
Proline and Proline Analogues Improve Development of Mouse Preimplantation Embryos by Protecting Them against Oxidative Stress

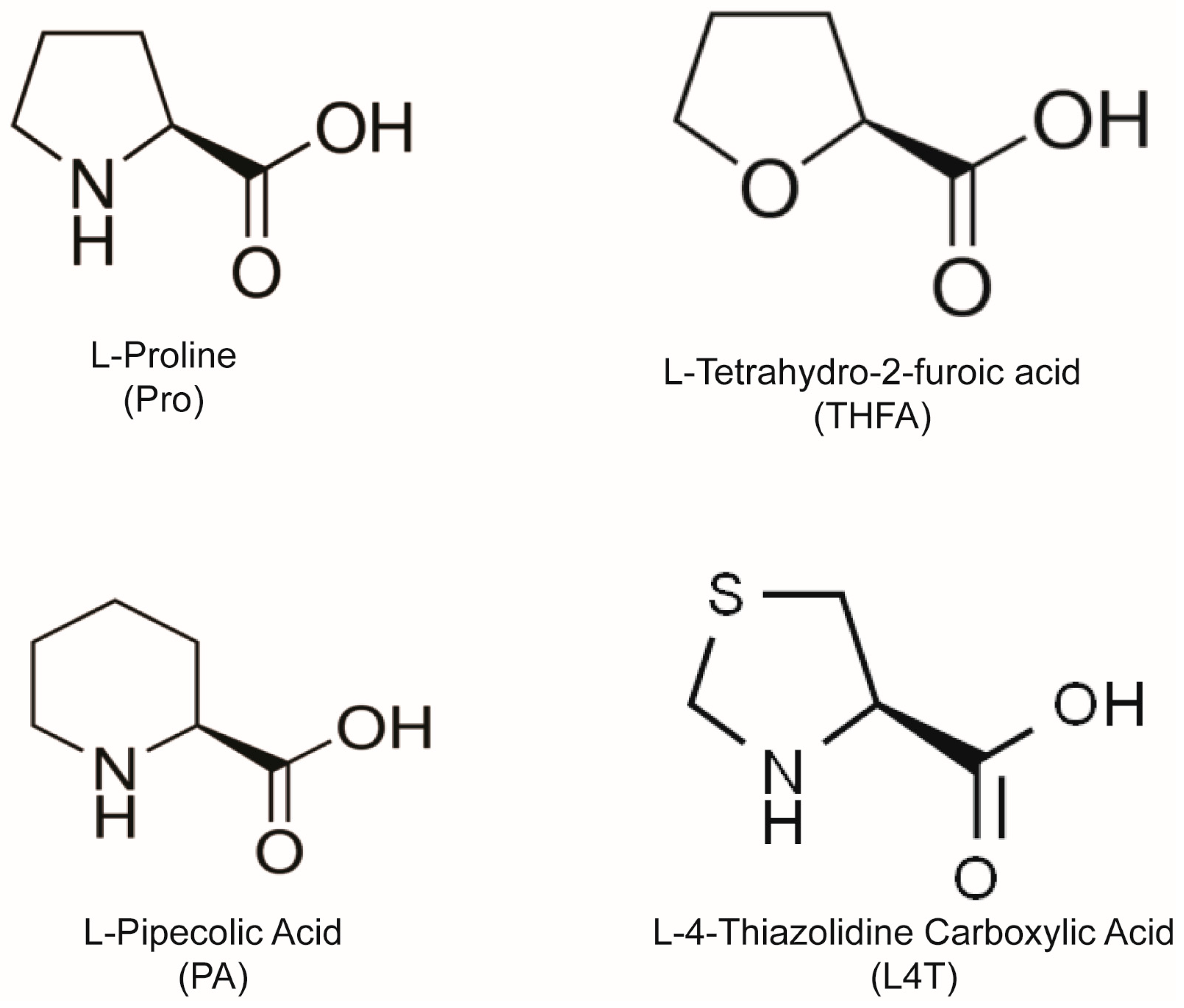{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals (Mus Musculus)
2.2. Zygote Collection and Media Preparation
2.3. Embryo Culture and Scoring Developmental Progression
2.4. Imaging of Reactive Oxygen Species and Mitochondrial Activity
2.5. Analysis of Confocal Images
2.6. Measurement of L-[3H]-Pro Uptake in 2- and 4-Cell Embryos
3. Results
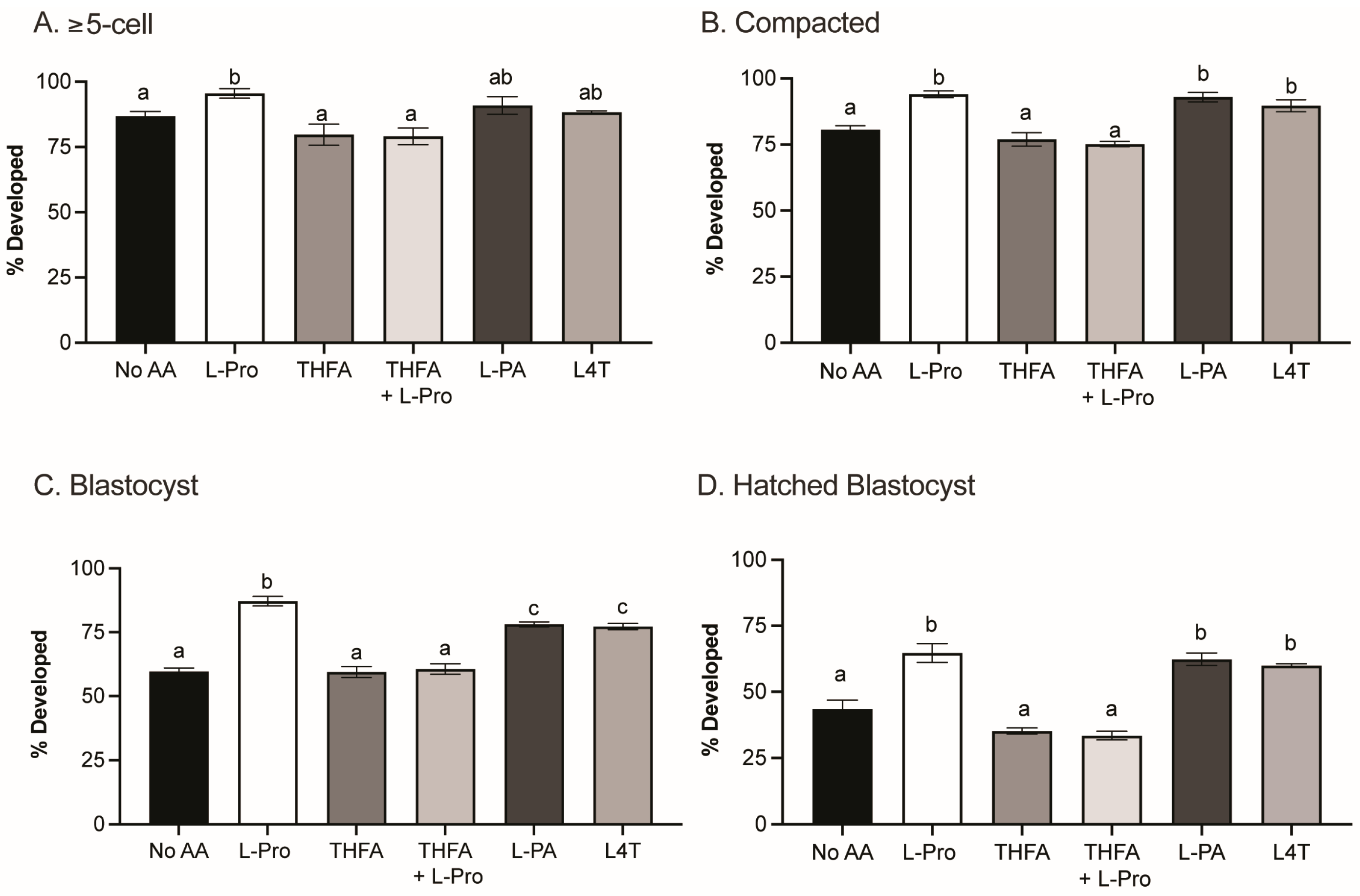
3.1. Pro, PA and L4T Improve Embryo Development to the Blastocyst Stage
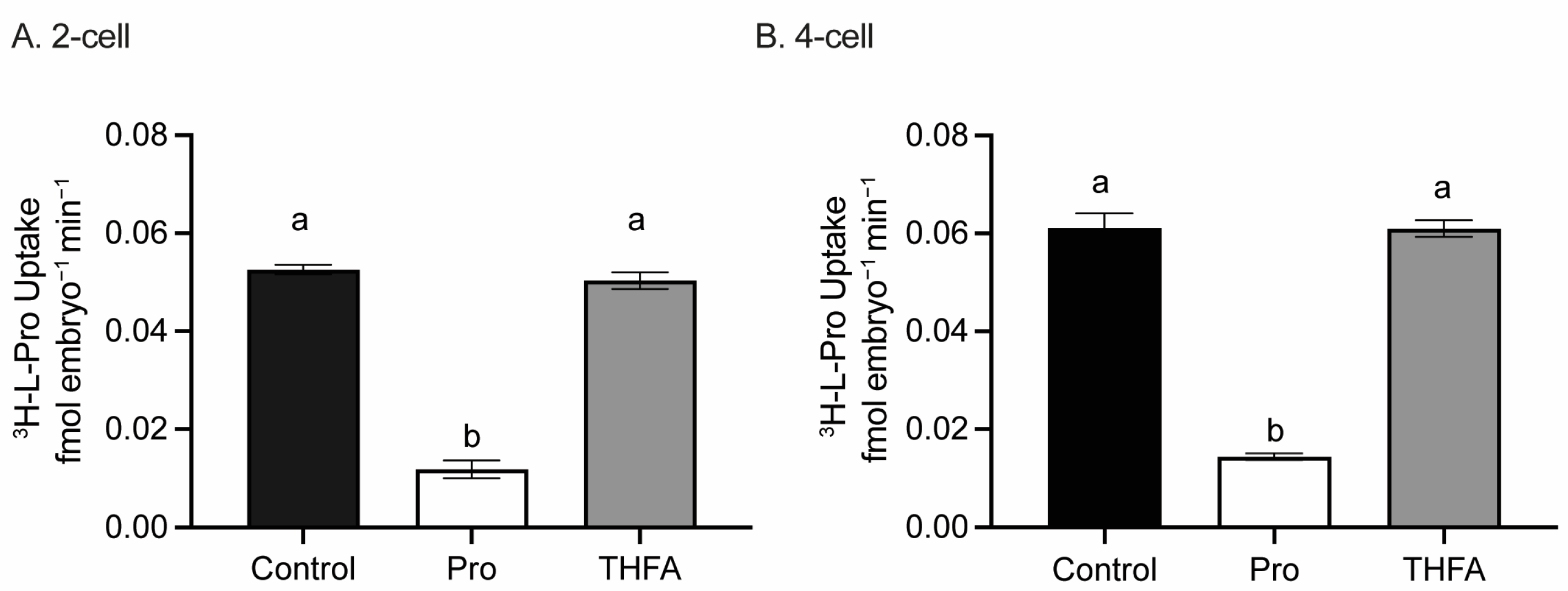
3.2. THFA Does Not Prevent the Uptake of Pro in 2- or 4-Cell Embryos
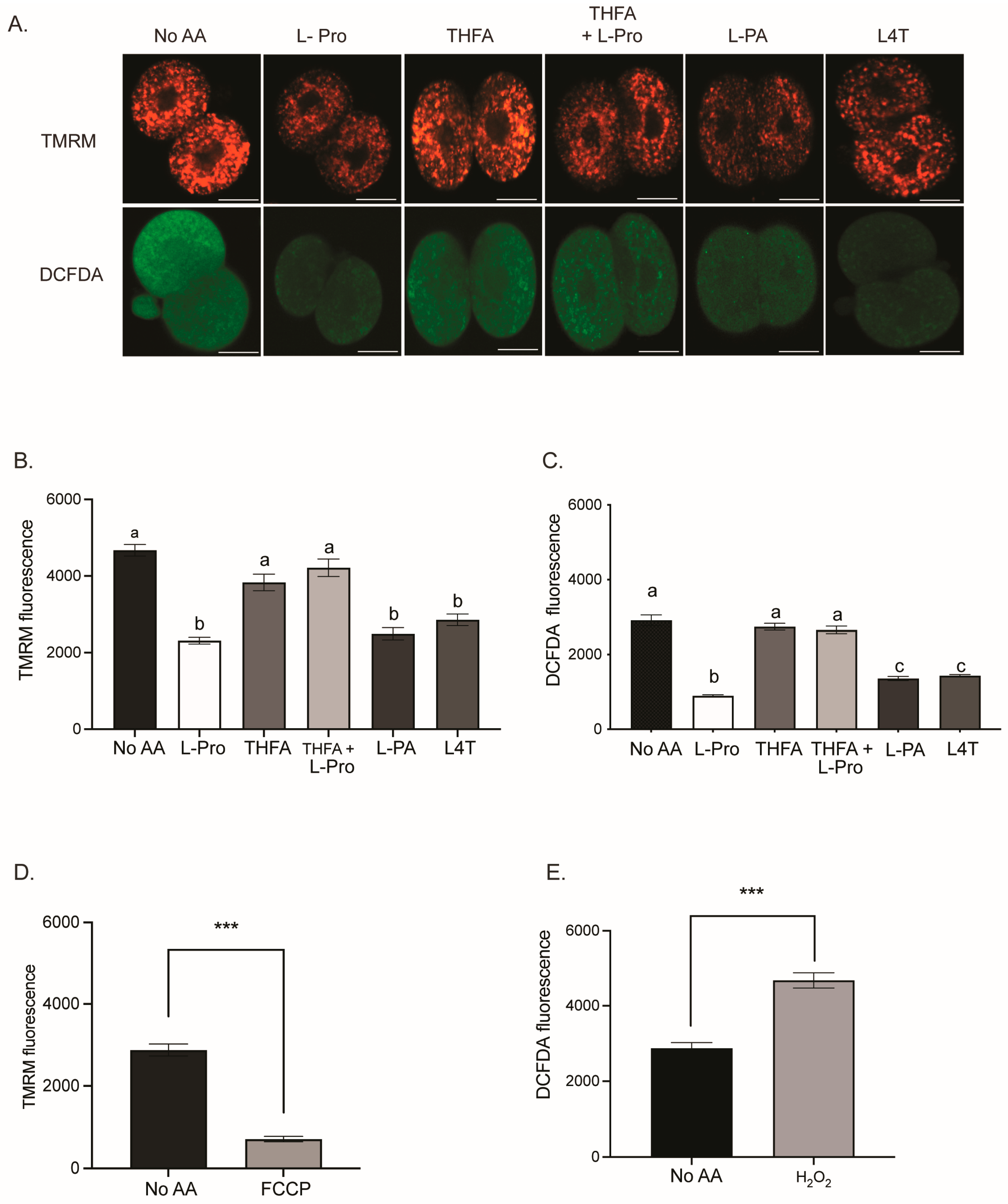
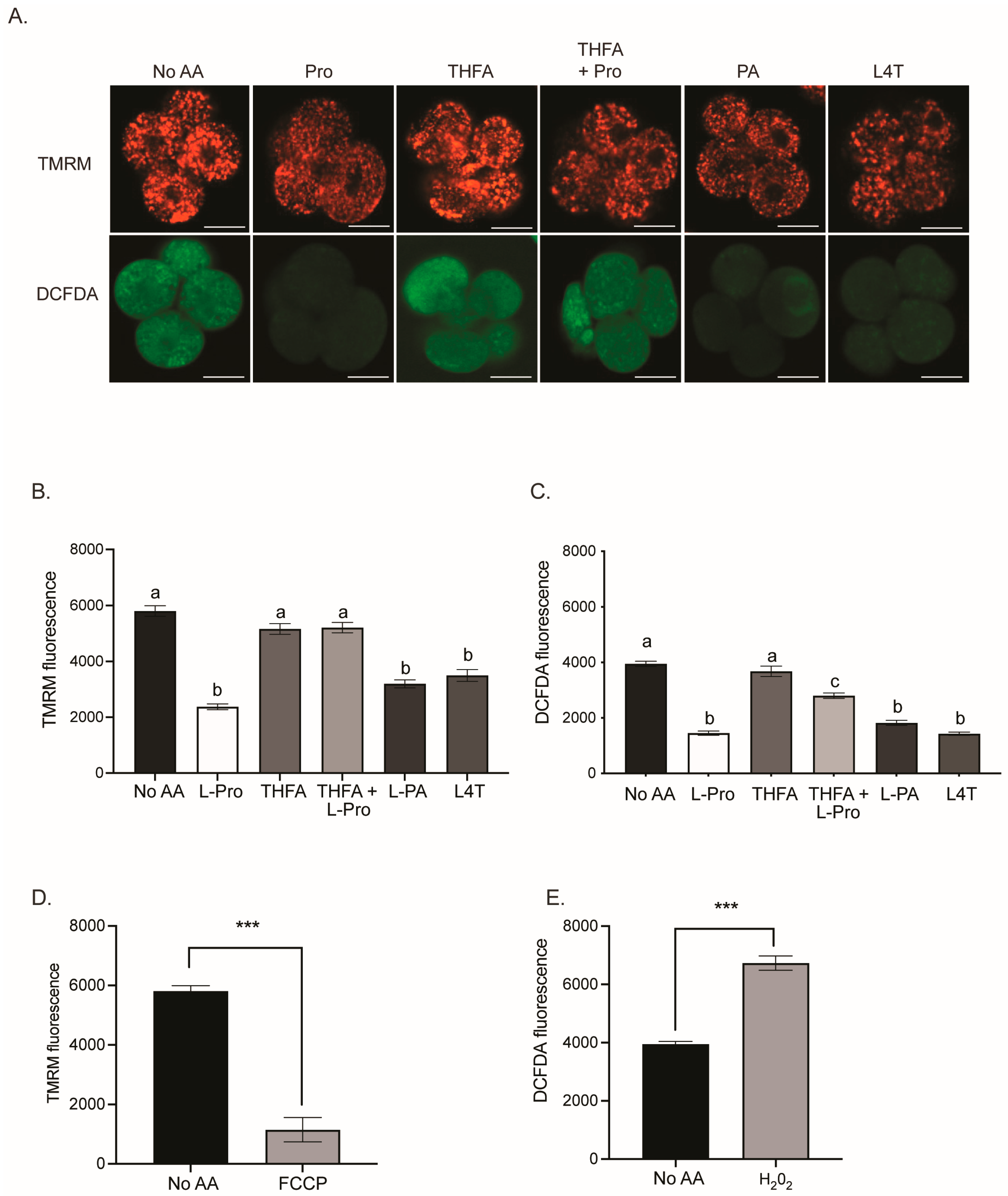
3.3. Pro and Analogues PA and L4T Decrease Mitochondrial Activity and ROS in 2- and 4-Cell Embryos
4. Discussion
4.1. Reduction in Mitochondrial Activity Reduces ROS
4.2. Direct Scavenging of ROS
4.3. Reduction of ROS through Production of GSH
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Takahashi, M. Oxidative stress and redox regulation on in vitro development of mammalian embryos. J. Reprod. Dev 2012, 58, 1–9. [Google Scholar] [CrossRef]
- Harlow, G.; Quinn, P. Development of preimplantation mouse embryos in vivo and in vitro. Aust. J. Biol. Sci. 1982, 35, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Baumann, C.G.; Morris, D.G.; Sreenan, J.M.; Leese, H.J. The quiet embryo hypothesis: Molecular characteristics favoring viability. Mol. Reprod. Dev. 2007, 74, 1345–1353. [Google Scholar] [CrossRef] [PubMed]
- Zamfirescu, R.C.; Day, M.L.; Morris, M.B. mTORC1/2 signaling is downregulated by amino acid-free culture of mouse preimplantation embryos and is only partially restored by amino acid readdition. Am. J. Physiol.-Cell Physiol. 2021, 320, C30–C44. [Google Scholar] [CrossRef] [PubMed]
- Gardner, D.K.; Sakkas, D. Mouse embryo cleavage, metabolism and viability: Role of medium composition. Hum. Reprod. 1993, 8, 288–295. [Google Scholar] [CrossRef] [PubMed]
- Gardner, D.K. Changes in requirements and utilization of nutrients during mammalian preimplantation embryo development and their significance in embryo culture. Theriogenology 1998, 49, 83–102. [Google Scholar] [CrossRef]
- Summers, M.C.; Biggers, J.D. Chemically defined media and the culture of mammalian preimplantation embryos: Historical perspective and current issues. Hum. Reprod. Update 2003, 9, 557–582. [Google Scholar] [CrossRef]
- Nasr-Esfahani, M.H.; Aitken, J.R.; Johnson, M.H. Hydrogen peroxide levels in mouse oocytes and early cleavage stage embryos developed in vitro or in vivo. Development 1990, 109, 501–507. [Google Scholar] [CrossRef]
- Goto, Y.; Noda, Y.; Narimoto, K.; Umaoka, Y.; Mori, T. Oxidative stress on mouse embryo development in vitro. Free Radic. Biol. Med. 1992, 13, 47–53. [Google Scholar] [CrossRef]
- Das, S.; Chattopadhyay, R.; Ghosh, S.; Ghosh, S.; Goswami, S.K.; Chakravarty, B.N.; Chaudhury, K. Reactive oxygen species level in follicular fluid—Embryo quality marker in IVF? Hum. Reprod. 2006, 21, 2403–2407. [Google Scholar] [CrossRef]
- Murphy, M.P. How mitochondria produce reactive oxygen species. Biochem. J. 2009, 417, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Harper, M.E.; Bevilacqua, L.; Hagopian, K.; Weindruch, R.; Ramsey, J.J. Ageing, oxidative stress, and mitochondrial uncoupling. Acta Physiol. Scand. 2004, 182, 321–331. [Google Scholar] [CrossRef] [PubMed]
- Hancock, C.N.; Liu, W.; Alvord, W.G.; Phang, J.M. Co-regulation of mitochondrial respiration by proline dehydrogenase/oxidase and succinate. Amino Acids 2016, 48, 859–872. [Google Scholar] [CrossRef] [PubMed]
- Hardy, M.L.; Day, M.L.; Morris, M.B. Redox Regulation and Oxidative stress in mammalian oocytes and embryos developed in vivo and in vitro. Int. J. Environ. Res. Public Health 2021, 18, 11374. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Fiskum, G.; Schubert, D. Generation of reactive oxygen species by the mitochondrial electron transport chain. J. Neurochem. 2002, 80, 780–787. [Google Scholar] [CrossRef]
- Velez-Pardo, C.; Morales, A.T.; Rio, M.J.D.; Olivera-Angel, M. Endogenously generated hydrogen peroxide induces apoptosis via mitochondrial damage independent of NF-κB and p53 activation in bovine embryos. Theriogenology 2007, 67, 1285–1296. [Google Scholar] [CrossRef]
- Betteridge, D.J. What is oxidative stress? Metabolism 2000, 49, 3–8. [Google Scholar] [CrossRef]
- Cadenas, E.; Davies, K.J. Mitochondrial free radical generation, oxidative stress, and aging. Free Radic. Biol. Med. 2000, 29, 222–230. [Google Scholar] [CrossRef]
- Sies, H.; Jones, D.P. Reactive oxygen species (ROS) as pleiotropic physiological signalling agents. Nat. Rev. Mol. Cell Biol. 2020, 21, 363–383. [Google Scholar] [CrossRef]
- Covarrubias, L.; Hernández-García, D.; Schnabel, D.; Salas-Vidal, E.; Castro-Obregón, S. Function of reactive oxygen species during animal development: Passive or active? Dev. Biol. 2008, 320, 1–11. [Google Scholar] [CrossRef]
- Veal, E.A.; Day, A.M.; Morgan, B.A. Hydrogen peroxide sensing and signaling. Mol. Cell 2007, 26, 1–14. [Google Scholar] [CrossRef] [PubMed]
- He, A.; Dean, J.M.; Lodhi, I.J. Peroxisomes as cellular adaptors to metabolic and environmental stress. Trends Cell Biol. 2021, 31, 656–670. [Google Scholar] [CrossRef]
- Ryter, S.W.; Kim, H.P.; Hoetzel, A.; Park, J.W.; Nakahira, K.; Wang, X.; Choi, A.M. Mechanisms of cell death in oxidative stress. Antioxid. Redox Sign. 2007, 9, 49–89. [Google Scholar] [CrossRef] [PubMed]
- Leese, H.J.; Guerif, F.; Allgar, V.; Brison, D.R.; Lundin, K.; Sturmey, R.G. Biological optimization, the Goldilocks principle, and how much is lagom in the preimplantation embryo. Mol. Reprod. Dev. 2016, 83, 748–754. [Google Scholar] [CrossRef]
- Dumollard, R.; Duchen, M.; Carroll, J. The role of mitochondrial function in the oocyte and embryo. Curr. Top. Dev. Biol. 2007, 77, 21–49. [Google Scholar] [CrossRef] [PubMed]
- Leese, H.J. Quiet please, do not disturb: A hypothesis of embryo metabolism and viability. Bioessays 2002, 24, 845–849. [Google Scholar] [CrossRef]
- Houghton, F.D.; Hawkhead, J.A.; Humpherson, P.G.; Hogg, J.E.; Balen, A.H.; Rutherford, A.J.; Leese, H.J. Non-invasive amino acid turnover predicts human embryo developmental capacity. Hum. Reprod. 2002, 17, 999–1005. [Google Scholar] [CrossRef]
- Leese, H.J.; Baumann, C.G.; Brison, D.R.; McEvoy, T.G.; Sturmey, R.G. Metabolism of the viable mammalian embryo: Quietness revisited. Mol. Hum. Reprod. 2008, 14, 667–672. [Google Scholar] [CrossRef]
- Larsson, N.-G.; Wang, J.; Wilhelmsson, H.; Oldfors, A.; Rustin, P.; Lewandoski, M.; Barsh, G.S.; Clayton, D.A. Mitochondrial transcription factor A is necessary for mtDNA maintance and embryogenesis in mice. Nat. Genet. 1998, 18, 231–236. [Google Scholar] [CrossRef]
- Houghton, F.D.; Leese, H.J. Metabolism and developmental competence of the preimplantation embryo. Eur. J. Obstet. Gynecol. Reprod. Biol. 2004, 115 (Suppl. S1), S92–S96. [Google Scholar] [CrossRef]
- Picton, H.M.; Elder, K.; Houghton, F.D.; Hawkhead, J.A.; Rutherford, A.J.; Hogg, J.E.; Leese, H.J.; Harris, S.E. Association between amino acid turnover and chromosome aneuploidy during human preimplantation embryo development in vitro. Mol. Hum. Reprod. 2010, 16, 557–569. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.; Han, H.B.; Tian, X.Z.; Tan, D.X.; Wang, L.; Zhou, G.B.; Zhu, S.E.; Liu, G.S. Melatonin promotes embryonic development and reduces reactive oxygen species in vitrified mouse 2-cell embryos. J. Pineal Res. 2012, 52, 305–311. [Google Scholar] [CrossRef] [PubMed]
- Cavallari, F.d.C.; Leal, C.L.V.; Roth, Z.; Hansen, P.J. Effects of melatonin on production of reactive oxygen species and developmental competence of bovine oocytes exposed to heat shock and oxidative stress during in vitro maturation. Zygote 2019, 27, 180–186. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.-J.; Pang, Y.-W.; Liu, Y.; Hao, H.-S.; Zhao, X.-M.; Qin, T.; Zhu, H.-B.; Du, W.-H. Exogenous glutathione supplementation in culture medium improves the bovine embryo development after in vitro fertilization. Theriogenology 2015, 84, 716–723. [Google Scholar] [CrossRef]
- Castillo-Martin, M.; Bonet, S.; Morató, R.; Yeste, M. Comparative effects of adding β-mercaptoethanol or L-ascorbic acid to culture or vitrification–warming media on IVF porcine embryos. Reprod. Fertil. Dev. 2014, 26, 875–882. [Google Scholar] [CrossRef]
- Lee, T.-H.; Lee, M.-S.; Liu, C.-H.; Tsao, H.-M.; Huang, C.-C.; Yang, Y.-S. The association between microenvironmental reactive oxygen species and embryo development in assisted reproduction technology cycles. Reprod. Sci. 2012, 19, 725–732. [Google Scholar] [CrossRef]
- Ho, Y.; Wigglesworth, K.; Eppig, J.J.; Schultz, R.M. Preimplantation development of mouse embryos in KSOM: Augmentation by amino acids and analysis of gene expression. Mol. Reprod. Dev. 1995, 41, 232–238. [Google Scholar] [CrossRef]
- Morris, M.B.; Ozsoy, S.; Zada, M.; Zada, M.; Zamfirescu, R.C.; Todorova, M.G.; Day, M.L. Selected amino acids promote mouse pre-implantation embryo development in a growth factor-like manner. Front. Physiol. 2020, 11, 140. [Google Scholar] [CrossRef]
- Treleaven, T.; Hardy, M.L.M.; Guttman-Jones, M.; Morris, M.B.; Day, M.L. In vitro fertilisation of mouse oocytes in L-proline and L-pipecolic acid improves subsequent development. Cells 2021, 10, 1352. [Google Scholar] [CrossRef]
- Van Winkle, L.J. Amino acid transport regulation and early embryo development. Biol. Reprod. 2001, 64, 1–12. [Google Scholar] [CrossRef]
- Glover, H.J.; Holliday, H.; Shparberg, R.A.; Winkler, D.; Day, M.; Morris, M.B. Signalling pathway crosstalk stimulated by L-proline drives mouse embryonic stem cells to primitive-ectoderm-like cells. Development 2023, 150, dev201704. [Google Scholar] [CrossRef]
- Aguilar, J.; Reyley, M. The uterine tubal fluid: Secretion, composition and biological effects. Anim. Reprod. 2018, 2, 91–105. [Google Scholar] [CrossRef]
- Guerin, P.; El Mouatassim, S.; Menezo, Y. Oxidative stress and protection against reactive oxygen species in the pre-implantation embryo and its surroundings. Hum. Reprod. Update 2001, 7, 175–189. [Google Scholar] [CrossRef] [PubMed]
- Józwik, M.; Józwik, M.; Teng, C.; Battaglia, F.C. Amino acid, ammonia and urea concentrations in human pre-ovulatory ovarian follicular fluid. Hum. Reprod. 2006, 21, 2776–2782. [Google Scholar] [CrossRef] [PubMed]
- Elhassan, Y.; Wu, G.; Leanez, A.; Tasca, R.; Watson, A.; Westhusin, M. Amino acid concentrations in fluids from the bovine oviduct and uterus and in KSOM-based culture media. Theriogenology 2001, 55, 1907–1918. [Google Scholar] [CrossRef]
- Anas, M.-K.I.; Lee, M.B.; Zhou, C.; Hammer, M.-A.; Slow, S.; Karmouch, J.; Liu, X.J.; Bröer, S.; Lever, M.; Baltz, J.M. SIT1 is a betaine/proline transporter that is activated in mouse eggs after fertilization and functions until the 2-cell stage. Development 2008, 135, 4123–4130. [Google Scholar] [CrossRef]
- Anas, M.K.; Hammer, M.A.; Lever, M.; Stanton, J.A.L.; Baltz, J.M. The organic osmolytes betaine and proline are transported by a shared system in early preimplantation mouse embryos. J. Cell. Physiol. 2007, 210, 266–277. [Google Scholar] [CrossRef]
- Van Winkle, L.J.; Haghighat, N.; Campione, A.L.; Gorman, J.M. Glycine transport in mouse eggs ad preimplantation conceptuses. BBA-Biomembranes 1988, 941, 241–256. [Google Scholar] [CrossRef]
- Dawson, K.M.; Baltz, J.M. Organic osmolytes and embryos: Substrates of the Gly and β transport systems protect mouse zygotes against the effects of raised osmolarity. Biol. Reprod. 1997, 56, 1550–1558. [Google Scholar] [CrossRef]
- Tan, B.S.N.; Lonic, A.; Morris, M.B.; Rathjen, P.D.; Rathjen, J. The amino acid transporter SNAT2 mediates L-proline-induced differentiation of ES cells. Am. J. Physiol.-Cell Physiol. 2011, 300, C1270–C1279. [Google Scholar] [CrossRef]
- Bloise, E.; Lin, W.; Liu, X.; Simbulan, R.; Kolahi, K.S.; Petraglia, F.; Maltepe, E.; Donjacour, A.; Rinaudo, P. Impaired placental nutrient transport in mice generated by in vitro fertilization. Endocrinology 2012, 153, 3457–3467. [Google Scholar] [CrossRef] [PubMed]
- Treleaven, T.; Zada, M.; Nagarajah, R.; Bailey, C.G.; Rasko, J.E.J.; Morris, M.B.; Day, M.L. Stage-specific L-Proline uptake by amino acid transporter Slc6a19/B0AT1 is required for optimal preimplantation embryo development in mice. Cells 2023, 12, 18. [Google Scholar] [CrossRef] [PubMed]
- Bröer, S.; Gether, U. The solute carrier 6 family of transporters. Br. J. Pharmacol. 2012, 167, 256–278. [Google Scholar] [CrossRef] [PubMed]
- Comes, S.; Gagliardi, M.; Laprano, N.; Fico, A.; Cimmino, A.; Palamidessi, A.; De Cesare, D.; De Falco, S.; Angelini, C.; Scita, G.; et al. L-Proline induces a mesenchymal-like invasive program in embryonic stem cells by remodeling H3K9 and H3K36 methylation. Stem Cell Rep. 2013, 1, 307–321. [Google Scholar] [CrossRef]
- Phang, J.M. Proline metabolism in cell regulation and cancer biology: Recent advances and hypotheses. Antioxid. Redox Sign. 2019, 30, 635–649. [Google Scholar] [CrossRef]
- D’Aniello, C.; Fico, A.; Casalino, L.; Guardiola, O.; Di Napoli, G.; Cermola, F.; De Cesare, D.; Tate, R.; Cobellis, G.; Patriarca, E. A novel autoregulatory loop between the Gcn2-Atf4 pathway and L-Proline metabolism controls stem cell identity. Cell Death Differ. 2015, 22, 1094–1105. [Google Scholar] [CrossRef]
- Washington, J.M.; Rathjen, J.; Felquer, F.; Lonic, A.; Bettess, M.D.; Hamra, N.; Semendric, L.; Tan, B.S.N.; Lake, J.-A.; Keough, R.A.; et al. L-Proline induces differentiation of ES cells: A novel role for an amino acid in the regulation of pluripotent cells in culture. Am. J. Physiol.-Cell Physiol. 2010, 298, C982–C992. [Google Scholar] [CrossRef]
- Tanner, J.J.; Fendt, S.-M.; Becker, D.F. The proline cycle as a potential cancer therapy target. Biochemistry 2018, 57, 3433–3444. [Google Scholar] [CrossRef]
- Phang, J.M.; Yeh, G.C.; Hagedorn, C.H. The intercellular proline cycle. Life Sci. 1981, 28, 53–58. [Google Scholar] [CrossRef]
- Phang, J.M. The proline regulatory axis and cancer. Front. Oncol. 2012, 2, 60. [Google Scholar] [CrossRef]
- Liang, X.; Zhang, L.; Natarajan, S.K.; Becker, D.F. Proline mechanisms of stress survival. Antioxid. Redox Sign. 2013, 19, 998–1011. [Google Scholar] [CrossRef]
- Matysik, J.; Alia; Bhalu, B.; Mohanty, P. Molecular mechanisms of quenching of reactive oxygen species by proline under stress in plants. Curr. Sci. 2002, 82, 525–532. [Google Scholar]
- Krishnan, N.; Dickman, M.B.; Becker, D.F. Proline modulates the intracellular redox environment and protects mammalian cells against oxidative stress. Free Radic. Biol. Med. 2008, 44, 671–681. [Google Scholar] [CrossRef]
- Tan, B.S.N.; Kwek, J.; Wong, C.K.E.; Saner, N.J.; Yap, C.; Felquer, F.; Morris, M.B.; Gardner, D.K.; Rathjen, P.D.; Rathjen, J. Src family kinases and p38 mitogen-activated protein kinases regulate pluripotent cell differentiation in culture. PLoS ONE 2016, 11, e0163244. [Google Scholar] [CrossRef] [PubMed]
- Feng, C.; Zhu, Z.; Bai, W.; Li, R.; Zheng, Y.; Tian, X.E.; Wu, D.; Lu, H.; Wang, Y.; Zeng, W. Proline protects boar sperm against oxidative stress through proline dehydrogenase-mediated metabolism and the amine structure of pyrrolidine. Animals 2020, 10, 1549. [Google Scholar] [CrossRef] [PubMed]
- Creed, S.; McKenzie, M. Measurement of Mitochondrial Membrane Potential with the fluorescent dye tetramethylrhodamine methyl ester (TMRM). In Cancer Metabolism: Methods and Protocols; Haznadar, M., Ed.; Springer: New York, NY, USA, 2019; pp. 69–76. [Google Scholar] [CrossRef]
- Szychowski, K.A.; Rybczyńska-Tkaczyk, K.; Leja, M.L.; Wójtowicz, A.K.; Gmiński, J. Tetrabromobisphenol A (TBBPA)-stimulated reactive oxygen species (ROS) production in cell-free model using the 2′, 7′-dichlorodihydrofluorescein diacetate (H 2 DCFDA) assay—Limitations of method. Environ. Sci. Pollut. Res. 2016, 23, 12246–12252. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Cardenas, L.; Villanueva-Chimal, E.; Salinas, L.; Jose-Nunez, C.; de Gomez Puyou, M.T.; Navarro, R. Caenorhabditis elegans ATPase inhibitor factor 1 (IF1) MAI-2 preserves the mitochondrial membrane potential (Δψm) and is important to induce germ cell apoptosis. PLoS ONE 2017, 12, e0181984. [Google Scholar] [CrossRef]
- Bendich, A.J. Mitochondrial DNA, chloroplast DNA and the origins of development in eukaryotic organisms. Biol. Direct 2010, 5, 42. [Google Scholar] [CrossRef]
- Steeves, C.L.; Hammer, M.-A.; Walker, G.B.; Rae, D.; Stewart, N.A.; Baltz, J.M. The glycine neurotransmitter transporter GLYT1 is an organic osmolyte transporter regulating cell volume in cleavage-stage embryos. Proc. Natl. Acad. Sci. USA 2003, 100, 13982–13987. [Google Scholar] [CrossRef]
- Kowalczuk, S.; Broer, A.; Munzinger, M.; Tietze, N.; Klingel, K.; Broer, S. Molecular cloning of the mouse IMINO system: An Na+- and Cl−-dependent proline transporter. Biochem. J. 2005, 386, 417–422. [Google Scholar] [CrossRef]
- Takanaga, H.; Mackenzie, B.; Suzuki, Y.; Hediger, M.A. Identification of mammalian proline transporter SIT1 (SLC6A20) with characteristics of classical system imino. J. Biol. Chem. 2005, 280, 8974–8984. [Google Scholar] [CrossRef] [PubMed]
- Metzner, L.; Neubert, K.; Brandsch, M. Substrate specificity of the amino acid transporter PAT1. Amino Acids 2006, 31, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Edwards, N.; Anderson, C.M.; Gatfield, K.M.; Jevons, M.P.; Ganapathy, V.; Thwaites, D.T. Amino acid derivatives are substrates or non-transported inhibitors of the amino acid transporter PAT2 (slc36a2). BBA-Biomembranes 2011, 1808, 260–270. [Google Scholar] [CrossRef] [PubMed]
- Bartosz, G. Reactive oxygen species: Destroyers or messengers? Biochem. Pharmacol. 2009, 77, 1303–1315. [Google Scholar] [CrossRef]
- Di Meo, S.; Reed, T.T.; Venditti, P.; Victor, V.M. Role of ROS and RNS sources in physiological and pathological conditions. Oxid. Med. Cell. Longev. 2016, 2016, 1245049. [Google Scholar] [CrossRef]
- Zhang, L.; Xue, X.; Yan, J.; Yan, L.Y.; Jin, X.H.; Zhu, X.H.; He, Z.Z.; Liu, J.; Li, R.; Qiao, J. L-proline: A highly effective cryoprotectant for mouse oocyte vitrification. Sci. Rep. 2016, 6, 26326. [Google Scholar] [CrossRef]
- Delic, V.; Griffin, J.W.D.; Zivkovic, S.; Zhang, Y.; Phan, T.A.; Gong, H.; Chaput, D.; Reynes, C.; Dinh, V.B.; Cruz, J.; et al. Individual amino acid supplementation can improve energy metabolism and decrease ROS production in neuronal cells overexpressing alpha-synuclein. Neuromol. Med. 2017, 19, 322–344. [Google Scholar] [CrossRef]
- Liu, W.; Phang, J.M. Proline dehydrogenase (oxidase) in cancer. Biofactors 2012, 38, 398–406. [Google Scholar] [CrossRef]
- Adams, E. Metabolism of proline and of hydroxyproline. Int. Rev. Connect. Tissue Res. 1970, 5, 1–91. [Google Scholar] [CrossRef]
- Donald, S.P.; Sun, X.-Y.; Hu, C.-A.A.; Yu, J.; Mei, J.M.; Valle, D.; Phang, J.M. Proline oxidase, encoded by p53-induced gene-6, catalyzes the generation of proline-dependent reactive oxygen species. Cancer Res. 2001, 61, 1810–1815. [Google Scholar] [PubMed]
- Liu, N.; Yang, Y.; Si, X.; Jia, H.; Zhang, Y.; Jiang, D.; Dai, Z.; Wu, Z. L-Proline activates mammalian target of rapamycin Complex 1 and modulates redox environment in porcine trophectoderm cells. Biomolecules 2021, 11, 742. [Google Scholar] [CrossRef] [PubMed]
- Pena, I.A.; Marques, L.A.; Laranjeira, Â.B.A.; Yunes, J.A.; Eberlin, M.N.; MacKenzie, A.; Arruda, P. Mouse lysine catabolism to aminoadipate occurs primarily through the saccharopine pathway; implications for pyridoxine dependent epilepsy (PDE). BBA-Mol. Basis Dis. 2017, 1863, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Dodt, G.; Kim, D.G.; Reimann, S.A.; Reuber, B.E.; McCabe, K.; Gould, S.J.; Mihalik, S.J. L-pipecolic acid oxidase, a human enzyme essential for the degradation of L-pipecolic acid, is most similar to the monomeric sarcosine oxidases. Biochem. J. 2000, 345, 487–494. [Google Scholar] [CrossRef]
- Natarajan, S.K.; Muthukrishnan, E.; Khalimonchuk, O.; Mott, J.L.; Becker, D.F. Evidence for pipecolate oxidase in mediating protection against hydrogen peroxide stress. J. Cell. Biochem. 2017, 118, 1678–1688. [Google Scholar] [CrossRef] [PubMed]
- Reuber, B.E.; Karl, C.; Reimann, S.A.; Mihalik, S.J.; Dodt, G. Cloning and functional expression of a mammalian gene for a peroxisomal sarcosine oxidase. J. Biol. Chem. 1997, 272, 6766–6776. [Google Scholar] [CrossRef]
- Lahham, M.; Jha, S.; Goj, D.; Macheroux, P.; Wallner, S. The family of sarcosine oxidases: Same reaction, different products. Arch. Biochem. Biophys. 2021, 704, 108868. [Google Scholar] [CrossRef]
- Zhu, W.; Gincherman, Y.; Docherty, P.; Spilling, C.D.; Becker, D.F. Effects of proline analog binding on the spectroscopic and redox properties of PutA. Arch. Biochem. Biophys. 2002, 408, 131–136. [Google Scholar] [CrossRef]
- Jeelani, G.; Sato, D.; Soga, T.; Watanabe, H.; Nozaki, T. Mass spectrometric analysis of L-cysteine metabolism: Physiological role and fate of L-cysteine in the enteric protozoan parasite Entamoeba histolytica. MBio 2014, 5, e01995-14. [Google Scholar] [CrossRef]
- Lesarri, A.; Mata, S.; Cocinero, E.J.; Blanco, S.; Lopez, J.C.; Alonso, J.L. The structure of neutral proline. Angew. Chem. Int. Ed. Engl. 2002, 41, 4673–4676. [Google Scholar] [CrossRef]
- Weber, H.U.; Fleming, J.F.; Miquel, J. Thiazolidine-4-carboxylic acid, a physiologic sulfhydryl antioxidant with potential value in geriatric medicine. Arch. Gerontol. Geriatr. 1982, 1, 299–310. [Google Scholar] [CrossRef]
- Ham, Y.-H.; Jason Chan, K.; Chan, W. Thioproline serves as an efficient antioxidant protecting human cells from oxidative stress and improves cell viability. Chem. Res. Toxicol. 2020, 33, 1815–1821. [Google Scholar] [CrossRef] [PubMed]
- Holtzapple, P.; Genel, M.; Rea, C.; Segal, S. Metabolism and uptake of L-proline by human kidney cortex. Pediatr. Res. 1973, 7, 818–825. [Google Scholar] [CrossRef]
- Wang, J.; Xue, Z.; Hua, C.; Lin, J.; Shen, Z.; Song, Y.; Ying, H.; Lv, Q.; Wang, M.; Zhou, B. Metabolomic analysis of the ameliorative effect of enhanced proline metabolism on hypoxia-induced injury in cardiomyocytes. Oxid. Med. Cell. Longev. 2020, 2020, 8866946. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Si, X.; Ji, Y.; Yang, Q.; Bai, J.; He, Y.; Jia, H.; Song, Z.; Chen, J.; Yang, L.; et al. L-Proline improves the cytoplasmic maturation of mouse oocyte by regulating glutathione-related redox homeostasis. Theriogenology 2023, 195, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Van Winkle, L.J. Perspective: Might maternal dietary monosodium glutamate (MSG) consumption impact pre- and peri-implantation embryos and their subsequent development? Int. J. Environ. Res. Public Health 2022, 19, 13611. [Google Scholar] [CrossRef] [PubMed]
- Natarajan, S.K.; Zhu, W.; Liang, X.; Zhang, L.; Demers, A.J.; Zimmerman, M.C.; Simpson, M.A.; Becker, D.F. Proline dehydrogenase is essential for proline protection against hydrogen peroxide-induced cell death. Free Radic. Biol. Med. 2012, 53, 1181–1191. [Google Scholar] [CrossRef]
- Papes, F.; Surpili, M.J.; Langone, F.; Trigo, J.R.; Arruda, P. The essential amino acid lysine acts as precursor of glutamate in the mammalian central nervous system. FEBS Lett. 2001, 488, 34–38. [Google Scholar] [CrossRef]
- Bayram, F.E.Ö.; Sipahi, H.; Acar, E.T.; Ulugöl, R.K.; Buran, K.; Akgün, H. The cysteine releasing pattern of some antioxidant thiazolidine-4-carboxylic acids. Eur. J. Med. Chem. 2016, 114, 337–344. [Google Scholar] [CrossRef]
- Banjac, A.; Perisic, T.; Sato, H.; Seiler, A.; Bannai, S.; Weiss, N.; Kölle, P.; Tschoep, K.; Issels, R.; Daniel, P. The cystine/cysteine cycle: A redox cycle regulating susceptibility versus resistance to cell death. Oncogene 2008, 27, 1618–1628. [Google Scholar] [CrossRef]
- Shparberg, R.A.; Glover, H.J.; Morris, M.B. Modeling mammalian commitment to the neural lineage using embryos and embryonic stem cells. Front. Physiol. 2019, 10, 705. [Google Scholar] [CrossRef]
- Lonic, A. Molecular Mechanism of L-Proline Induced EPL-Cell Formation. Ph.D. Thesis, University of Adelaide, Adelaide, SA, Australia, 2007. [Google Scholar]
- Zhong, X.; Zeng, M.; Bian, H.; Zhong, C.; Xiao, F. An evaluation of the protective role of vitamin C in reactive oxygen species-induced hepatotoxicity due to hexavalent chromium in vitro and in vivo. J. Occup. Med. Toxicol. 2017, 12, 15. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hardy, M.L.M.; Lakhiani, D.; Morris, M.B.; Day, M.L. Proline and Proline Analogues Improve Development of Mouse Preimplantation Embryos by Protecting Them against Oxidative Stress. Cells 2023, 12, 2640. https://doi.org/10.3390/cells12222640
Hardy MLM, Lakhiani D, Morris MB, Day ML. Proline and Proline Analogues Improve Development of Mouse Preimplantation Embryos by Protecting Them against Oxidative Stress. Cells. 2023; 12(22):2640. https://doi.org/10.3390/cells12222640
Chicago/Turabian StyleHardy, Madeleine L. M., Dheerja Lakhiani, Michael B. Morris, and Margot L. Day. 2023. "Proline and Proline Analogues Improve Development of Mouse Preimplantation Embryos by Protecting Them against Oxidative Stress" Cells 12, no. 22: 2640. https://doi.org/10.3390/cells12222640