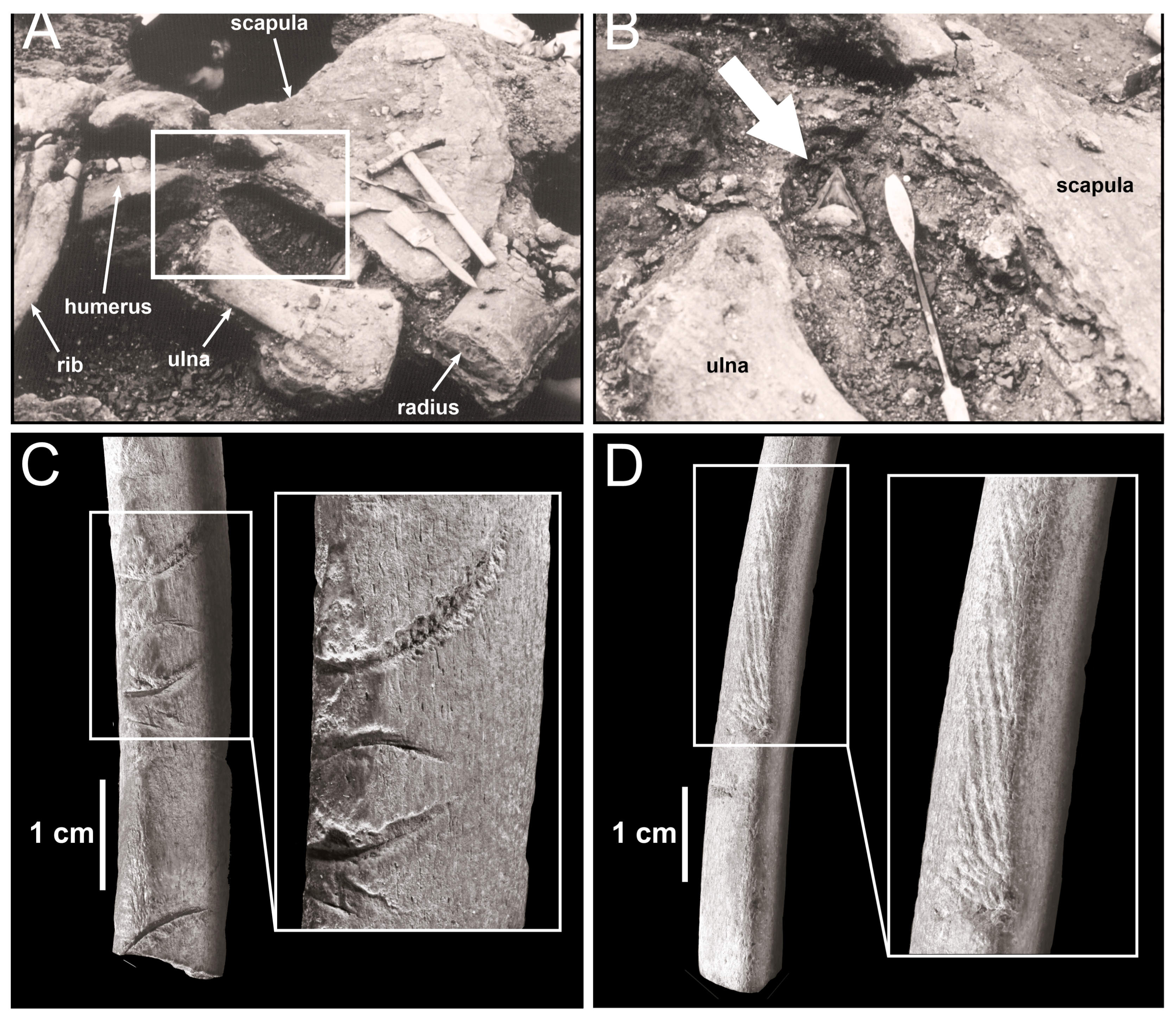
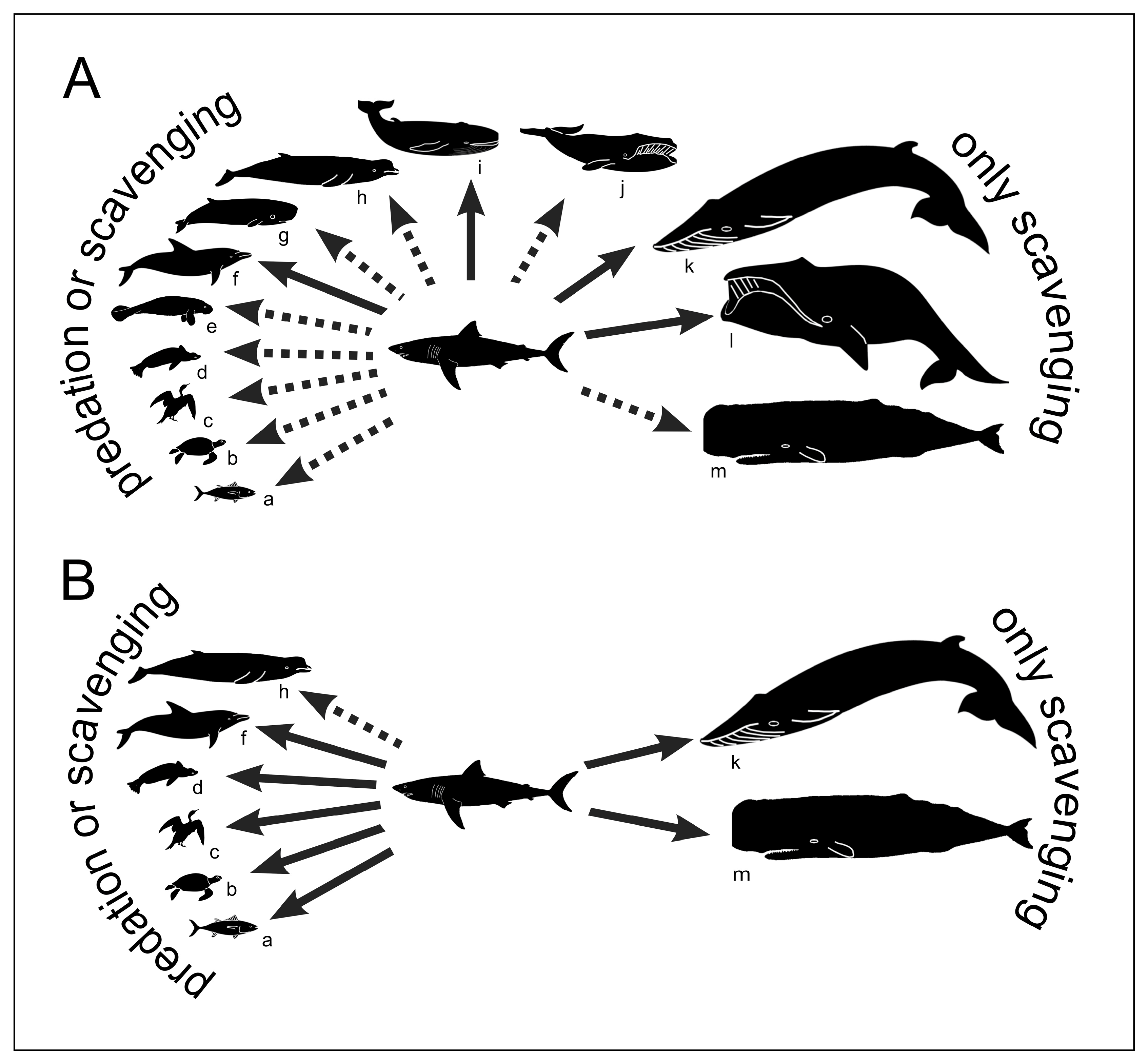
4. Trophic Ecology of the Long-Gone Mediterranean White Sharks


5. Tracing the Rise of White Sharks as the Main Mediterranean Top Predators
6. Concluding Remarks
6.1. Conclusions
6.2. Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Compagno, L.J.V. FAO Species Catalogue Vol. 4. Sharks of the World. An Annotated and Illustrated Catalogue of Shark Species Known to Date. Part 1—Hexanchiformes to Lamniformes; FAO Fisheries Synopsis No. 125, Vol. 4, Pt. 1; FAO: Rome, Italy, 1984; pp. 1–250. [Google Scholar]
- Compagno, L.J.V.; Dando, M.; Fowler, S.L. A Field Guide to the Sharks of the World; HarperCollins: London, UK, 2005; pp. 1–368. [Google Scholar]
- Serena, F. Field Identification Guide to the Sharks and Rays of the Mediterranean and Black Sea; FAO: Rome, Italy, 2005; pp. 1–97. [Google Scholar]
- Ebert, D.A.; Stehmann, M.F.W. Sharks, Batoids, and Chimaeras of the North Atlantic; FAO: Rome, Italy, 2013; pp. 1–523. [Google Scholar]
- Boldrocchi, G.; Kiszka, J.; Purkis, S.; Storai, T.; Zinzula, L.; Burkholder, D. Distribution, ecology, and status of the white shark, Carcharodon carcharias, in the Mediterranean Sea. Rev. Fish Biol. Fish. 2017, 27, 515–534. [Google Scholar] [CrossRef]
- Gubili, C.; Bilgin, R.; Kalkan, E.; Karhan, S.Ü.; Jones, C.S.; Sims, D.W.; Kabasakal, H.; Martin, A.P.; Noble, L.R. Antipodean white sharks on a Mediterranean walkabout? Historical dispersal leads to genetic discontinuity and an endangered anomalous population. P. Roy. Soc. B-Biol. Sci. 2011, 278, 1679–1686. [Google Scholar] [CrossRef]
- Moro, S.; Jona-Lasinio, G.; Block, B.; Micheli, F.; De Leo, G.; Serena, F.; Bottaro, M.; Scacco, U.; Ferretti, F. Abundance and distribution of the white shark in the Mediterranean Sea. Fish Fish. 2020, 21, 338–349. [Google Scholar] [CrossRef]
- Sperone, E.; Parise, G.; Leone, A.; Milazzo, C.; Circosta, V.; Santoro, G.; Paolillo, G.; Micarelli, P.; Tripepi, S. Spatiotemporal patterns of distribution of large predatory sharks in Calabria (central Mediterranean, southern Italy). Acta Adriat. 2012, 53, 13–24. [Google Scholar]
- Cortés, E. Standardized diet compositions and trophic levels of sharks. ICES J. Mar. Sci. 1999, 56, 707–717. [Google Scholar] [CrossRef]
- Assemat, A.; Adnet, S.; Bayez, K.; Hassler, A.; Arnaud-Godet, F.; Mollen, F.H.; Girard, C.; Martin, J.E. Exploring diet shifts and ecology in modern sharks using calcium isotopes and trace metal records of their teeth. J. Fish Biol. 2022, in press. [CrossRef]
- Bianucci, G.; Bisconti, M.; Landini, W.; Storai, T.; Zuffa, M.; Giuliani, S.; Mojetta, A. Trophic interactions between white sharks (Carcharodon carcharias) and cetaceans: A comparison between Pliocene and recent data. In Proc. 4th Europ. Elasm Assoc. Meet. Livorno; Vacchi, M., La Mesa, G., Serena, F., Sèret, B., Eds.; F. Paillart: Abbeville, France, 2002; pp. 33–48. [Google Scholar]
- Storai, T.; Vanni, S.; Zuffa, M.; Biagi, V. Presenza di Carcharodon carcharias (Linnaeus, 1758) nelle acque toscane (Mar Ligure meridionale e Mar Tirreno settentrionale; Mediterraneo): Analisi e revisione delle segnalazioni (1839–2004). Atti Soc. Toscana Sci. Nat. Mem. Ser. B 2005, 112, 153–166. [Google Scholar]
- Pujol, J.A. Un antiguo caso de predación de foca monje mediterránea adulta, Monachus monachus (Hermann, 1779) por tiburón blanco, Carcharodon carcharias (Linnaeus, 1758) en las Islas Baleares, España. Bol. R Soc. Esp. Hist. Nat. Sec. Biol. 2015, 109, 71–74. [Google Scholar]
- Serena, F.; Abella, A.J.; Bargnesi, F.; Barone, M.; Colloca, F.; Ferretti, F.; Fiorentino, F.; Jenrette, J.; Moro, S. Species diversity, taxonomy and distribution of Chondrichthyes in the Mediterranean and Black Sea. Eur. Zool. J. 2020, 87, 497–536. [Google Scholar] [CrossRef]
- Mancusi, C.; Baino, R.; Fortuna, C.; De Sola, L.G.; Morey, G.; Bradai, M.N.; Kallianotis, A.; Soldo, A.; Hemida, F.; Saad, A.A.; et al. MEDLEM database, a data collection on large Elasmobranchs in the Mediterranean and Black seas. Mediterr. Mar. Sci. 2020, 21, 276–288. [Google Scholar] [CrossRef]
- Saïdi, B.; Bradaï, M.N.; Bouaïn, A.; Guelorget, O.; Capapé, C. Capture of a pregnant female white shark, Carcharodon carcharias (Lamnidae) in the Gulf of Gabès (southern Tunisia, central Mediterranean) with comments on oophagy in sharks. Cybium 2005, 29, 303–307. [Google Scholar]
- Bradaï, M.N.; Saïdi, B. On the occurrence of the great white shark (Carcharodon carcharias) in Tunisian coasts. Rapp. Comm. Int. Mer Médit. 2013, 40, 439. [Google Scholar]
- Tiralongo, F.; Monaco, C.; De Maddalena, A. Report on a great white shark Carcharodon carcharias observed off Lampedusa, Italy. Ann. Ser. Hist. Nat. 2020, 30, 181–186. [Google Scholar] [CrossRef]
- Kabasakal, H.; Kabasakal, Ö. Recent record of the great white shark, Carcharodon carcharias (Linnaeus, 1758), from central Aegean Sea off Turkey’s coast. Ann. Ser. Hist. Nat. 2015, 25, 11–14. [Google Scholar]
- Kabasakal, H. Exploring a possible nursery ground of white shark Carcharodon carcharias in Edremit Bay (northeastern Aegean Sea, Turkey). J. Black Sea/Medit. Environ. 2020, 26, 176–189. [Google Scholar]
- Soldo, A.; Bradaï, M.N.; Walls, R.H.L. Carcharodon carcharias (Mediterranean Assessment). The IUCN Red List of Threatened Species 2016, e.T3855A16527829. Available online: https://www.iucnredlist.org/ (accessed on 23 May 2023).
- Collareta, A.; Casati, S.; Di Cencio, A.; Bianucci, G. Quaternary records of Carcharodon carcharias (Linnaeus, 1758) in Tuscany (central Italy): Implications for the palaeobiology of Mediterranean white sharks. N. Jahrb. Geol. Paläontol. Abh. 2023, 309, 65–76. [Google Scholar] [CrossRef]
- Armstrong, J. The History of the Island of Minorca; Davis: London, UK, 1752; pp. 1–260. [Google Scholar]
- Bernárdez, E.; Rábano, I. Las primeras ilustraciones de fósiles españoles: Nuevas observaciones acerca de las glosopetras de Armstrong (1752) y de Torrubia (1754). Geogaceta 2022, 71, 7–10. [Google Scholar] [CrossRef]
- Vicens, D.; Gracia, F. Carcharodon carcharias (L. 1758) en el Plistocè superior de Mallorca. Boll. Soc. Hist. Nat. Balear. 1990, 42, 167–170. [Google Scholar]
- Bisconti, M. An early Late Pliocene right whale (genus Eubalaena) from Tuscany (central Italy). Boll. Soc. Paleontol. Ital. 2002, 41, 83–92. [Google Scholar]
- Dominici, S.; Cioppi, E.; Danise, S.; Betocchi, U.; Gallai, G.; Tangocci, F.; Valleri, G.; Monechi, S. Mediterranean fossil whale falls and the adaptation of mollusks to extreme habitats. Geology 2009, 37, 815–818. [Google Scholar] [CrossRef]
- Adnet, S.; Balbino, A.C.; Antunes, M.T.; Marín-Ferrer, J.M. New fossil teeth of the White Shark (Carcharodon carcharias) from the Early Pliocene of Spain. Implication for its paleoecology in the Mediterranean. N. Jahrb. Geol. Paläontol. Abh. 2010, 256, 7–16. [Google Scholar] [CrossRef]
- Pawellek, T.; Adnet, S.; Cappetta, H.; Metais, E.; Salem, M.; Brunet, M.; Jaeger, J.J. Discovery of an earliest Pliocene relic tropical fish fauna in a newly detected cliff section (Sabratah Basin, NW Libya). N. Jahrb. Geol. Paläontol. Abh. 2012, 266, 93–114. [Google Scholar] [CrossRef]
- Danise, S.; Dominici, S. A record of fossil shallow-water whale falls from Italy. Lethaia 2014, 47, 229–243. [Google Scholar] [CrossRef]
- Freschi, A. New Carcharodon scavenging evidence on Pliocene whale bones remains from Northern Appennines. Quad. Mus. Civ. St. Nat. Ferrara 2017, 5, 33–36. [Google Scholar]
- Bendella, M.; Benyoucef, M.; Mikuláš, R.; Bouchemla, I.; Martinell, J.; Ferré, B. Shallow to marginal marine ichnoassemblages from the Upper Pliocene Slama Formation (Lower Chelif Basin, NW Algeria). Geol. Carpath. 2021, 72, 529–548. [Google Scholar] [CrossRef]
- Zazzera, A.; Girone, A.; La Perna, R.; Marino, M.; Maiorano, P.; Sardella, R.; Montenegro, V.; Francescangeli, R.; Bianucci, G. Systematics, taphonomy and palaeobiogeography of a balaenopterid (Cetacea, Mysticeti) from the Early Pleistocene of southern Italy. Geobios-Lyon 2022, 71, 51–65. [Google Scholar] [CrossRef]
- Mendiola, C. Hallazgo de Carcharodon carcharias (Linnaeus 1758) en el Plioceno superior de Conil de la Frontera (Cádiz, España). Rev. Soc. Paleontol. Elx. 2001, 7, 1–9. [Google Scholar]
- Gibbard, P.L.; Head, M.J.; Walker, M.J.C. Formal ratification of the quaternary system/period and the Pleistocene series/epoch with a base at 2.58 Ma. J. Quat. Sci. 2010, 25, 96–102. [Google Scholar] [CrossRef]
- Marsili, S. Analisi sistematica, Paleoecologica e Paleobiogeografica Della Selaciofauna Plio–Pleistocenica del Mediterraneo. Ph.D. Thesis, University of Pisa, Pisa, Italy, 2007, unpublished. [Google Scholar]
- Casier, E. Note sur la collection des poissons paléocènes et eocènes de l’enclaver de Cabinda (Congo). Ann. Mus. Roy. Congo Belge Sér. Min. Géol. Paléontol. 1960, 1, 1–48. [Google Scholar]
- Muizon, C.; de DeVries, T.J. Geology and paleontology of late Cenozoic marine deposits in the Sacaco area (Peru). Geol. Rundsch. 1985, 74, 547–563. [Google Scholar] [CrossRef]
- Nyberg, K.G.; Ciampaglio, C.N.; Wray, G.A. Tracing the ancestry of the great white shark, Carcharodon carcharias, using morphometric analyses of fossil teeth. J. Vertebr. Paleontol. 2006, 26, 806–814. [Google Scholar] [CrossRef]
- Boessenecker, R.W. A new marine vertebrate assemblage from the late Neogene Purisima formation in Central California, Part I: Fossil sharks, bony fish, birds, and implications for the age of the Purisma formation West of the San Gregorio Fault. PalArch J. Vertebr. Palaeontol. 2011, 8, 815–940. [Google Scholar]
- Cappetta, H. Handbook of Paleoichthyology, Vol. 3E. Chondrichthyes: Mesozoic and Cenozoic Elasmobranchii: Teeth; Dr. Friedrich Pfeil: Munich, Germany, 2012; pp. 1–512. [Google Scholar]
- Ehret, D.J.; Hubbell, G.; MacFadden, B.J. Exceptional preservation of the white shark Carcharodon (Lamniformes, Lamnidae) from the early Pliocene of Peru. J. Vertebr. Paleontol. 2009, 29, 1–13. [Google Scholar] [CrossRef]
- Ehret, D.J.; Macfadden, B.J.; Jones, D.S.; Devries, T.J.; Foster, D.A.; Salas-Gismondi, R. Origin of the white shark Carcharodon (Lamniformes: Lamnidae) based on recalibration of the Upper Neogene Pisco Formation of Peru. Palaeontology 2012, 55, 1139–1153. [Google Scholar] [CrossRef]
- Cione, A.L.; Cabrera, D.A.; Barla, M.J. Oldest record of the great white shark (Lamnidae, Carcharodon; Miocene) in the Southern Atlantic. Geobios-Lyon 2012, 45, 167–172. [Google Scholar] [CrossRef]
- Boessenecker, R.W.; Ehret, D.J.; Long, D.J.; Churchill, M.; Martin, E.; Boessenecker, S.J. The Early Pliocene extinction of the mega-toothed shark Otodus megalodon: A view from the eastern North Pacific. PeerJ 2019, 7, e6088. [Google Scholar] [CrossRef]
- Leone, A.; Puncher, G.N.; Ferretti, F.; Sperone, E.; Tripepi, S.; Micarelli, P.; Gambarelli, A.; Sarà, M.; Arculeo, M.; Doria, G.; et al. Pliocene colonization of the Mediterranean by Great White Shark inferred from fossil records, historical jaws, phylogeographic and divergence time analyses. J. Biogeogr. 2020, 47, 1119–1129. [Google Scholar] [CrossRef]
- O’Dea, A.; Lessios, H.A.; Coates, A.G.; Eytan, R.I.; Restrepo-Moreno, S.A.; Cione, A.L.; Collins, L.S.; de Queiroz, A.; Farris, D.W.; Norris, R.D.; et al. Formation of the Isthmus of Panama. Sci. Adv. 2016, 2, e1600883. [Google Scholar] [CrossRef]
- Pimiento, C.; Clements, C.F. When did Carcharocles megalodon become extinct? A new analysis of the fossil record. PLoS ONE 2014, 9, e111086. [Google Scholar] [CrossRef]
- Collareta, A.; Merella, M.; Mollen, F.H.; Casati, S.; Di Cencio, A. The extinct catshark Pachyscyllium distans (Probst, 1879) (Elasmobranchii: Carcharhiniformes) in the Pliocene of the Mediterranean Sea. N. Jahrb. Geol. Paläontol. Abh. 2020, 295, 129–139. [Google Scholar] [CrossRef]
- Marini, M.; Foresi, L.M.; Barbagallo, V.; Bisconti, M.; Di Stefano, A.; Muttoni, G.; Martini, I. Age and depositional environment of whale-bearing sedimentary succession from the Lower Pliocene of Tuscany (Italy): Insights from palaeomagnetism, calcareous microfossils and facies analyses. J. Mar. Sci. Eng. 2023, 11, 455. [Google Scholar] [CrossRef]
- Carnevale, G.; Gennari, R.; Lozar, F.; Natalicchio, M.; Pellegrino, L.; Dela Pierre, F. Living in a deep desiccated Mediterranean Sea: An overview of the Italian fossil record of the Messinian salinity crisis. Boll. Soc. Paleontol. Ital. 2019, 58, 109–140. [Google Scholar] [CrossRef]
- Randall, J.E. Size of the great white shark (Carcharodon). Science 1973, 181, 169–170. [Google Scholar] [CrossRef] [PubMed]
- Randall, J.E. Refutation of lenghts of 11.3, 9.0, and 6.4 attributed to the white shark, Carcharodon carcharias. Calif. Fish Game 1987, 73, 163–168. [Google Scholar]
- Castro, J.I. A summary of observations on the maximum size attained by the white shark, Carcharodon carcharias. In Global Perspectives on the Biology and Life History of the White Shark; Domeier, M.L., Ed.; CRC Press: Boca Raton, FL, USA, 2012; pp. 85–90. [Google Scholar] [CrossRef]
- De Maddalena, A.; Glaizot, O.; Oliver, G. On the great white shark, Carcharodon carcharias (Linnaeus, 1758), preserved in the Museum of Zoology in Lausanne. Mar. Life 2003, 13, 53–59. [Google Scholar]
- De Maddalena, A.; Zuffa, M.; Lipej, L.; Celona, A. An analysis of the photographic evidences of the largest great white sharks, Carcharodon carcharias (Linnaeus, 1758), captured in the Mediterranean Sea with considerations about the maximum size of the species. Ann. Ser. Hist. Nat. 2001, 11, 193–206. [Google Scholar]
- Shimada, K. The relationship between the tooth size and total body length in the white shark. J. Fossil Res. 2002, 35, 28–33. [Google Scholar]
- Shimada, K. The size of the megatooth shark, Otodus megalodon (Lamniformes: Otodontidae), revisited. Hist. Biol. 2021, 33, 904–911. [Google Scholar] [CrossRef]
- Cigala Fulgosi, F. Predation (or possible scavenging) by a great white shark on an extinct species of bottlenose dolphin in the Italian Pliocene. Tert. Res. 1990, 12, 17–336. [Google Scholar]
- De Stefano, G. Appunti sulla ittiofauna fossile dell’Emilia conservata nel Museo Geologico dell’Università di Parma. Boll. Soc. Geol. Ital. 1912, 31, 35–78. [Google Scholar]
- Manganelli, G.; Spadini, V. Gli Squali del Pliocene Senese; Cantagalli: Siena, Italy, 2003; pp. 1–80. [Google Scholar]
- Cappo, M. Size and age of the white pointer shark Carcharodon carcharias (Linnaeus): Was Peter Riseley’s white pointer a world record? Safish 1998, 13, 11–13. [Google Scholar]
- Freschi, A.; Cau, S. Nuovi segni d’interazione trofica tra Carcharodon carcharias ed un odontoceto dal bacino plio- pleistocenico di Castell’Arquato (Piacenza, Appennino Settentrionale). Parva Nat. 2021, 15, 133–156. [Google Scholar]
- Long, D.J.; Jones, R.E. White shark predation and scavenging on cetaceans in the eastern North Pacific Ocean. In Great White Sharks: The Biology of Carcharodon carcharias; Klimley, A.P., Ainley, D.G., Eds.; Academic Press: San Diego, CA, USA, 1996; pp. 293–307. [Google Scholar] [CrossRef]
- Estrada, J.A.; Rice, A.N.; Natanson, L.J.; Skomal, G.B. Use of isotopic analysis of vertebrae in reconstructing ontogenetic feeding ecology in white sharks. Ecology 2006, 87, 829–834. [Google Scholar] [CrossRef] [PubMed]
- Martin, J.E.; Tacail, T.; Adnet, S.; Girard, C.; Balter, V. Calcium isotopes reveal the trophic position of extant and fossil elasmobranchs. Chem. Geol. 2015, 415, 118–125. [Google Scholar] [CrossRef]
- Kast, E.R.; Griffiths, M.L.; Kim, S.L.; Rao, Z.C.; Shimada, K.; Becker, M.A.; Maisch, H.M.; Eagle, R.A.; Clarke, C.A.; Neumann, A.N.; et al. Cenozoic megatooth sharks occupied extremely high trophic positions. Sci. Adv. 2022, 8, eabl6529. [Google Scholar] [CrossRef]
- McCormack, J.; Griffiths, M.L.; Kim, S.L.; Shimada, K.; Karnes, M.; Maisch, H.; Pederzani, S.; Bourgon, N.; Jaouen, K.; Becker, M.A.; et al. Trophic position of Otodus megalodon and great white sharks through time revealed by zinc isotopes. Nat. Commun. 2022, 13, 2980. [Google Scholar] [CrossRef]
- Marx, F.G.; Fordyce, R.E. Baleen boom and bust: A synthesis of mysticete phylogeny, diversity and disparity. Roy. Soc. Open Sci. 2015, 2, 140434. [Google Scholar] [CrossRef]
- Bianucci, G.; Sorbi, S.; Vaiani, S.C.; Landini, W. Pliocene marine mammals from Italy: A systematic and stratigraphic overview. In International Conference on Vertebrate Palaeobiogeography and Continental Bridges across Tethys, Mesogea, and Mediterranean Sea; Abstracts Book; Fanti, F., Spalletta, C., Eds.; University of Bologna: Bologna, Italy, 2009; pp. 9–12. [Google Scholar]
- Bianucci, G.; Pesci, F.; Collareta, A.; Tinelli, C. A new Monodontidae (Cetacea, Delphinoidea) from the lower Pliocene of Italy supports a warm-water origin for narwhals and white whales. J. Vertebr. Paleontol. 2019, 39, e1645148. [Google Scholar] [CrossRef]
- Sorbi, S.; Domning, D.P.; Vaiani, S.C.; Bianucci, G. Metaxytherium subapenninum (Bruno, 1839) (Mammalia, Dugongidae), the latest sirenian of the Mediterranean Basin. J. Vertebr. Paleontol. 2012, 32, 686–707. [Google Scholar] [CrossRef]
- Barbolani di Montauto, G. L’Histiophorus herschelii (Gray) nel Terziario superiore. Palaeontogr. It. 1910, 16, 1–22. [Google Scholar]
- Taskavak, E.; Akcinar, S.C. Marine records of the Nile soft-shelled turtle, Trionyx triunguis from Turkey. Mar. Biodivers. Rec. 2009, 2, e9. [Google Scholar] [CrossRef]
- Georgalis, G.L.; Joyce, W.G. A review of the fossil record of Old World turtles of the clade Pan-Trionychidae. Bull. Peabody Mus. Nat. Hist. 2017, 58, 115–208. [Google Scholar] [CrossRef]
- Arnold, P.W. Predation on harbour porpoise, Phocoena phocoena, by a white shark, Carcharodon carcharias. J. Fish Res. Board Can. 1972, 29, 1213–1214. [Google Scholar] [CrossRef]
- Fergusson, I.K.; Compagno, L.J.V.; Marks, M.A. Predation by white sharks Carcharodon carcharias (Chondrichthyes: Lamnidae) upon chelonians, with new records from the Mediterranean Sea and a first record of the ocean sunfish Mola mola (Osteichthyes: Molidae) as stomach contents. Environ. Biol. Fishes 2000, 58, 447–453. [Google Scholar] [CrossRef]
- Fallows, C.; Gallagher, A.J.; Hammerschlag, N. White sharks (Carcharodon carcharias) scavenging on whales and its potential role in further shaping the ecology of an apex predator. PLoS ONE 2013, 8, e60797. [Google Scholar] [CrossRef]
- Swartz, S.L. Gray Whale: Eschrichtius robustus. In Encyclopedia of Marine Mammals, 3rd, ed.; Würsig, B., Thewissen, J.G.M., Kovacs, K.M., Eds.; Academic Press: London, UK, 2018; pp. 422–428. [Google Scholar]
- Dines, S.; Gennari, E. First observations of white sharks (Carcharodon carcharias) attacking a live humpback whale (Megaptera novaeangliae). Mar. Freshw. Res. 2020, 71, 1205–1210. [Google Scholar] [CrossRef]
- Collareta, A.; Casati, S.; Di Cencio, A. The palaeobiology of the false mako shark, Parotodus benedenii (Le Hon, 1871): A view from the Pliocene Mediterranean Sea. J. Mar. Sci. Eng. 2023, 11, 1990. [Google Scholar] [CrossRef]
- Collareta, A.; Merella, M.; Casati, S.; Di Cencio, A.; Bianucci, G. Smoking guns for cold cases: The find of a Carcharhinus tooth piercing a fossil cetacean rib, with notes on the feeding ecology of some Mediterranean Pliocene requiem sharks. N. Jahrb. Geol. Paläontol. Abh. 2022, 305, 145–152. [Google Scholar] [CrossRef]
- Bisconti, M. Evolutionary history of Balaenidae. Cranium 2003, 20, 9–50. [Google Scholar]
- Bianucci, G.; Landini, W.; Buckeridge, J.S. Whale barnacles and Neogene cetacean migration routes. N. Z. J. Geol. Geop. 2006, 49, 115–120. [Google Scholar] [CrossRef]
- Collareta, A.; Margiotta, S.; Varola, A.; Catanzariti, R.; Bosselaers, M.; Bianucci, G. A new whale barnacle from the early Pleistocene of Italy suggests an ancient right whale breeding ground in the Mediterranean. C.R. Palevol. 2016, 15, 473–481. [Google Scholar] [CrossRef]
- Collareta, A.; Regattieri, E.; Zanchetta, G.; Lambert, O.; Catanzariti, R.; Bosselaers, M.; Covelo, P.; Varola, A.; Bianucci, G. New insights on ancient cetacean movement patterns from oxygen-isotope analyses of a Mediterranean Pleistocene whale barnacle. N. Jahrb. Geol. Paläontol. Abh. 2018, 288, 143–159. [Google Scholar] [CrossRef]
- Taylor, J.K.D.; Mandelman, J.W.; McLellan, W.A.; Moore, M.J.; Skomal, G.B.; Rotstein, D.S.; Kraus, S.D. Shark predation on North Atlantic right whales (Eubalaena glacialis) in the southeastern United States calving ground. Mar. Mammal Sci. 2013, 29, 204–212. [Google Scholar] [CrossRef]
- Shimada, K. Skeletal anatomy of the Late Cretaceous lamniform shark, Cretoxyrhina mantelli, from the Niobrara Chalk in Kansas. J. Vert. Paleontol. 1997, 17, 642–652. [Google Scholar] [CrossRef]
- Kriwet, J.; Witzmann, F.; Klug, S.; Heidtke, U.H.J. First direct evidence of a vertebrate three-level trophic chain in the fossil record. Proc. R. Soc. B Biol. Sci. 2008, 275, 181–186. [Google Scholar] [CrossRef]
- Fanti, F.; Minelli, D.; Conte, G.L.; Miyashita, T. An exceptionally preserved Eocene shark and the rise of modern predator–prey interactions in the coral reef food web. Zool. Lett. 2016, 2, 1–18. [Google Scholar] [CrossRef]
- Amalfitano, J.; Dalla Vecchia, F.M.; Giusberti, L.; Fornaciari, E.; Luciani, V.; Roghi, G. Direct evidence of trophic interaction between a large lamniform shark, Cretodus sp., and a marine turtle from the Cretaceous of northeastern Italy. Palaeogeogr. Palaeoclim. 2017, 469, 104–121. [Google Scholar] [CrossRef]
- Collareta, A.; Landini, W.; Chacaltana-Budiel, C.A.; Valdivia-Vera, W.; Altamirano-Sierra, A.; Urbina, M.; Bianucci, G. A well preserved skeleton of the fossil shark Cosmopolitodus hastalis from the late Miocene of Peru, featuring fish remains as fossilized stomach contents. Riv. Ital. Paleontol. Stratigr. 2017, 123, 11–22. [Google Scholar] [CrossRef]
- Citron, S.; Geisler, J.H.; Collareta, A.; Bianucci, G. Systematics, phylogeny and feeding behavior of the oldest killer whale: A reappraisal of Orcinus citoniensis from the Pliocene of Tuscany (Italy). Boll. Della Soc. Paleontol. Ital. 2022, 61, 167–186. [Google Scholar] [CrossRef]
- Capellini, G. Balenottere fossili del Bolognese. Mem. Accad. Sci. Istit. Bologna Ser. 2 1865, 4, 3–24. [Google Scholar]
- Bianucci, G.; Sorce, B.; Storai, T.; Landini, W. Killing in the Pliocene: Shark attack on a dolphin from Italy. Palaeontology 2010, 53, 457–470. [Google Scholar] [CrossRef]
- Collareta, A.; Cigala Fulgosi, F.; Bianucci, G. A new kogiid sperm whale from northern Italy supports psychrospheric conditions in the early Pliocene Mediterranean Sea. Acta Palaeontol. Pol. 2019, 64, 609–626. [Google Scholar] [CrossRef]
- Muñiz, F.; Belaústegui, Z.; Toscano, A.; Ramirez-Cruzado, S.; Gámez Vintaned, J.A. New ichnospecies of Linichnus Jacobsen & Bromley, 2009. Ichnos 2020, 27, 344–351. [Google Scholar] [CrossRef]
- Bisconti, M.; Damarco, P.; Pavia, M.; Sorce, B.; Carnevale, G. Marzanoptera tersillae, a new balaenopterid genus and species from the Pliocene of Piedmont, north-west Italy. Zool. J. Linn. Soc.-Lond. 2021, 192, 1253–1292. [Google Scholar] [CrossRef]
- Kent, B.W. Speculations on the size and morphology of the extinct lamnoid shark, Parotodus benedeni (Le Hon). Mosasaur 1999, 6, 11–15. [Google Scholar]
- Kent, B.W.; Powell, G.W., Jr. Reconstructed dentition of the rare lamnoid shark, Parotodus benedeni (Le Hon) from the Yorktown Formation (Early Pliocene) at Lee Creek Mine, North Carolina. Mosasaur 1999, 6, 1–10. [Google Scholar]
- Marsili, S. Systematic, paleoecologic and paleobiogeographic analysis of the Plio-Pleistocene Mediterranean elasmobranch fauna. Atti Soc. Toscana Sci. Nat. Mem. Ser. A 2008, 113, 81–88. [Google Scholar]
- Portis, A. Catalogo descrittivo dei Talassoterii rinvenuti nei terreni Terziarii del Piemonte e della Liguria. Mem. R. Accad. Sci. Torino 1885, 37, 247–365. [Google Scholar] [CrossRef]
- Scott, B.; Anderson, P. Identifying competitive exclusion in the vertebrate fossil record: Lessons from early vertebrate morphometrics. Vertebr. Anat. Morph. Palaeontol. 2021, 9, 33–34. [Google Scholar]
- Collareta, A.; Insacco, G.; Reitano, A.; Catanzariti, R.; Bosselaers, M.; Montes, M.; Bianucci, G. Fossil whale barnacles from the lower Pleistocene of Sicily shed light on the coeval Mediterranean cetacean fauna. Carnets Geol. 2018, 18, 9–22. [Google Scholar] [CrossRef]
- Pimiento, C.; Griffin, J.N.; Clements, C.F.; Silvestro, D.; Varela, S.; Uhen, M.D.; Jaramillo, C. The Pliocene marine megafauna extinction and its impact on functional diversity. Nat. Ecol. Evol. 2017, 1, 1100–1106. [Google Scholar] [CrossRef] [PubMed]
- Prista, G.; Estevens, M.; Agostinho, R.; Cachão, M. The disappearance of the European/North African Sirenia (Mammalia). Palaeogeogr. Palaeoclim. 2013, 387, 1–5. [Google Scholar] [CrossRef]
- Rodrigues, A.S.L.; Charpentier, A.; Bernal-Casasola, D.; Gardeisen, A.; Nores, C.; Pis Millán, J.A.; McGrath, K.; Speller, C.F. Forgotten Mediterranean calving grounds of grey and North Atlantic right whales: Evidence from Roman archaeological records. P. Roy. Soc. B-Biol. Sci. 2018, 285, 20180961. [Google Scholar] [CrossRef] [PubMed]
- Rey-Iglesia, A.; Gaubert, P.; Espregueira Themudo, G.; Pires, R.; De La Fuente, C.; Freitas, L.; Aguilar, A.; Borrell, A.; Krakhmalnaya, T.; Vasconcelos, R.; et al. Mitogenomics of the endangered Mediterranean monk seal (Monachus monachus) reveals dramatic loss of diversity and supports historical gene-flow between Atlantic and eastern Mediterranean populations. Zool. J. Linn. Soc.-Lond. 2021, 191, 1147–1159. [Google Scholar] [CrossRef]
- Pethybridge, H.R.; Parrish, C.C.; Bruce, B.D.; Young, J.W.; Nichols, P.D. Lipid, fatty acid and energy density profiles of white sharks: Insights into the feeding ecology and ecophysiology of a complex top predator. PLoS ONE 2014, 9, e97877. [Google Scholar] [CrossRef]
- Goldman, K.J. Regulation of body temperature in the white shark, Carcharodon carcharias. J. Comp. Physiol. B 1997, 167, 423–429. [Google Scholar] [CrossRef]
- Dickson, K.A.; Graham, J.B. Evolution and consequences of endothermy in fishes. Physiol. Biochem. Zool. 2004, 77, 998–1018. [Google Scholar] [CrossRef]
- Watanabe, Y.Y.; Goldman, K.J.; Caselle, J.E.; Chapman, D.D.; Papastamatiou, Y.P. Comparative analyses of animal-tracking data reveal ecological significance of endothermy in fishes. Proc. Natl. Acad. Sci. USA 2015, 112, 6104–6109. [Google Scholar] [CrossRef]
- Griffiths, L.M.; Eagle, R.A.; Kim, S.L.; Flores, R.; Becker, M.A.; Maisch, H.M., IV; Trayler, R.B.; Chan, R.L.; McCormack, J.; Akhtara, A.A.; et al. Endothermic physiology of extinct megatooth sharks. Proc. Natl. Acad. Sci. USA 2023, 120, e221815312. [Google Scholar] [CrossRef]
- Dolton, H.R.; Jackson, A.L.; Deaville, R.; Hall, J.; Hall, G.; McManus, G.; Perkins, M.W.; Rolfe, R.A.; Snelling, E.P.; Houghton, J.D.R.; et al. Regionally endothermic traits in planktivorous basking sharks Cetorhinus maximus. Endanger. Species Res. 2023, 51, 227–232. [Google Scholar] [CrossRef]
- Landini, W. Revisione degli «ittiodontoliti pliocenici» della Collezione Lawley. Palaeontogr. Ital. 1977, 70, 92–134. [Google Scholar]
- Cigala Fulgosi, F.; Casati, S.; Orlandini, A.; Persico, D. A small fossil fish fauna, rich in Chlamydoselachus teeth, from the Late Pliocene of Tuscany (Siena, central Italy). Cainozoic Res. 2009, 6, 3–23. [Google Scholar]
- Bianucci, G.; Geisler, J.H.; Citron, S.; Collareta, A. The origins of the killer whale ecomorph. Curr. Biol. 2022, 32, 1843–1851. [Google Scholar] [CrossRef] [PubMed]
- Villafaña, J.A.; Hernandez, S.; Alvarado, A.; Shimada, K.; Pimiento, C.; Rivadeneira, M.M.; Kriwet, J. First evidence of a palaeo-nursery area of the great white shark. Sci. Rep. 2020, 10, 8502. [Google Scholar] [CrossRef] [PubMed]
- Ghelardoni, R. Resti fossili di pesci nel livello a “Purpura” delle sabbie di Torre del Lago. Atti Soc. Toscana Sci. Nat. 1956, 63, 33–39. [Google Scholar]
- Fergusson, I.K. Distribution and autecology of the white shark in the Eastern North Atlantic Ocean and the Mediterranean Sea. In Great White Sharks: The Biology of Carcharodon Carcharias; Klimley, A.P., Ainley, D.G., Eds.; Academic Press: San Diego, CA, USA, 1996; pp. 321–345. [Google Scholar] [CrossRef]
- Dietl, G.P.; Kidwell, S.M.; Brenner, M.; Burney, D.A.; Flessa, K.W.; Jackson, S.T.; Koch, P.L. Conservation paleobiology: Leveraging knowledge of the past to inform conservation and restoration. Annu. Rev. Earth Planet. Sci. 2015, 43, 79–103. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Collareta, A.; Casati, S.; Di Cencio, A.; Bianucci, G. The Deep Past of the White Shark, Carcharodon carcharias, in the Mediterranean Sea: A Synthesis of Its Palaeobiology and Palaeoecology. Life 2023, 13, 2085. https://doi.org/10.3390/life13102085
Collareta A, Casati S, Di Cencio A, Bianucci G. The Deep Past of the White Shark, Carcharodon carcharias, in the Mediterranean Sea: A Synthesis of Its Palaeobiology and Palaeoecology. Life. 2023; 13(10):2085. https://doi.org/10.3390/life13102085
Chicago/Turabian StyleCollareta, Alberto, Simone Casati, Andrea Di Cencio, and Giovanni Bianucci. 2023. "The Deep Past of the White Shark, Carcharodon carcharias, in the Mediterranean Sea: A Synthesis of Its Palaeobiology and Palaeoecology" Life 13, no. 10: 2085. https://doi.org/10.3390/life13102085