
The Multitasker Protein: A Look at the Multiple Capabilities of NUMB

1
Instituto de Biomedicina de Sevilla (IBIS), Hospital Universitario Virgen del Rocío (HUVR), Consejo Superior de Investigaciones Científicas, Universidad de Sevilla, 41013 Sevilla, Spain
2
CIBERONC, Instituto de Salud Carlos III, 28029 Madrid, Spain
3
Departamento de Bioquímica Vegetal y Biología Molecular, Universidad de Sevilla, 41012 Sevilla, Spain
*
Author to whom correspondence should be addressed.
Cells 2023, 12(2), 333; https://doi.org/10.3390/cells12020333
Submission received: 1 December 2022
/
Revised: 8 January 2023
/
Accepted: 13 January 2023
/
Published: 15 January 2023
(This article belongs to the Special Issue Cancers: Genetics and Cellular Perspective)
{kind=link}
{kind=link}
{kind=link}
Abstract
:NUMB, a plasma membrane-associated protein originally described in Drosophila, is involved in determining cell function and fate during early stages of development. It is secreted asymmetrically in dividing cells, with one daughter cell inheriting NUMB and the other inheriting its antagonist, NOTCH. NUMB has been proposed as a polarizing agent and has multiple functions, including endocytosis and serving as an adaptor in various cellular pathways such as NOTCH, Hedgehog, and the P53-MDM2 axis. Due to its role in maintaining cellular homeostasis, it has been suggested that NUMB may be involved in various human pathologies such as cancer and Alzheimer’s disease. Further research on NUMB could aid in understanding disease mechanisms and advancing the field of personalized medicine and the development of new therapies.
1. Introduction
The study of the bases of development has been shown to be an effective way to understand the molecular basis of human pathologies. Investigating proteins that are involved in human diseases and play a role in development and cell fate could be key to making connections and advances by understanding how their presence and function modify cell behavior. One such multitasking proteins is NUMB [1], which has a wide range of functions in various species [2]. It is a cell membrane-associated protein that plays a crucial role in determining cell fate. Originally, NUMB was first described as an antagonist of the NOTCH membrane receptor in sensory organ precursor cells (SOPs) in Drosophila melanogaster [3]. It is secreted asymmetrically during cell division, producing two types of cells: one that retains the characteristics of the parent cell and one that is capable of differentiation [4,5].
NUMB is highly evolutionarily conserved. Since its discovery in Drosophila, two homologous proteins, NUMB and NUMBL, have been found in multiple mammals, although most of the research has been carried out in mice (mNumb/mNumbl) and humans (NUMB/NUMBL) [6,7]. NUMBL performs many functions that overlap with NUMB, although certain aspects of NUMBL remain to be understood [6]. NUMB is involved in important cellular processes, such as protein labeling for endocytosis, ubiquitination, cell adhesion, migration, and asymmetric cell division. Unlike NUMB, NUMBL is secreted symmetrically to daughter cells [7]. The role of NUMB as a cell fate determinant has been related to processes where there is a balance between self-renewal and cell fate determination [8,9,10,11,12,13,14,15]. This has led to its proposed involvement in pathologies related to the disruption of this balance, such as cancer or Alzheimer’s disease [16,17,18,19,20,21,22,23]. NUMB has been proposed to be involved in the regulation of multiple cellular pathways commonly altered in cancer, such as Wnt, Notch, or Hedgehog, due to its interactions with a wide variety of proteins [24,25,26,27,28]. It is also thought to be involved in the maintenance of the cancer stem cell (CSC) pool [28,29]. NUMB has also been linked to Alzheimer’s disease, where it plays a role as an adaptor of amyloid precursor protein (APP), an essential protein in the pathogenesis of the disease [23]. Recently, differential expression of NUMB has been proposed as a prognostic factor in various types of cancer [2,17,30,31,32,33]. It has been described that eukaryotic NUMB mRNA can undergo alternative splicing to produce at least four different isoforms at the protein level (called p72, p71, p66, and p65) produced by the inclusion or exclusion of exons 3 and 9, making NUMB an even more multifunctional protein [34,35,36]. This has led to the proposed prognostic value of each of them in some cases [34,37,38,39,40]. Because of its adaptor role, NUMB has also been proposed as a therapeutic target, not only in different types of cancer but also in various pathologies such as Alzheimer’s disease, among others [41,42,43,44,45,46].
In this work, we review the various roles of NUMB in the cell as an adaptor, polarization agent, and endocytic protein, and how these functions are involved in the development of pathologies such as Alzheimer’s disease and cancer. We also detail the possible mechanisms responsible for regulating NUMB and its potential as a target for the development of therapies for various diseases, as well as its usefulness as a biomarker.
2. NUMB Gene
NUMB, also known as S171, C14orf41, or C14-5527, is a gene located on the 14q24.3 chromosomal region that encodes an endocytic protein of the same name [47,48]. It is one of 19 different transcripts derived from genes in the AD3 region of chromosome 14, which is associated with familial Alzheimer’s disease [28,49]. This form of dementia develops slowly and progressively worsens, causing problems with memory, coordination, and thinking [50]. It is characterized by the aggregation of β-amyloid peptides, which are generated from APP by sequential cleavage, first by β-secretase and then by γ-secretase [51]. Although most cases of Alzheimer’s disease have a sporadic onset, familial Alzheimer’s disease is mainly caused by mutations in three genes: APP, presenilin 1 (PSEN1), and presenilin 2 (PSEN2) [52]. In fact, the AD3 region contains, among others, the PSEN1 gene, which encodes the catalytic domain of γ-secretase that is also responsible for cleaving the NOTCH receptor [28,49].
NUMB is expressed in different adult human tissues, with the highest levels found in the blood, lung, and gallbladder and the lowest in the pancreas [48,53]. However, during development, NUMB expression varies in a stage-dependent manner, peaking at the two-cell embryo stage and gradually decreasing in later stages before blastocyst formation [54]. There are no known germline diseases due to single nucleotide polymorphism changes in the NUMB gene [55]. In addition, NUMB mutations are usually not point mutations, but rather amplifications or deep deletions [56]. However, little is known about the epigenetic status of NUMB in neurogenesis and development. The NUMB promoter appears to be hypermethylated in tumors, such as breast invasive carcinoma, lung adenocarcinoma, and colon adenocarcinoma, compared to normal tissue. On the other hand, its close homolog, NUMBL, appears to be hypomethylated in tumors such as colon and lung adenocarcinoma, compared to non-tumor tissue [6,57].
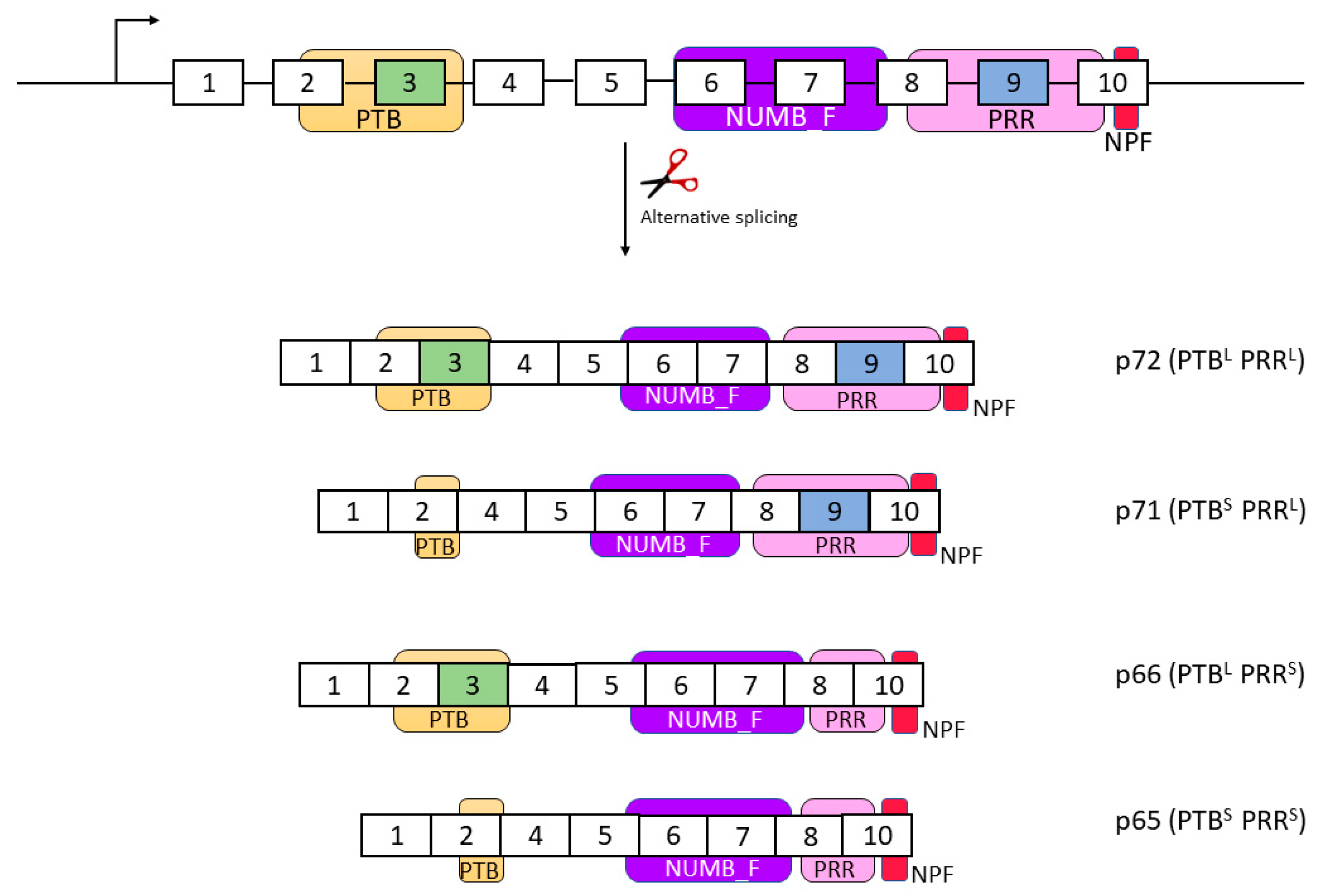
This gene is composed of nine exons that encode up to nine isoforms through alternative splicing. Only four of these isoforms encode a protein, which are named according to their molecular weight: p72 (variant 1), p66 (variant 2), p71 (variant 3), and p65 (variant 4) [35,58,59]. The longest transcript and isoform is p72, which includes all exons. Isoform p66 lacks exon 9, while isoform p71 lacks exon 3. The shortest variant is p65, which lacks both exons 3 and 9 (Figure 1) [35,48,59].
3. NUMB Protein
NUMB is a membrane-associated protein that is primarily found in the cell and peripheral membrane [48,53]. However, it is also believed to be present in the endosomal membrane, clathrin vesicles, the basolateral membrane, nuclei, and cytoplasm [58]. The N-terminal region of NUMB presents a phosphotyrosine binding (PTB) domain, while the C-terminal region presents an Asn-Pro-Phe (NPF) motif, which allows it to interact with the EH domains found in endocytic proteins [49,55,60,61]. In addition, NUMB has a domain located near the PTB domain, called NUMB-F, the function of which remains unknown [49,55]. NUMB also has a proline-rich region (PRR) that serves as an Src homology 3-binding domain. The PTB and PRR domains are affected by alternative splicing, as they include exons 3 and 9, respectively. As a result, the presence or absence of these exons can result in long or short PTB (PTBL/S) and long or short PRR (PRRL/S) domains in the isoforms [62,63] (Figure 1).
NUMB interacts with a variety of proteins, including REPS1 (RalBP1-associated Eps domain-containing protein 1), CTNNB1 (β-catenin 1), A4 (APP amyloid β precursor 4 protein), p53, and Mdm2/HDM2 (Murine/Human Double Minute 2) [39,48,58]. Furthermore, the interaction of NUMB with the oncogenic protein MAP17 leads to the aberrant activation of the NOTCH pathway and an increase in tumorigenic cell properties [28]. NUMB can also be post-translationally modified, mainly through phosphorylation and methylation, which can alter its ability to interact with other proteins. For example, NUMB phosphorylation at the Ser276 and/or Ser295 residues by CAMK-1 protein reduces the stability of one of the NUMB interactors, p53, by disrupting the NUMB-p53 interaction [64,65,66]. NUMB methylation by SET8 at Lys158 and Lys163 also causes dissociation of its interaction with p53 [67]. There are also isoform-specific modifications, as only p72 and p66, which contain the full PTB domain, can be ubiquitinated by the Ligand of the NUMB X (LNX) protein for subsequent degradation in the proteasome [53]. These facts make the NUMB interactome a very complex and dynamic network.
4. NUMB Is Involved in Asymmetric-Division Related Pathologies
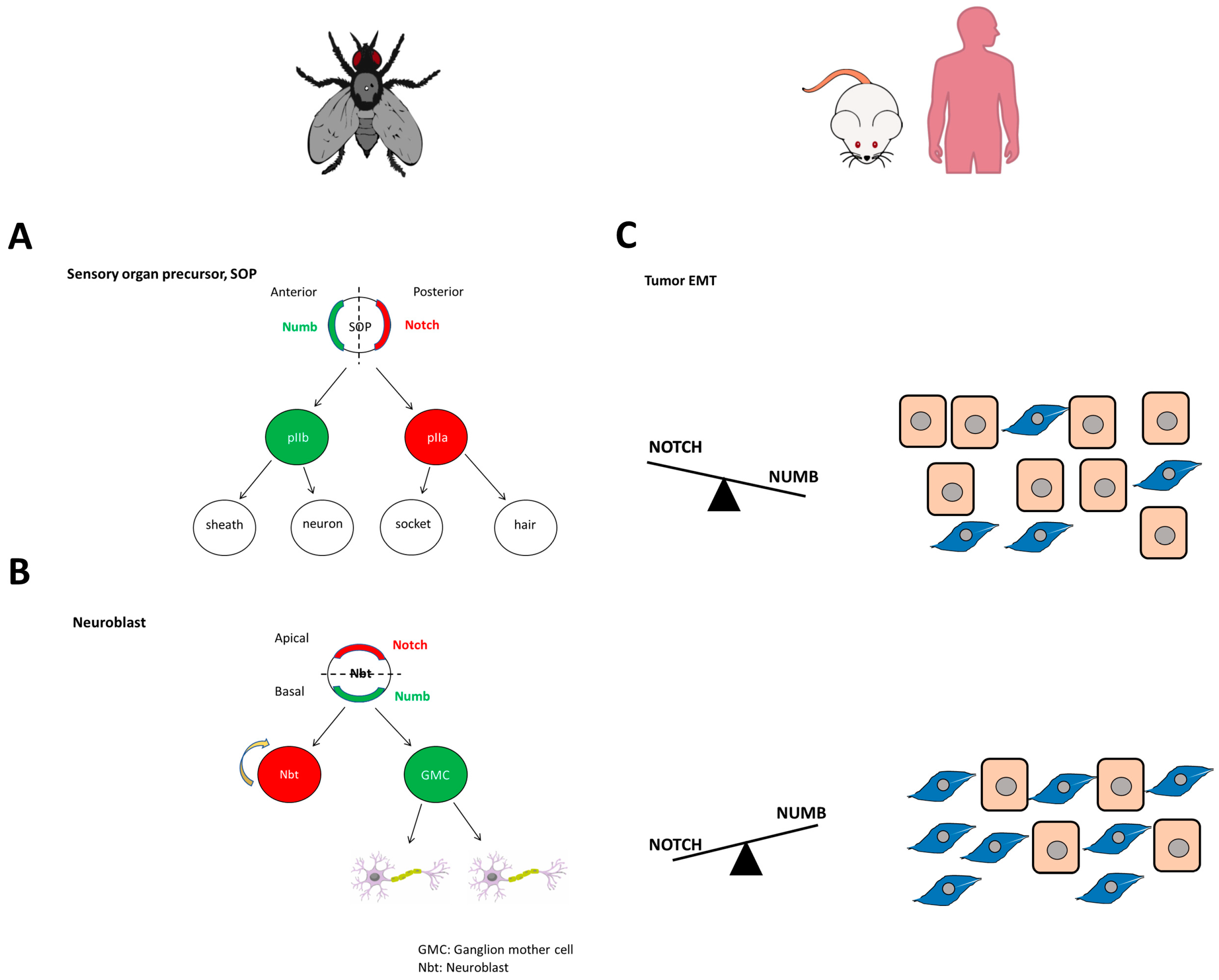
Asymmetric division plays a vital role in generating cell diversity [68]. This phenomenon was first found in Drosophila and Caenorhabditis elegans, where cell fate proteins are passed asymmetrically to daughter cells in precursor cell division [69]. The SOP in Drosophila is crucial for the development of the central and peripheral nervous system [68,70]. It undergoes rounds of asymmetric division to produce four daughter cells: sheath, neuron, socket, and hair [68,70]. NUMB is secreted in one of the daughter cells (anterior pI daughter cell b, pIIb), while its antagonist, NOTCH, is secreted in the other daughter cell (pIIa) [68,70,71]. This is due to NUMB being polarized to one side of the cell during mitosis, demonstrating its role as a cell fate determinant (Figure 2A) [68,70,71]. NUMB and NOTCH also control asymmetric division in neuroblasts, which inherit NOTCH, and neural stem cells (NSCs), also called ganglion mother cells, which inherit NUMB (Figure 2B) [72,73]. Par6, Baz, and aPKC, apical polarity proteins, are located on one side of the neuroblast, while Lgl (Lethal giant larvae), Dlg (Discs large), and NUMB are located at the basal pole [72,74]. The partition defective complex (Par3-Par6-aPKC) leads to polarized organization and asymmetric segregation of NUMB [75]. Additionally, Lgl is required for the proper asymmetric segregation of NUMB into daughter cells, as it promotes the formation of a basal crescent form of this protein [74]. However, proper positioning of NUMB and the orientation of the mitotic spindle also depend on the presence of the Inscuteable protein (Insc), a key component of the asymmetric segregation machinery in Drosophila [76,77]. Insc is located in the apical cortex of the cell before and during neuroblast mitosis, and it must interact with Bazooka to maintain the apical-basal polarity necessary for NUMB and other proteins to be asymmetrically segregated in metaphase [78]. Therefore, the relationship between apical-basal polarity and asymmetric segregation has been demonstrated [78]. Because NUMB interacts with other proteins through its PTB domain, it has been classified as a polarity marker [79,80,81].
The role of NUMB in maintaining neural progenitors has been well-stablished [72]. Both NUMB and NUMBL have been suggested to be involved in maintaining highly polarized radial glial cells and in cortical neurogenesis [72,82,83]. As a result, deleting NUMB and NUMBL in mice is embryonically lethal [84]. Therefore, to study the effects of NUMB and NUMBL deficiency in mice, a conditional NUMB–NUMBL knock-out model was created using Emx1-Cre (expression was induced on Day 9.5, avoiding early embryonic lethality). This resulted in disruption of the neuroepithelium, severe hydrocephalus, delayed cell cycle exit, impaired neural differentiation, and progenitor hyperproliferation [72,82].
Furthermore, NUMB has been shown to play an important role in the functions of the Par complex (as mentioned above) and cell-cell junctions, both processes commonly associated with epithelial-mesenchymal transition (EMT) [85,86,87]. In this process, epithelial cells lose their cell polarity and cell-cell adhesion, gaining both migratory and invasive properties to become mesenchymal stem cells [85,86,87].
NUMB has also been described as an essential agent for maintaining cell packing density during the elongation process of the mammary duct epithelial tube. NUMB loss caused an aberrant distribution of E-cadherin, leading to cells with lower tension, altered shape, and increased packing, resulting in a reduction in duct elongation [88]. Additionally, NUMB and NUMBL, in cooperation with sarcomeric α-actin, have been found to be essential for regulating Z-disc consolidation in sarcomere assembly and its maintenance in striated muscle [89,90].
On the other hand, EMT is also involved in several pathologies, such as endometriosis, where NUMB down-regulation is correlated with increased cell migration and invasion [91]. A more representative example is its role in tumorigenesis, where NUMB and NOTCH establish a delicate balance that, if disturbed, can lead to aberrant differentiation and cancer progression and metastasis (Figure 2C) [92,93]. Specifically, an increase in the number of asymmetric divisions has been identified as the starting point for the development of cancer stem cells. In more detail, it has been proposed that NUMB isoforms with PRRL may be involved in the early stages of cancer development, promoting proliferation, while NUMB isoforms with PRRS may be involved in the latter stages of cancer, inducing differentiation and loss of cell polarity [94]. Furthermore, NUMB overexpression appears to regulate the malignant transition through the regulation of different pathways. It has been shown to promote EMT through TGFB-dependent ZEB1/Snail2 and MAPK signaling in pancreatic cancer, but it has also been found to inhibit EMT in tongue cancer through RBP-JK-dependent NOTCH1/PTEN/FAK signaling, PAK1/β-catenin signaling in ovarian cancer, and through WNT in colorectal cancer [95,96,97,98,99,100].
NUMB may also play a role in regulating cell adhesion and polarity in response to tyrosine kinase signaling [101]. Interestingly, there is evidence to suggest that in many types of tumors, the EMT transition may be caused by abnormal activity in this pathway [85,101]. When NUMB was knocked down in MDCK cells, it led to the delocalization of the Par3 complex and aPKC, as well as the apical-basal translocation of E-cadherin and β-catenin, polymerization of F-actin, and a decrease in cell-cell adhesion, resulting in an increase in cell proliferation and migration [85,101].
5. NUMB Maintains Cellular Homeostasis by the Regulation of the Endocytic Machinery
NUMB has been suggested to play a role in maintaining cellular equilibrium, specifically in the endocytosis process [102]. This was suggested by the discovery of the interaction of the NPF NUMB motif with Eps15, a component of the endocytic machinery, through its EH domain [2,102,103]. Eps15 is involved in the transport and sorting of molecules [60,104]. However, the deletion of the NPF NUMB motif does not appear to affect NUMB functions, leading to the possibility that its endocytic role may follow an independent proteasome pathway that could play a major role in determining cell fate [105,106,107]. Both Eps15 and the AP-2 adaptor complex are involved in clathrin-mediated endocytosis [105]. Interestingly, Eps15 and the three subunits of the AP-2 adaptor complex appear to interact more strongly with NUMB isoforms containing exon 9 [106]. These interactions allow NUMB to function as a protein involved in the endocytic machinery [2].
Clathrin-dependent endocytosis is a process in which extracellular fluid and proteins are mixed and packaged into clathrin-coated vesicles [68]. NUMB appears to be involved in the localization and co-trafficking of endocytic organelles, as well in the endocytosis of internalized receptors [102]. In Drosophila, NUMB segregation in one of the SOP daughters that results in increased endocytosis and inhibition of NOTCH signaling [61,108,109]. As mentioned above, NUMB is also required for the establishment of NOTCH signaling during cytokinesis [110]. In dividing cells, NUMB delocalizes from the basal cortex of pIIb in a process dependent on the protein α-adaptin [110,111,112]. The interaction between NUMB and the Ear domain of α-adaptin, a subunit of the AP-2 complex, leads to the preferential secretion of α-adaptin in pIIb cells during asymmetric division [111,112]. NUMB appears to act as an adaptor, allowing AP-2 to bind to the NOTCH intracellular domain (NICD) on one side of the pIIb cell. This results in the internalization of the NOTCH receptor and decreased NOTCH activity in pIIb cells [112,113,114]. Through this mechanism, NUMB acts as an inhibitor of the NOTCH pathway through the polarized endocytosis of the NOTCH receptor, while also serving as a cell fate regulator through its binding to NICD [112,115]. Endocytosis is also thought to be critical for the balance between self-renewal and differentiation in NSCs [116]. In this process, NUMB interacts with α-adaptin through the Trunk domain of α-adaptin, regulating the behavior of NSCs through NOTCH. However, this interaction occurs through a different domain of α-adaptin than the one involved in NUMB/α-adaptin interaction in SOPs [116].
In addition, the four-pass transmembrane protein Sanpodo (Spdo) is also involved in NOTCH signaling, which determines NUMB-mediated cell fate [108]. SPDO is internalized during cytokinesis [108] and accumulates with NUMB in pIIb cells, interacting with the NUMB PTB domain through its NPAF (Asn-Pro-Ala-Phe) motif [110,117,118]. In plla cells, SPDO is found on the cortical surface, but in pllb cells, it colocalizes with NOTCH and Delta in RAB5/RAB7-positive endocytic vesicles [119]. NUMB has been proposed to interact with NOTCH-SPDO oligomers in early endosomes, inhibiting NOTCH recycling and allowing for asymmetric distribution of NOTCH on the surface of pIIa/b daughter cells, thereby regulating cell fate determination [110,117]. NUMB also promotes SPDO targeting from endosomes to the plasma membrane, possibly due to NOTCH inhibition [111,118,120]. NUMB is responsible for SPDO removal from the membrane, and its internalization is incompatible with productive NOTCH signaling in pIIb cells. However, in pIIa cells, where NUMB is absent, SPDO remains in the membrane [121].
NUMB has been proposed to act as a regulator of the balance between NOTCH recycling and targeting to late endosomes in neural progenitor cells in Drosophila [122]. Interestingly, NUMB regulates NOTCH trafficking to RAB7-labeled late endosomes, but not to early endosomes [122]. In mammals, NOTCH1 is also constitutively internalized, with differences in its trafficking dynamics depending on changes in NUMB expression [113]. NUMB can also inhibit NOTCH1 activity by regulating post-endocytic sorting events that lead to the degradation of NOTCH1, redirecting the protein to the late endosome compartment [113]. However, NUMB/NUMBL also appears to play a role in sensory axon arborization in neurons by regulating NOTCH1 through the endocytic-lysosomal pathway. In mice, conditional deletion of NUMB in a NUMBL null background resulted in reduced endocytosis and a decrease in axon branch points [83]. It also resulted in a reduction of overall axon length, likely due to the accumulation of NOTCH1 in nuclei [123]. Mammalian NUMB can also antagonize the NOTCH pathway by controlling the post-endocytic trafficking of the NOTCH ligand Delta-like 4 (DLL4) [124]. Low NUMB levels lead to the accumulation of DLL4 on the cell surface, causing aberrant activation of the NOTCH pathway. The NUMB/NUMBL knockdown model showed impairment in the targeting of DLL4 to lysosomes, allowing it to be recycled from the cell surface by RAB11-positive endosomes [124].
NUMB functions as an endocytic regulator of various adhesion molecules, such as integrins and E-cadherin [88]. Clathrin-dependent endocytosis of integrins is a widely recognized process that is essential for cell migration [125]. NUMB binds to β-integrins and colocalizes with them to clathrin-coated structures (CCSs). NUMB phosphorylation by aPKC results in its release from CCSs, preventing it from binding to integrins [126,127]. This inhibition of binding may contribute to cell migration through NUMB’s interaction with Par-3, directing integrin endocytosis to the leading edge [126]. On the other hand, NUMB can also control cadherin-based adhesion through its interaction with p120 catenin [128], a protein that inhibits E-cadherin internalization. Phosphorylation of NUMB by aPKC can prevent its association with p120, attenuating E-cadherin endocytosis and maintaining apicobasal polarity [128]. In addition, up to 25 Ser/Thr phosphorylation sites have been described [129], suggesting a complex regulation of NUMB functions.
As mentioned above, NUMB acts as a cargo-selective endocytic adaptor protein by binding other proteins to the clathrin α-adaptin adaptor [130,131]. This binding is regulated by NUMB phosphorylation, specifically at Ser265 and Ser284, which promotes the recruitment of the 14-3-3 protein. This causes NUMB to dissociate from α-adaptin and translocate from the cortical membrane to the cytosol [130]. NUMB can also be phosphorylated at Ser283, after initial phosphorylation at Ser264, by Ca2+/calmodulin-dependent protein kinase, which abolishes the binding of AP-2 to NUMB and promotes the NUMB-14-3-3 interaction [132,133]. Ca2+/calmodulin-dependent protein kinase may also disrupt the NUMB/AP-2 interaction by phosphorylating NUMB at Ser276 [134]. NUMB phosphorylation by AAK1 (adaptor-associated kinase) appears to be essential for reducing the clathrin coat [135]. Furthermore, CDK5 can phosphorylate NUMB at Ser288, which may play a role in the RAC/RHO axis for controlling cell adhesion and migration [129].
Interestingly, NUMB isoforms appear to regulate the internalization of mGluR5 and mGluR1 differently [136,137,138]. These two metabotropic glutamate receptors are responsible for synaptic development and emotional and motor behaviors in the central nervous system [136,137,138]. The p72 isoform, but not p65, binds to mGluR5 or mGluR1, and increases its expression in the neuronal membrane by inhibiting their endocytosis [136,137]. Additionally, NUMB also regulates the different responses of neural progenitors in an isoform-dependent manner through the regulation of Ca2+ channels [139]. The NUMB PTBL isoforms (p72 and p66) are closely associated with the endocytosis of Ca2+ channels, resulting in their accumulation inside the cell. In contrast, the PTBS isoforms (p71 and p65) preferentially localize the channels to the membrane [139]. NUMB isoforms also appear to have differential role in the NSCs of the brain of Drosophila larvae [36]. PRRL isoforms are expressed during early neurogenesis and promote proliferation, while PRRS isoforms are expressed during neurogenesis and inhibit stem cell proliferation while promoting differentiation. This differential function in different phases of neurogenesis results from endocytic degradation [36].
The presence of enlarged endosomes is an early feature of Alzheimer’s disease and indicates deregulated endocytosis [140]. Furthermore, 70% of the β-amyloid peptide secreted in interstitial fluid is generated through processes related to endocytosis [140,141]. It has been proposed that NUMB may play a role in APP trafficking in an isoform-dependent manner. The PTBL isoforms of NUMB appear to be responsible for targeting APP to late endosome/lysosome, while PTBS isoforms may be responsible for APP accumulation in early endosomes [23]. The C-terminus of the APP family, specifically the YENTPY (Tyr-Glu-Asn-Thr-Pro-Tyr) domain, acts as an anchor for many proteins involved in clathrin-mediated endocytosis and exhibits increased affinity for NUMB PTBL isoforms [23,140,142]. The collapsin response mediator protein 2 (CRMP2) is involved in NUMB-mediated endocytosis, and its increased phosphorylated state is considered an early sign of Alzheimer’s disease [133,143]. CRMP2 colocalizes with NUMB in the central region of axonal growing cones in the neurons of the hippocampus. The NUMB–CRMP2 interaction is mediated by the NUMB-PTB domain, which regulates NUMB-dependent endocytosis in the growth cone [133]. CRMP2 is also involved in NUMB-dependent endocytosis of other proteins, such as the neural L1 cell adhesion molecule (L1CAM), which is endocytosed and recycled in the growth cone, where NUMB and CRMP2 are located [144].
In cancer, NUMB is involved in the non-random segregation of subcellular vesicles [96]. Furthermore, NUMB plays a role in regulating the endocytosis of ALK (anaplastic lymphoma kinase), a receptor that is often aberrantly expressed in cancer [145,146,147]. Interestingly, this regulation appears to be isoform-specific; both NUMB PTBL isoforms can promote receptor endocytosis, but p66 promotes ALK lysosomal degradation through RAB7-containing late endosomes, while p72 allows the kinase to remain active by promoting its recycling back to the plasma membrane [145].
6. NUMB Acts as an Adaptor Protein in the Multiple Signaling Pathways Involved in Morphogenesis Processes and Cancer Development
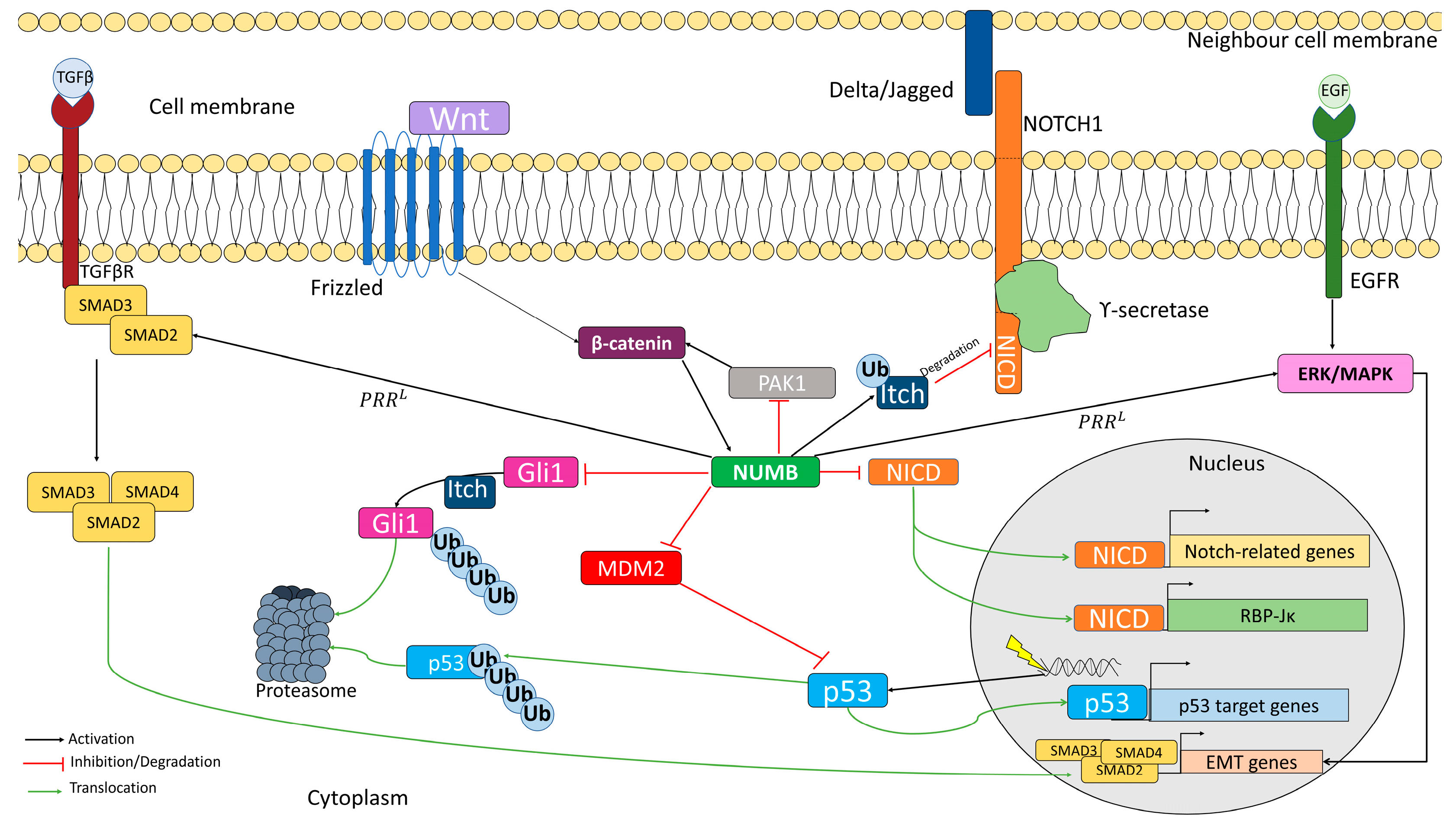
NUMB has been classified as a cargo-selective adaptor and is involved in several important cellular pathways (Figure 3) [24,25,26,27,28,70]. NUMB promotes the ubiquitination of NOTCH on the membrane by recruiting ITCH, which is a ubiquitin E3 ligase that promotes NICD degradation [26,148,149]. This prevents NICD from translocating into the nucleus and the subsequent transcription of NOTCH target genes [26,148,149]. The four NUMB isoforms appear to negatively regulate the transcriptional activity of NOTCH1, but not NOTCH2 or NOTCH3 [148]. Furthermore, NUMB and NOTCH are also inversely expressed during the progression of oligodendrocyte differentiation, with higher NUMB expression in mature oligodendrocytes [150]. In the developing neocortex, NUMB and NOTCH are expressed in the ventricular zone of progenitors, while NUMBL is expressed in postmitotic neurons in the cortical zone [7]. NUMB also plays a role in the proliferation of cardiomyocytes and trabecular morphogenesis through its interaction with NOTCH1 [151]. However, NUMB/NUMBL can inhibit NOTCH2 signaling to control heart myocardial compaction [152]. On the other hand, PRRS isoforms have been suggested to suppress NOTCH signaling in lung cancer cells, while PRRL isoforms increase it [153,154]. There is also a significant inverse correlation between NOTCH1 and NUMB expression in non-small cell lung cancer (NSCLC) [153,154].
In progenitor cells, NUMB has been described as a target of canonical WNT signaling. When activated, the canonical WNT signaling pathway activates the β-catenin cascade, which leads to the induction of NUMB expression. NUMB acts to inhibit NOTCH in progenitors, thereby promoting cell differentiation [24]. The NUMB-mediated WNT-NOTCH network is regulated by androgen receptors [155]. Its disruption has been found to play a role in several types of cancer, such as breast and colorectal cancer [24,97]. NUMB also serves as a key inhibitor of the Hedgehog pathway [156]. Thus, in early brain granule progenitor cells, NUMB overexpression leads to the inhibition of GLI1, which results in the inhibition of stem cell growth and self-renewal, and the promotion of cell differentiation [25,157]. Increased expression of SMO, a key component of the Hedgehog pathway, has been found to increase both the number of stem cells and the spread of chronic myeloid leukemia by decreasing NUMB levels in patients. However, in the SMO KD cell model, NUMB levels are increased, leading to stem cell depletion and slowing the spread of the disease [158]. Decreased NUMB levels have also been found to increase the castration-resistant population in prostate cancer cells as a result of the deregulation of the NOTCH and Hedgehog pathways [159]. It is important to note that NUMB and NUMBL exhibit different behaviors in the regulation of the pathways involved in stem maintenance. While NUMB is an activator of the WNT and Hedgehog pathways, NUMBL acts by inhibiting both pathways. However, both proteins inhibit NOTCH signaling [6].
In parallel, NUMB regulates p53 function by forming a tricomplex with both p53 and the E3 ubiquitin ligase MDM2. This prevents the ubiquitination and subsequent degradation of p53, resulting in elevated levels of p53 [19,160,161,162]. For MDM2 inhibition, the presence of exon 3, present in PTBL isoforms, appears to be required [39,104,163]. Dysregulation of the NUMB/p53/MDM2 complex has been implicated in several tumorigenic events, including the initiation of kidney, breast, or pancreatic cancer [64,164,165]. However, the binding between NUMB and p53 is not only important in cancer, as NUMB also enhances asymmetric mammary stem cell divisions through its interaction with p53 [166].
7. Regulation of NUMB Expression
NUMB expression can be regulated by miRNAs [167,168,169]. In Drosophila, Bantam miRNA controls cell proliferation by inhibiting NUMB in order to bypass cell growth control and regulate the feedback process to maintain the robustness of the NOTCH pathway, which is essential for the fate and self-renewal of NSCs [170,171]. In humans, NUMB mRNA is often targeted by miR-146 family genes, which can affect tissue differentiation and contribute to the development of various diseases [168]. MiR-146a appears to regulate NUMB by influencing the balance between symmetric and asymmetric cell division. In colorectal cancer, it directs symmetric division by suppressing NUMB [172]. This miRNA also plays a role in oral carcinogenesis, promoting cancer cell proliferation and migration by targeting NUMB, IRAK and TRAF6 [173]. However, miR-146a has been shown to have effects beyond tumorigenesis, including skewing the balance between muscle differentiation and cell proliferation by negatively regulating NUMB [174]. Furthermore, miR-146a down-regulation and subsequent NUMB overexpression have also been linked to the suppression of apoptosis and promotion of autophagy in chondrocytes in osteoarthritis and inflammation in the active phase of thyroid-associated ophthalmopathy [175]. MiR-146b has also been shown to promote carcinogenesis in neuroblastoma by targeting NUMB [176].
The miR-31/96/182 families are frequently up-regulated in tumors, such as head and neck squamous cell carcinoma (HNSCC), colorectal or prostate cancer. Therefore, upregulation of these miRNAs in HNSCC increases cell invasiveness and migration by targeting NUMB [167]. Specifically, miR-31, which targets NUMB, has been shown to promote carcinogenesis in colorectal cancer [177]. Furthermore, miR-9-5P, has also been shown to increase stem cell growth and metastasis in prostate cancer by negatively regulating NUMB [178]. Alternative splicing of NUMB isoforms can be indirectly modulated by miR-335, which targets the splicing factor RBM10. In tumors, overexpression of miR-335 has been linked to increased tumor growth and decreased expression of RBM10, as well as increased expression of long NUMB isoforms (p72/71) [179].
NUMB also appears to be involved in other pathologies, such as preeclampsia, a pregnancy disorder associated with an increased risk of neonatal, fetal, or maternal morbidity/mortality. Increased levels of miRNA-524-5p in this pathology are involved in the regulation of trophoblast proliferation and invasiveness by targeting NUMB and subsequent regulation of the NOTCH pathway [180].
8. NUMB as a Therapeutic Tool for Various Pathologies
Recent studies have suggested NUMB as a possible biomarker for prognosis and/or response to certain therapies for cancer, Alzheimer’s disease, and other pathologies, with potential clinical applications [21,140,181,182,183,184].
Interestingly, there is controversy regarding the role of NUMB as a tumor suppressor, as oncogenic behavior has been observed in some cases [185]. NUMB downregulation has been linked to poor prognosis in various types of carcinomas, melanoma, and glioblastoma, among others [95,97,98,159,164,183,185,186,187,188,189,190,191,192]. On the other hand, NUMB up-regulation has been associated with poor prognosis in certain tumors, such as hepatocellular carcinoma (HCC) [193] and esophageal squamous cell carcinoma (ESCC), where NUMB overexpression has been linked to increased tumor recurrence and poor overall survival [194], but more specifically, the possible functional diversity of the isoforms has been investigated. In these carcinomas, p72/71 is often found to be downregulated. In both HCC and ESCC, the expression of p72/p71 NUMB isoforms has been associated with increased early recurrence and lower overall survival after surgery due to increased proliferation, migration, and invasion in cancer cells. On the other hand, higher p66/p65 expression levels promote the opposite effects [193,194,195,196]. Therefore, in these carcinomas, a difference in prognostic meaning can be made between the isoforms [193,194,195,196].
However, the role of NUMB appears to be tissue-dependent and the prognostic significance of the isoforms cannot be extrapolated to all tumors. For example, in medulloblastoma the opposite effect occurs, with p72/71 being upregulated and p66/65 downregulated [38].
In addition, NUMB has become a predictive biomarker for patients for whom therapy would be appropriate, for example, in patients with prostate cancer who could benefit from NOTCH inhibition or therapies that restore p53 function, such as the Nutlin-related class of anti-MDM2 inhibitors, which are already used in breast cancer [41,189].
NUMB overexpression has been associated with increased sensitivity to cisplatin treatment in patients with epithelioid malignant pleural mesothelioma, but with poor response to treatment in ESCC, suggesting a role for NUMB in resistance to therapy [194,197]. Thus, NUMB downregulation has been associated with increased castration-resistant progenitors in prostate cancer, resistance to imatinib in chronic myeloid leukemia, and increased radioresistance in pancreatic cancer. This last effect was reversed by NUMB upregulation through metformin treatment [159,198]. Inhibition of NOTCH/NUMB signaling has also been associated with increased radiation sensitivity in nasopharyngeal carcinoma [199]. In contrast, NUMB overexpression in NSCLC has been associated with increased sensitivity to radiation [200].
Regarding Alzheimer’s disease, pharmacological modulation of APP by downregulating NUMB has been proposed as a novel therapeutic strategy. The effect reduced the cleavage of APP by γ-secretase, subsequently reducing β-amyloid peptide levels [201]. Furthermore, the proposal of NUMB as a possible therapeutic target for Alzheimer’s disease has been taken even further, with the possibility that the therapeutic value of the isoforms could be differentiated. The switch from p72/66 to p71/65 isoforms is an essential step in increasing the accumulation of β-amyloid peptide plaques in this disease [202]. Therefore, NUMB could represent a potential therapeutic target for decreasing the accumulation of these peptides [202]. In addition, NUMB appears to be involved in an isoform-specific manner in the regulation of Tau protein levels, which is also implicated in this pathology. Only the overexpression of p72 is able to decrease intracellular Tau levels, enhancing neuronal electrical activity. Consequently, this isoform could be considered as an important therapeutic factor [46].
Finally, NUMB has also been proposed as a possible biomarker or therapeutic target for other pathologies. In the renal field, it seems to be related to diabetic nephropathy, kidney fibrosis, acute kidney injury, and proteinuric diseases [43,182,203,204,205]. In the case of renal fibrosis, NUMB appears to be overexpressed in affected kidneys compared to healthy kidneys, making it a potential biomarker for the disease [182,206]. Conversely, in acute kidney disease, NUMB promotes the activation of p53-mediated protective autophagy, making it a potential therapeutic target for this disease [43,205,207]. Similarly, NUMB has emerged as a promising therapeutic target for proteinuric pathologies due to its protective role against endoplasmic reticulum stress-associated apoptosis in these diseases [203].
Furthermore, NUMB inhibition has also been proposed as a possible effective therapeutic strategy for another type of fibrosis, lung fibrosis. In this disease, NUMB prevents the activation of β-catenin signaling through its interaction with casein kinase 2 [181].
In terms of cardiovascular pathologies, NUMB has been proposed as an indicator of an increased risk of coronary artery disease. In this case, lower NUMB expression indicates a higher risk of developing the disease [208]. In addition, NUMB has also been proposed as a therapeutic candidate for muscle and cardiac regeneration, as well as for congenital heart disease [209,210]. Therefore, NUMB downregulation has been classified as a potential strategy to inhibit ischemia-induced apoptosis [211].
9. Conclusions
The multitasking role of NUMB demonstrates its importance in maintaining cellular homeostasis. NUMB has been characterized as an essential protein for cell polarization and asymmetric division as well as for other processes, such as endocytosis and the regulation of multiple cellular pathways. Dysregulation of its function could be a cause of pathologies such as Alzheimer’s disease or cancer, further emphasizing its importance in the cell. However, much is still unknown about NUMB and its isoforms, which increases its potential for a variety of functions. Further study of NUMB and its isoforms could increase our knowledge of cellular mechanisms and the molecular causes of different pathologies, potentially leading to advances in personalized medicine.
Author Contributions
S.M.O.-C. and J.M.G.-H. conceived and designed this work and wrote and edited the manuscript. All authors revised the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by different grants: Grants RTI2018-097455-B-I00 and PID2021-122629OB-I00 funded by MCIN/AEI/10.13039/501100011033 and by “ERDF A way of making Europe”, by the “European Union”. Additional grants from CIBER de Cáncer (CB16/12/00275), from Consejeria de Salud (PI-0397-2017) and Project P18-RT-2501 from 2018 competitive research projects call within the scope of PAIDI 2020—80% co-financed by the European Regional Development Fund (ERDF) from the Regional Ministry of Economic Transformation, Industry, Knowledge and Universities. Junta de Andalucía.Special thanks to the AECC (Spanish Association of Cancer Research) Founding Ref. GC16173720CARR for supporting this work.
Acknowledgments
Special thanks to the Fundación AECC.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Yan, B. Numb—From Flies to Humans. Brain Dev. 2010, 32, 293–298. [Google Scholar] [CrossRef] [PubMed]
- Gulino, A.; di Marcotullio, L.; Screpanti, I. The Multiple Functions of Numb. Exp. Cell Res. 2010, 316, 900–906. [Google Scholar] [CrossRef] [PubMed]
- Knoblich, J.A.; Jan, L.Y.; Nung Jan, Y. The N terminus of the Drosophila Numb protein directs membrane association and actin-dependent asymmetric localization. Dev. Biol. 1997, 94, 13005–13010. [Google Scholar] [CrossRef] [Green Version]
- Knoblich, J.A.; Jan, L.Y.; Nung Jan, Y. Asymmetric Segregation of Numb and Prospero during Cell Division. Nature 1995, 377, 624–627. [Google Scholar] [CrossRef]
- Rhyu, M.S.; Yeh Jan, L.; Nung Jan, Y. Asymmetric Distribution of Numb Protein during Division of the Sensory Organ Precursor Cell Confers Distinct Fates to Daughter Cells. Cell 1994, 76, 477–491. [Google Scholar] [CrossRef]
- García-Heredia, J.M.; Carnero, A. NUMB and NUMBL Differences in Gene Regulation. Oncotarget 2018, 9, 9219–9234. [Google Scholar] [CrossRef] [Green Version]
- Zhong, W.; Jiang, M.-M.; Weinmaster, G.; Yeh Jan, L.; Nung Jan, Y. Differential Expression of Mammalian Numb, Numblike and Notch1 Suggests Distinct Roles during Mouse Cortical Neurogenesis. Development 1997, 124, 1887–1897. [Google Scholar] [CrossRef] [PubMed]
- Uemura, T.; Shepherd, S.; Ackerman, L.; Jan, L.Y.; Jan, Y.N. Numb, a Gene Required in Determination of Cell Fate during Sensory Organ Formation in Drosophila Embryos. Cell 1989, 58, 349–360. [Google Scholar] [CrossRef]
- Cayouette, M.; Raff, M. Asymmetric Segregation of Numb: A Mechanism for Neural Specification from Drosophila to Mammals. Nat. Neurosci. 2002, 5, 1265–1269. [Google Scholar] [CrossRef] [PubMed]
- Cicalese, A.; Bonizzi, G.; Pasi, C.E.; Faretta, M.; Ronzoni, S.; Giulini, B.; Brisken, C.; Minucci, S.; di Fiore, P.P.; Pelicci, P.G. The Tumor Suppressor P53 Regulates Polarity of Self-Renewing Divisions in Mammary Stem Cells. Cell 2009, 138, 1083–1095. [Google Scholar] [CrossRef]
- El-Hashash, A.H.K.; Warburton, D. Numb Expression and Asymmetric versus Symmetric Cell Division in Distal Embryonic Lung Epithelium. J. Histochem. Cytochem. 2012, 60, 675–682. [Google Scholar] [CrossRef] [Green Version]
- Wakamatsu, Y.; Maynard, T.M.; Jones, S.U.; Weston, J.A. NUMB Localizes in the Basal Cortex of Mitotic Avian Neuroepithelial Cells and Modulates Neuronal Differentiation by Binding to NOTCH-1. Neuron 1999, 23, 71–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wakamatsu, Y.; Maynard, T.M.; Weston, J.A. Fate Determination of Neural Crest Cells by NOTCH-Mediated Lateral Inhibition and Asymmetrical Cell Division during Gangliogenesis. Development 2000, 127, 2811–2821. [Google Scholar] [CrossRef] [PubMed]
- Venters, S.J.; Ordahl, C.P. Asymmetric Cell Divisions Are Concentrated in the Dermomyotome Dorsomedial Lip during Epaxial Primary Myotome Morphogenesis. Anat. Embryol. 2005, 209, 449–460. [Google Scholar] [CrossRef]
- Holowacz, T.; Zeng, L.; Lassar, A.B. Asymmetric Localization of Numb in the Chick Somite and the Influence of Myogenic Signals. Dev. Dyn. 2006, 235, 633–645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tosoni, D.; Zecchini, S.; Coazzoli, M.; Colaluca, I.; Mazzarol, G.; Rubio, A.; Caccia, M.; Villa, E.; Zilian, O.; di Fiore, P.P.; et al. The Numb/P53 Circuitry Couples Replicative Self-Renewal and Tumor Suppression in Mammary Epithelial Cells. J. Cell Biol. 2015, 211, 845–862. [Google Scholar] [CrossRef] [Green Version]
- Yan, B.; Omar, F.M.; Das, K.; Ng, W.H.; Lim, C.; Shiuan, K.; Yap, C.T.; Salto-Tellez, M. Characterization of Numb Expression in Astrocytomas. Neuropathology 2008, 28, 479–484. [Google Scholar] [CrossRef]
- Rennstam, K.; McMichael, N.; Berglund, P.; Honeth, G.; Hegardt, C.; Rydén, L.; Luts, L.; Bendahl, P.-O.; Hedenfalk, I. Numb Protein Expression Correlates with a Basal-like Phenotype and Cancer Stem Cell Markers in Primary Breast Cancer. Breast Cancer Res. Treat. 2010, 122, 315–324. [Google Scholar] [CrossRef] [Green Version]
- Colaluca, I.N.; Tosoni, D.; Nuciforo, P.; Senic-Matuglia, F.; Galimberti, V.; Viale, G.; Pece, S.; di Fiore, P.P. NUMB Controls P53 Tumour Suppressor Activity. Nature 2008, 451, 76–80. [Google Scholar] [CrossRef]
- Sherrington, R.; Rogaev, E.I.; Liang, Y.; Rogaeva, E.A.; Levesque, G.; Ikeda, M.; Chi, H.; Lin, C.; Li, G.; Holman, K.; et al. Cloning of a Gene Bearing Missense Mutations in Early-Onset Familial Alzheimer’s Disease. Nature 1995, 375, 754–760. [Google Scholar] [CrossRef]
- Kyriazis, G.A.; Wei, Z.; Vandermey, M.; Jo, D.G.; Xin, O.; Mattson, M.P.; Chan, S.L. Numb Endocytic Adapter Proteins Regulate the Transport and Processing of the Amyloid Precursor Protein in an Isoform-Dependent Manner: Implications for Alzheimer Disease Pathogenesis. J. Biol. Chem. 2008, 283, 25492–25502. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Zhou, T.; Sun, Z.; Ye, T.; Zhou, S.; Li, J.; Liu, Y.; Kong, L.; Tang, J.; Liu, D.; et al. Zeb1 Regulates the Symmetric Division of Mouse Lewis Lung Carcinoma Stem Cells through Numb Mediated by MiR-31. Int. J. Biol. Sci. 2018, 14, 1399–1410. [Google Scholar] [CrossRef]
- Sun, M.; Asghar, S.Z.; Zhang, H. The Polarity Protein Par3 Regulates APP Trafficking and Processing through the Endocytic Adaptor Protein Numb. Neurobiol. Dis. 2016, 93, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Katoh, M.; Katoh, M. NUMB Is a Break of WNT—Notch Signaling Cycle. Int. J. Mol. Med. 2006, 18, 517–521. [Google Scholar] [CrossRef] [Green Version]
- di Marcotullio, L.; Ferretti, E.; Greco, A.; de Smaele, E.; Po, A.; Sico, M.A.; Alimandi, M.; Giannini, G.; Maroder, M.; Screpanti, I.; et al. Numb Is a Suppressor of Hedgehog Signalling and Targets Gli1 for Itch-Dependent Ubiquitination. Nat. Cell Biol. 2006, 8, 1415–1423. [Google Scholar] [CrossRef] [PubMed]
- McGill, M.A.; McGlade, C.J. Mammalian Numb Proteins Promote Notch1 Receptor Ubiquitination and Degradation of the Notch1 Intracellular Domain. J. Biol. Chem. 2003, 278, 23196–23203. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.R.; Sun, J.; Wang, J.; Lu, Y.Y. Advances in research on cell fate determinant Numb regulating liver cancer. Zhonghua Gan Zang Bing Za Zhi 2018, 26, 714–717. [Google Scholar] [PubMed]
- Garcia-Heredia, J.M.; Lucena-Cacace, A.; Verdugo-Sivianes, E.M.; Perez, M.; Carnero, A. The Cargo Protein MAP17 (PDZK1IP1) Regulates the Cancer Stem Cell Pool Activating the Notch Pathway by Abducting NUMB. Clin. Cancer Res. 2017, 23, 3871–3883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filippone, M.G.; Freddi, S.; Zecchini, S.; Restelli, S.; Colaluca, I.; Bertalot, G.; Pece, S.; Tosoni, D.; di Fiore, P.P. Aberrant Phosphorylation Inactivates Numb in Breast Cancer Causing Expansion of the Stem Cell Pool. J. Cell Biol. 2022, 221, e202112001. [Google Scholar] [CrossRef]
- Pece, S.; Serresi, M.; Santolini, E.; Capra, M.; Hulleman, E.; Galimberti, V.; Zurrida, S.; Maisonneuve, P.; Viale, G.; di Fiore, P.P. Loss of Negative Regulation by Numb over Notch Is Relevant to Human Breast Carcinogenesis. J. Cell Biol. 2004, 167, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Thol, F.; Winschel, C.; Sonntag, A.-K.; Damm, F.; Wagner, K.; Chaturvedi, A.; Göhring, G.; Schlegelberger, B.; Lübbert, M.; Fiedler, W.; et al. Prognostic Significance of Expression Levels of Stem Cell Regulators MSI2 and NUMB in Acute Myeloid Leukemia. Ann. Hematol. 2013, 92, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Westhoff, B.; Colaluca, I.N.; D’Ario, G.; Donzelli, M.; Tosoni, D.; Volorio, S.; Pelosi, G.; Spaggiari, L.; Mazzarol, G.; Viale, G.; et al. Alterations of the Notch Pathway in Lung Cancer. Proc. Natl. Acad. Sci. USA 2009, 106, 22293–22298. [Google Scholar] [CrossRef] [Green Version]
- Maiorano, E.; Favia, G.; Pece, S.; Resta, L.; Maisonneuve, P.; di Fiore, P.P.; Capodiferro, S.; Urbani, U.; Viale, G. Prognostic Implications of Numb Immunoreactivity in Salivary Gland Carcinomas. Int. J. Immunopathol. Pharmacol. 2007, 20, 779–789. [Google Scholar] [CrossRef] [Green Version]
- Choi, H.Y.; Seok, J.; Kang, G.H.; Lim, K.M.; Cho, S.G. nbib The Role of NUMB/NUMB Isoforms in Cancer Stem Cells. BMB Rep. 2021, 54, 335–343. [Google Scholar] [CrossRef]
- Dho, S.E.; French, M.B.; Woods, S.A.; McGlade, C.J. Characterization of Four Mammalian Numb Protein Isoforms. Identification of Cytoplasmic and Membrane-Associated Variants of the Phosphotyrosine Binding Domain. J. Biol. Chem. 1999, 274, 33097–33104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toriya, M.; Tokunaga, A.; Sawamoto, K.; Nakao, K.; Okano, H. Distinct Functions of Human Numb Isoforms Revealed by Misexpression in the Neural Stem Cell Lineage in the Drosophila Larval Brain. Dev. Neurosci. 2006, 28, 142–155. [Google Scholar] [CrossRef]
- Zhang, Y.; Dho, S.E.; Othman, K.; Simpson, C.D.; Lapierre, J.; Bondoc, A.; McGlade, C.J. Numb Exon 9 Inclusion Regulates Integrinβ5 Surface Expression and Promotes Breast Cancer Metastasis. Oncogene 2022, 41, 2079–2094. [Google Scholar] [CrossRef]
- Abballe, L.; Mastronuzzi, A.; Miele, E.; Carai, A.; Besharat, Z.M.; Moretti, M.; de Smaele, E.; Giangaspero, F.; Locatelli, F.; Ferretti, E.; et al. Numb Isoforms Deregulation in Medulloblastoma and Role of P66 Isoform in Cancer and Neural Stem Cells. Front. Pediatr. 2018, 6, 315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colaluca, I.N.; Basile, A.; Freiburger, L.; D’Uva, V.; Disalvatore, D.; Vecchi, M.; Confalonieri, S.; Tosoni, D.; Cecatiello, V.; Malabarba, M.G.; et al. A Numb-Mdm2 Fuzzy Complex Reveals an Isoformspecific Involvement of Numb in Breast Cancer. J. Cell Biol. 2018, 217, 745–762. [Google Scholar] [CrossRef] [Green Version]
- Karaczyn, A.; Bani-Yaghoub, M.; Tremblay, R.; Kubu, C.; Cowling, R.; Adams, T.L.; Prudovsky, I.; Spicer, D.; Friesel, R.; Vary, C.; et al. Two Novel Human NUMB Isoforms Provide a Potential Link between Development and Cancer. Neural Dev. 2010, 5, 31. [Google Scholar] [CrossRef]
- Tosoni, D.; Pambianco, S.; Ekalle Soppo, B.; Zecchini, S.; Bertalot, G.; Pruneri, G.; Viale, G.; di Fiore, P.P.; Pece, S. Pre-Clinical Validation of a Selective Anti-Cancer Stem Cell Therapy for Numb-Deficient Human Breast Cancers. EMBO Mol. Med. 2017, 9, 655–671. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Qi, S.; Fu, Y.; Shen, L.; Li, M.; Lu, J.; Zhao, X.; Zhang, H. NUMB Knockdown Enhanced the Anti-Tumor Role of Cisplatin on Ovarian Cancer Cells by Inhibiting Cell Proliferation and Epithelial-Mesenchymal Transition. Transl. Cancer Res. 2019, 8, 379–388. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Li, Z.; Chen, Z.; Li, C.; Lei, L.; Wu, X.; Li, Y. Numb Ameliorates Necrosis and Inflammation in Acute Kidney Injury Induced by Cisplatin. Chem. Biol. Interact. 2020, 330, 109251. [Google Scholar] [CrossRef]
- Zhu, D.; Xia, J.; Liu, C.; Fang, C. Numb/Notch/PLK1 Signaling Pathway Mediated Hyperglycemic Memory in Pancreatic Cancer Cell Radioresistance and the Therapeutic Effects of Metformin. Cell. Signal. 2022, 93, 110268. [Google Scholar] [CrossRef]
- Yang, Y.; Li, L.; He, H.; Shi, M.; He, L.; Liang, S.; Qi, J.; Chen, W. Numb Inhibits Migration and Promotes Proliferation of Colon Cancer Cells via RhoA/ROCK Signaling Pathway Repression. Exp. Cell Res. 2022, 411, 113004. [Google Scholar] [CrossRef] [PubMed]
- Lacomme, M.; Hales, S.C.; Brown, T.W.; Stevanovic, K.; Jolicoeur, C.; Cai, J.; Bois, T.; Desrosiers, M.; Dalkara, D.; Cayouette, M. Numb Regulates Tau Levels and Prevents Neurodegeneration in Tauopathy Mouse Models. Sci. Adv. 2022, 8, eabm4295. [Google Scholar] [CrossRef]
- Stelzer, G.; Rosen, N.; Plaschkes, I.; Zimmerman, S.; Twik, M.; Fishilevich, S.; Stein, T.I.; Nudel, R.; Lieder, I.; Mazor, Y.; et al. The GeneCards Suite: From Gene Data Mining to Disease Genome Sequence Analyses. Curr. Protoc. Bioinform. 2016, 54, 1.30.1–1.30.33. [Google Scholar] [CrossRef]
- Database Resources of the National Center for Biotechnology Information. Nucleic Acids Res. 2016, 44, D7–D19. [CrossRef] [PubMed] [Green Version]
- Hamosh, A.; Scott, A.F.; Amberger, J.S.; Bocchini, C.A.; McKusick, V.A. Online Mendelian Inheritance in Man (OMIM), a Knowledgebase of Human Genes and Genetic Disorders. Nucleic Acids Res. 2005, 33, D514–D517. [Google Scholar] [CrossRef]
- 2021 Alzheimer’s Disease Facts and Figures. Alzheimer’s Dement. 2021, 17, 327–406. [CrossRef]
- Zhang, Y.; Thompson, R.; Zhang, H.; Xu, H. APP Processing in Alzheimer ’s Disease. Mol. Brain 2011, 4, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lane, C.A.; Hardy, J.; Schott, J.M. Alzheimer’s Disease. Eur. J. Neurol. 2018, 25, 59–70. [Google Scholar] [CrossRef]
- Kent, W.J.; Sugnet, C.W.; Furey, T.S.; Roskin, K.M.; Pringle, T.H.; Zahler, A.M.; Haussler, D. The Human Genome Browser at UCSC. Genome Res. 2002, 12, 996–1006. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Feng, X.; Li, S.C. SCDevDB: A Database for Insights into Single-Cell Gene Expression Profiles during Human Developmental Processes. Front. Genet. 2019, 10, 903. [Google Scholar] [CrossRef]
- Hornbeck, P.V.; Zhang, B.; Murray, B.; Kornhauser, J.M.; Latham, V.; Skrzypek, E. PhosphoSitePlus, 2014: Mutations, PTMs and Recalibrations. Nucleic Acids Res. 2015, 43, D512–D520. [Google Scholar] [CrossRef] [Green Version]
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The CBio Cancer Genomics Portal: An Open Platform for Exploring Multidimensional Cancer Genomics Data. Cancer Discov. 2012, 2, 401–404. [Google Scholar] [CrossRef] [Green Version]
- Díez-Villanueva, A.; Mallona, I.; Peinado, M.A. Wanderer, an Interactive Viewer to Explore DNA Methylation and Gene Expression Data in Human Cancer. Epigenet. Chromatin 2015, 8, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Consortium, T.U. UniProt: A Hub for Protein Information. Nucleic Acids Res. 2014, 43, D204–D212. [Google Scholar] [CrossRef] [Green Version]
- Rajendran, D.; Zhang, Y.; Berry, D.M.; McGlade, C.J. Regulation of Numb Isoform Expression by Activated ERK Signaling. Oncogene 2016, 35, 5202–5213. [Google Scholar] [CrossRef] [PubMed]
- Salcini, A.E.; Confalonieri, S.; Doria, M.; Santolini, E.; Tassi, E.; Minenkova, O.; Cesareni, G.; Pelicci, P.G.; Paolo, P.; Fiore, D. Binding Specificity and in Vivo Targets of the EH Domain, a Novel Protein-Protein Interaction Module. Genes Dev. 1997, 11, 2239–2249. [Google Scholar] [CrossRef] [PubMed]
- Giebel, B.; Wordarz, A. Notch Signaling: Numb Makes the Difference. Curr. Biol. 2012, 22, R133–R135. [Google Scholar] [CrossRef] [Green Version]
- Verdi, J.M.; Bashirullah, A.; Goldhawk, D.E.; Kubu, C.J.; Jamali, M.; Meakin, S.O.; Lipshitz, H.D. Distinct Human NUMB Isoforms Regulate Differentiation vs. Proliferation in the Neuronal Lineage. Proc. Natl. Acad. Sci. USA 1999, 96, 10472–10476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedersen, W.A.; Chan, S.L.; Zhu, H.; Abdur-Rahman, L.A.; Verdi, J.M.; Mattson, M.P. Numb Isoforms Containing a Short PTB Domain Promote Neurotrophic Factor-Induced Differentiation and Neurotrophic Factor Withdrawal-Induced Death of PC12 Cells. J. Neurochem. 2002, 82, 976–986. [Google Scholar] [CrossRef] [PubMed]
- Siddique, H.R.; Feldman, D.E.; Chen, C.L.; Punj, V.; Tokumitsu, H.; Machida, K. NUMB Phosphorylation Destabilizes P53 and Promotes Self-Renewal of Tumor-Initiating Cells by a NANOG-Dependent Mechanism in Liver Cancer. Hepatology 2015, 62, 1466–1479. [Google Scholar] [CrossRef] [Green Version]
- Yogosawa, S.; Miyauchi, Y.; Honda, R.; Tanaka, H.; Yasuda, H. Mammalian Numb Is a Target Protein of Mdm2, Ubiquitin Ligase. Biochem. Biophys. Res. Commun. 2003, 302, 869–872. [Google Scholar] [CrossRef]
- Sczaniecka, M.; Gladstone, K.; Pettersson, S.; McLaren, L.; Huart, A.S.; Wallace, M. MDM2 Protein-Mediated Ubiquitination of NUMB Protein: Identification of a Second Physiological Substrate of MDM2 That Employs a Dual-Site Docking Mechanism. J. Biol. Chem. 2012, 287, 14052–14068. [Google Scholar] [CrossRef] [Green Version]
- Dhami, G.K.; Liu, H.; Galka, M.; Voss, C.; Wei, R.; Muranko, K.; Kaneko, T.; Cregan, S.P.; Li, L.; Li, S.S.C. Dynamic Methylation of Numb by Set8 Regulates Its Binding to P53 and Apoptosis. Mol. Cell 2013, 50, 565–576. [Google Scholar] [CrossRef] [Green Version]
- Shen, Q.; Temple, S. Creating Asymmetric Cell Divisions by Skewing Endocytosis. Sci. Signal. 2002, 162, pe52. [Google Scholar] [CrossRef]
- Cayouette, M.; Whitmore, A.V.; Jeffery, G.; Raff, M. Asymmetric Segregation of Numb in Retinal Development and the Influence of the Pigmented Epithelium. J. Neurosci. 2001, 21, 5643–5651. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Yeh Jan, L.; Nung Jan, Y. Control of Daughter Cell Fates during Asymmetric Division: Interaction of Numb and Notch. Neuron 1996, 17, 27–41. [Google Scholar] [CrossRef]
- Lu, B.; Rothenberg, M.; Jan, L.Y.; Nung Jan, Y. Partner of Numb Colocalizes with Numb during Mitosis and Directs Numb Asymmetric Localization in Drosophila Neural and Muscle Progenitors. Cell 1998, 95, 225–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.; Walsh, C.A. Numb, neurogenesis and epithelial polarity. Nat. Neurosci. 2007, 10, 812–813. [Google Scholar] [CrossRef]
- Roegiers, F.; Jan, Y.N. Asymmetric Cell Division. Curr. Opin. Cell Biol. 2004, 16, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Justice, N.; Roegiers, F.; Yeh Jan, L.; Nung Jan, Y. Lethal Giant Larvae Acts Together with Numb in Notch Inhibition and Cell Fate Specification in the Drosophila Adult Sensory Organ Precursor Lineage. Curr. Biol. 2003, 13, 778–783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, C.A.; Lau, K.M.; Rahmani, Z.; Dho, S.E.; Brothers, G.; She, Y.M.; Berry, D.M.; Bonneil, E.; Thibault, P.; Schweisguth, F.; et al. APKC-Mediated Phosphorylation Regulates Asymmetric Membrane Localization of the Cell Fate Determinant Numb. EMBO J. 2007, 26, 468–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kraut, R.; Chia, W.; Jan, L.Y.; Jan, Y.N.; Knoblich, J.A. Role of Inscuteable in Orienting Asymmetric Cell Divisions in Drosophila. Nature 1996, 383, 50–55. [Google Scholar] [CrossRef]
- Knoblich, J.A. Mechanisms of Asymmetric Cell Division during Animal Development. Curr. Opin. Cell Biol. 1997, 9, 833–841. [Google Scholar] [CrossRef] [PubMed]
- Schober, M.; Schaefer, M.; Knoblich, J.A. Bazooka Recruits Inscuteable to Orient Asymmetric Cell Divisions in Drosophila Neuroblasts. Nature 1999, 402, 548–551. [Google Scholar] [CrossRef] [PubMed]
- Charnley, M.; Ludford-Menting, M.; Pham, K.; Russell, S.M.; Lennon-Duménil, A.M. A new role for Notch in the control of polarity and asymmetric cell division of developing T cells. J. Cell Sci. 2020, 133, jcs235358. [Google Scholar] [CrossRef] [Green Version]
- Qin, H.; Percival-Smith, A.; Li, C.; Jia, C.Y.H.; Gloor, G.; Li, S.S.C. A Novel Transmembrane Protein Recruits Numb to the Plasma Membrane during Asymmetric Cell Division. J. Biol. Chem. 2004, 279, 11304–11312. [Google Scholar] [CrossRef]
- Domingos, P.M.; Jenny, A.; Combie, K.F.; del Alamo, D.; Mlodzik, M.; Steller, H.; Mollereau, B. Regulation of Numb during Planar Cell Polarity Establishment in the Drosophila Eye. Mech. Dev. 2019, 160, 103583. [Google Scholar] [CrossRef]
- Li, H.-S.; Wang, D.; Shen, Q.; Schonemann, M.D.; Gorski, J.A.; Jones, K.R.; Temple, S.; Yeh Jan, L.; Nung Jan, Y. Inactivation of Numb and Numblike in Embryonic Dorsal Forebrain Impairs Neurogenesis and Disrupts Cortical. Neuron 2003, 40, 1105–1118. [Google Scholar] [CrossRef] [Green Version]
- Rašin, M.R.; Gazula, V.R.; Breunig, J.J.; Kwan, K.Y.; Johnson, M.B.; Liu-Chen, S.; Li, H.S.; Jan, L.Y.; Jan, Y.N.; Rakic, P.; et al. Numb and Numbl Are Required for Maintenance of Cadherin-Based Adhesion and Polarity of Neural Progenitors. Nat. Neurosci. 2007, 10, 819–827. [Google Scholar] [CrossRef] [PubMed]
- Petersen, P.H.; Zou, K.; Hwang, J.K.; Jan, Y.N.; Zhong, W. Progenitor Cell Maintenance Requires Numb and Numblike during Mouse Neurogenesis. Nature 2002, 419, 929–934. [Google Scholar] [CrossRef]
- Wang, Z.; Li, S.S. Numb: A New Player in EMT. Cell Adhes. Migr. 2010, 4, 176–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, V.; Bhattacharya, S.; Chikkaputtaiah, C.; Hazra, S.; Pal, M. The Basics of Epithelial–Mesenchymal Transition (EMT): A Study from a Structure, Dynamics, and Functional Perspective. J. Cell Physiol. 2019, 234, 14535–14555. [Google Scholar] [CrossRef] [PubMed]
- Miao, L.; Li, J.; Li, J.; Lu, Y.; Shieh, D.; Mazurkiewicz, J.E.; Barroso, M.; Schwarz, J.J.; Xin, H.B.; Singer, H.A.; et al. Cardiomyocyte Orientation Modulated by the Numb Family Proteins-N-Cadherin Axis Is Essential for Ventricular Wall Morphogenesis. Proc. Natl. Acad. Sci. USA 2019, 116, 15560–15569. [Google Scholar] [CrossRef] [Green Version]
- Chatterjee, S.J.; Halaoui, R.; Deagle, R.C.; Rejon, C.; McCaffrey, L. Numb Regulates Cell Tension Required for Mammary Duct Elongation. Biol. Open 2019, 8, bio042341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, B.; Yang, M.; Li, S. Numb and Numblike Regulate Sarcomere Assembly and Maintenance. J. Clin. Investig. 2022, 132, e139420. [Google Scholar] [CrossRef]
- de Gasperi, R.; Mo, C.; Azulai, D.; Wang, Z.; Harlow, L.M.; Du, Y.; Graham, Z.; Pan, J.; Liu, X.; Guo, L.; et al. Numb Is Required for Optimal Contraction of Skeletal Muscle. J. Cachexia Sarcopenia Muscle 2022, 13, 454–466. [Google Scholar] [CrossRef]
- Schüring, A.N.; Dahlhues, B.; Korte, A.; Kiesel, L.; Titze, U.; Heitkötter, B.; Ruckert, C.; Götte, M. The Endometrial Stem Cell Markers Notch-1 and Numb Are Associated with Endometriosis. Reprod. Biomed. Online 2018, 36, 294–301. [Google Scholar] [CrossRef]
- Ostrakhovitch, E.A. Interplay between Numb and Notch in Epithelial Cancers: Role for Dual Oxidase Maturation Factor. Eur. J. Cancer 2009, 45, 2071–2076. [Google Scholar] [CrossRef]
- Hu, X.B.; Ouyang, L.Z.; He, Y.; Xia, M.Z. Numb Confers to Inhibit Epithelial Mesenchymal Transition via β-Catenin/Lin28 Signaling Pathway in Breast Cancer. Exp. Mol. Pathol. 2019, 109, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Chen, X.; Ye, F.; Lu, W.; Xie, X. Symmetric Division and Expression of Its Regulatory Gene Numb in Human Cervical Squamous Carcinoma Cells. Pathobiology 2009, 76, 149–154. [Google Scholar] [CrossRef] [PubMed]
- Li, J.Y.; Huang, W.X.; Zhou, X.; Chen, J.; Li, Z. Numb Inhibits Epithelial-Mesenchymal Transition via RBP-Jκ-Dependent Notch1/PTEN/FAK Signaling Pathway in Tongue Cancer. BMC Cancer 2019, 19, 391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, W.L.; Yang, M.H. Numb Is Involved in the Non-Random Segregation of Subcellular Vesicles in Colorectal Cancer Stem Cells. Cell Cycle 2016, 15, 2697–2703. [Google Scholar] [CrossRef] [Green Version]
- Cheng, C.; Huang, Z.; Zhou, R.; An, H.; Cao, G.; Ye, J.; Huang, C.; Daoyi Wu, X. Numb Negatively Regulates the Epithelial-to-Mesenchymal Transition in Colorectal Cancer through the Wnt Signaling Pathway. Am. J. Physiol. Gastrointest. Liver Physiol. 2020, 318, G841–G852. [Google Scholar] [CrossRef]
- Liang, J.; Han, B.; Zhang, Y.; Yue, Q. Numb Inhibits Cell Proliferation, Invasion, and Epithelial-Mesenchymal Transition through PAK1/β-Catenin Signaling Pathway in Ovarian Cancer. OncoTargets Ther. 2019, 12, 3223–3233. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Tang, J.; Sheng, W.; Sun, J.; Ma, Y.; Dong, M. ATP11A Promotes EMT by Regulating Numb PRRL in Pancreatic Cancer Cells. PeerJ 2022, 10, e13172. [Google Scholar] [CrossRef]
- Sheng, W.; Tang, J.; Cao, R.; Shi, X.; Ma, Y.; Dong, M. Numb-PRRL Promotes TGF-Β1- and EGF-Induced Epithelial-to-Mesenchymal Transition in Pancreatic Cancer. Cell Death Dis. 2022, 13, 173. [Google Scholar] [CrossRef]
- Lau, K.M.; McGlade, C.J. Numb Is a Negative Regulator of HGF Dependent Cell Scattering and Rac1 Activation. Exp. Cell Res. 2011, 317, 539–551. [Google Scholar] [CrossRef] [PubMed]
- Santolini, E.; Puri, C.; Salcini, A.E.; Gagliani, M.C.; Pelicci, P.G.; Tacchetti, C.; Paolo, P.; Fiore, D. Numb Is an Endocytic Protein. J. Cell Biol. 2000, 151, 1345–1352. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.A.; Dho, S.E.; Donaldson, J.; Tepass, U.; Jane McGlade, C.; Labatt Brain, S. The Cell Fate Determinant Numb Interacts with EHD/ Rme-1 Family Proteins and Has a Role in Endocytic Recycling. Mol. Biol. Cell 2004, 15, 3698–3708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Confalonieri, S.; Colaluca, I.N.; Basile, A.; Pece, S.; di Fiore, P.P. Exon 3 of the NumB Gene Emerged in the Chordate Lineage Coopting the Numb Protein to the Regulation of Mdm2. G3: Genes Genomes Genet. 2019, 9, 3359–3367. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Fre, S.; Slepnev, V.I.; Capua, M.R.; Takei, K.; Butler, M.H.; di Fiore, P.P.; de Camilli, P. Epsin Is an EH-Domain-Binding Protein Implicated in Clathrin-Mediated Endocytosis. Nature 1998, 394, 793–797. [Google Scholar] [CrossRef] [PubMed]
- Krieger, J.R.; Taylor, P.; Gajadhar, A.S.; Guha, A.; Moran, M.F.; McGlade, C.J. Identification and Selected Reaction Monitoring (SRM) Quantification of Endocytosis Factors Associated with Numb. Mol. Cell. Proteom. 2013, 12, 499–514. [Google Scholar] [CrossRef] [Green Version]
- Tang, H.; Rompani, S.B.; Atkins, J.B.; Zhou, Y.; Osterwalder, T.; Zhong, W. Numb Proteins Specify Asymmetric Cell Fates via an Endocytosis- and Proteasome-Independent Pathway. Mol. Cell. Biol. 2005, 25, 2899–2909. [Google Scholar] [CrossRef] [Green Version]
- Couturier, L.; Vodovar, N.; Schweisguth, F. Endocytosis by Numb Breaks Notch Symmetry at Cytokinesis. Nat. Cell Biol. 2012, 14, 131–139. [Google Scholar] [CrossRef]
- Spana, E.P.; Doe, C.Q. Numb antagonizes Notch signaling to specify sibling neuron cell fates. Neuron 1996, 17, 21–26. [Google Scholar] [CrossRef] [Green Version]
- Couturier, L.; Mazouni, K.; Schweisguth, F. Numb Localizes at Endosomes and Controls the Endosomal Sorting of Notch after Asymmetric Division in Drosophila. Curr. Biol. 2013, 23, 588–593. [Google Scholar] [CrossRef]
- Langevin, J.; le Borgne, R.; Rosenfeld, F.; Gho, M.; Schweisguth, F.; Bellaïche, Y. Lethal Giant Larvae Controls the Localization of Notch-Signaling Regulators Numb, Neuralized, and Sanpodo in Drosophila Sensory-Organ Precursor Cells. Curr. Biol. 2005, 15, 955–962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berdnik, D.; Tö, T.; Gonzá Lez-Gaitá, M.; Knoblich, J.A. The Endocytic Protein-Adaptin Is Required for Numb-Mediated Asymmetric Cell Division in Drosophila. Dev. Cell 2002, 3, 221–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGill, M.A.; Dho, S.E.; Weinmaster, G.; McGlade, C.J. Numb Regulates Post-Endocytic Trafficking and Degradation of Notch1. J. Biol. Chem. 2009, 284, 26427–26438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kandachar, V.; Roegiers, F. Endocytosis and Control of Notch Signaling. Curr. Opin. Cell Biol. 2012, 24, 534–540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jafar-Nejad, H.; Norga, K.; Bellen, H. Numb: “Adapting” notch for endocytosis. Dev. Cell 2002, 10, 155–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, Y.; Lu, B. Interaction of Notch Signaling Modulator Numb with α-Adaptin Regulates Endocytosis of Notch Pathway Components and Cell Fate Determination of Neural Stem Cells. J. Biol. Chem. 2012, 287, 17716–17728. [Google Scholar] [CrossRef] [Green Version]
- Couturier, L.; Trylinski, M.; Mazouni, K.; Darnet, L.; Schweisguth, F. A Fluorescent Tagging Approach in Drosophila Reveals Late Endosomal Trafficking of Notch and Sanpodo. J. Cell Biol. 2014, 207, 351–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, X.; Zitserman, D.; Serebriiskii, I.; Andrake, M.; Dunbrack, R.; Roegiers, F.; Gonzalez-Gaitan, M. Numb Independently Antagonizes Sanpodo Membrane Targeting and Notch Signaling in Drosophila Sensory Organ Precursor Cells. Mol. Biol. Cell 2010, 21, 802–810. [Google Scholar] [CrossRef] [Green Version]
- Hutterer, A.; Knoblich, J.A. Numb and α-Adaptin Regulate Sanpodo Endocytosis to Specify Cell Fate in Drosophila External Sensory Organs. EMBO Rep. 2005, 6, 836–842. [Google Scholar] [CrossRef] [Green Version]
- Couturier, L.; Mazouni, K.; Schweisguth, F. Inhibition of Notch Recycling by Numb Relevance and Mechanism(s). Cell Cycle 2013, 12, 1647–1648. [Google Scholar] [CrossRef]
- O’Connor-Giles, K.M.; Skeath, J.B. Numb Inhibits Membrane Localization of Sanpodo, a Four-Pass Transmembrane Protein, to Promote Asymmetric Divisions in Drosophila. Dev. Cell 2003, 5, 231–243. [Google Scholar] [CrossRef] [Green Version]
- Johnson, S.A.; Zitserman, D.; Roegiers, F. Numb Regulates the Balance between Notch Recycling and Late-Endosome Targeting in Drosophila Neural Progenitor Cells. Mol. Biol. Cell 2016, 27, 2857–2866. [Google Scholar] [CrossRef] [PubMed]
- Huang, E.J.; Li, H.; Tang, A.A.; Wiggins, A.K.; Neve, R.L.; Zhong, W.; Jan, L.Y.; Yuh, N.J. Targeted Deletion of Numb and Numblike in Sensory Neurons Reveals Their Essential Functions in Axon Arborization. Genes Dev. 2005, 19, 138–151. [Google Scholar] [CrossRef] [Green Version]
- Shao, X.; Ding, Z.; Zhao, M.; Liu, K.; Sun, H.; Chen, J.; Liu, X.; Zhang, Y.; Hong, Y.; Li, H.; et al. Mammalian Numb Protein Antagonizes Notch by Controlling Postendocytic Trafficking of the Notch Ligand Delta-like 4. J. Biol. Chem. 2017, 292, 20628–20643. [Google Scholar] [CrossRef] [Green Version]
- Bogdanović, O.; Delfino-Machín, M.; Nicolás-Pérez, M.; Gavilán, M.P.; Gago-Rodrigues, I.; Fernández-Miñán, A.; Lillo, C.; Ríos, R.M.; Wittbrodt, J.; Martínez-Morales, J.R. Numb/Numbl-Opo Antagonism Controls Retinal Epithelium Morphogenesis by Regulating Integrin Endocytosis. Dev. Cell 2012, 23, 782–795. [Google Scholar] [CrossRef] [Green Version]
- Nishimura, T.; Kaibuchi, K. Numb Controls Integrin Endocytosis for Directional Cell Migration with APKC and PAR-3. Dev. Cell 2007, 13, 15–28. [Google Scholar] [CrossRef] [Green Version]
- Gibbs, B.C.; Shenje, L.; Andersen, P.; Miyamoto, M.; Kwon, C. Β1-Integrin Is a Cell-Autonomous Factor Mediating the Numb Pathway for Cardiac Progenitor Maintenance. Biochem. Biophys. Res. Commun. 2018, 500, 256–260. [Google Scholar] [CrossRef]
- Sato, K.; Watanabe, T.; Wang, S.; Kakeno, M.; Matsuzawa, K.; Matsui, T.; Yokoi, K.; Murase, K.; Sugiyama, I.; Ozawa, M.; et al. Numb Controls E-Cadherin Endocytosis through P120 Catenin with APKC. Mol. Biol. Cell 2011, 22, 3103–3119. [Google Scholar] [CrossRef] [PubMed]
- Krieger, J.R.; Taylor, P.; Moran, M.F.; Mcglade, C.J. Comprehensive Identification of Phosphorylation Sites on the Numb Endocytic Adaptor Protein. Proteomics 2015, 15, 434–446. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Liu, Z.; Shan, Z.; Yao, W.; Gu, A.; Wen, W. Structural Determinants Controlling 14-3-3 Recruitment to the Endocytic Adaptor Numb and Dissociation of the Numb α-Adaptin Complex. J. Biol. Chem. 2018, 293, 4149–4158. [Google Scholar] [CrossRef] [PubMed]
- Li, P.S.; Fu, Z.Y.; Zhang, Y.Y.; Zhang, J.H.; Xu, C.Q.; Ma, Y.T.; Li, B.L.; Song, B.L. The Clathrin Adaptor Numb Regulates Intestinal Cholesterol Absorption through Dynamic Interaction with NPC1L1. Nat. Med. 2014, 20, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Tokumitsu, H.; Hatano, N.; Inuzuka, H.; Sueyoshi, Y.; Yokokura, S.; Ichimura, T.; Nozaki, N.; Kobayashi, R. Phosphorylation of Numb Family Proteins: Possible Involvement of Ca 2+/Calmodulin-Dependent Protein Kinases. J. Biol. Chem. 2005, 280, 35108–35118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tokumitsu, H.; Hatano, N.; Yokokura, S.; Sueyoshi, Y.; Nozaki, N.; Kobayashi, R. Phosphorylation of Numb Regulates Its Interaction with the Clathrin-Associated Adaptor AP-2. FEBS Lett. 2006, 580, 5797–5801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdi, K.; Neves, G.; Pyun, J.; Kiziltug, E.; Ahrens, A.; Kuo, C.T. EGFR Signaling Termination via Numb Trafficking in Ependymal Progenitors Controls Postnatal Neurogenic Niche Differentiation. Cell Rep. 2019, 28, 2012–2022.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sorensen, E.B.; Conner, S.D. AAK1 Regulates Numb Function at an Early Step in Clathrin-Mediated Endocytosis. Traffic 2008, 9, 1791–1800. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Wang, D.D.; Hou, X.; Li, X.; Shen, Y. Different Roles of Numb-P72 and Numb-P65 on the Trafficking of Metabotropic Glutamate Receptor 5. Mol. Biol. Rep. 2021, 48, 595–600. [Google Scholar] [CrossRef]
- Wang, N.; Cai, X.Y.; Gao, W.; Shen, Y. Numb-P72, but Not Numb-P65, Contributes to the Trafficking of Group i Metabotropic Glutamate Receptors. Neuroreport 2018, 29, 902–906. [Google Scholar] [CrossRef]
- Wang, N.; dan Wang, D.; Shen, Y. Numb Deficiency Causes Impaired Trafficking of MGlu5 in Neurons and Autistic-like Behaviors. Neurosci. Lett. 2019, 707, 134291. [Google Scholar] [CrossRef]
- Lu, C.B.; Fu, W.; Xu, X.; Mattson, M.P. Numb-Mediated Neurite Outgrowth Is Isoform-Dependent, and Requires Activation of Voltage-Dependent Calcium Channels. Neuroscience 2009, 161, 403–412. [Google Scholar] [CrossRef] [Green Version]
- Ntelios, D.; Berninger, B.; Tzimagiorgis, G. Numb and Alzheimer’s Disease: The Current Picture. Front. Neurosci. 2012, 6, 145. [Google Scholar] [CrossRef]
- Merthan, L.; Haller, A.; Thal, D.R.; von Einem, B.; von Arnim, C.A.F. The Role of PTB Domain Containing Adaptor Proteins on PICALM-Mediated APP Endocytosis and Localization. Biochem. J. 2019, 476, 2093–2109. [Google Scholar] [CrossRef] [PubMed]
- Matrone, C. A New Molecular Explanation for Age-Related Neurodegeneration: The Tyr682 Residue of Amyloid Precursor Protein. BioEssays 2013, 35, 847–852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez, K.; Ran, D.; Madura, C.L.; Moutal, A.; Khanna, R. Non-SUMOylated CRMP2 Decreases NaV1.7 Currents via the Endocytic Proteins Numb, Nedd4-2 and Eps15. Mol. Brain 2021, 14, 20. [Google Scholar] [CrossRef]
- Nishimura, T.; Fukata, Y.; Kato, K.; Yamaguchi, T.; Matsuura, Y.; Kamiguchi, H.; Kaibuchi, K. CRMP-2 Regulates Polarized Numb-Mediated Endocytosis for Axon Growth. Nat. Cell Biol. 2003, 5, 819–826. [Google Scholar] [CrossRef]
- Wei, R.; Liu, X.; Voss, C.; Qin, W.; Dagnino, L.; Li, L.; Vigny, M.; Li, S.S.C. NUMB Regulates the Endocytosis and Activity of the Anaplastic Lymphoma Kinase in an Isoform-Specific Manner. J. Mol. Cell Biol. 2019, 11, 994–1005. [Google Scholar] [CrossRef] [Green Version]
- Wei, R.; Kaneko, T.; Liu, X.; Liu, H.; Li, L.; Voss, C.; Liu, E.; He, N.; Li, S.S.C. Interactome Mapping Uncovers a General Role for Numb in Protein Kinase Regulation. Mol. Cell. Proteom. 2018, 17, 2216–2228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sorrell, F.J.; Szklarz, M.; Abdul Azeez, K.R.; Elkins, J.M.; Knapp, S. Family-Wide Structural Analysis of Human Numb-Associated Protein Kinases. Structure 2016, 24, 401–411. [Google Scholar] [CrossRef] [Green Version]
- Beres, B.J.; George, R.; Lougher, E.J.; Barton, M.; Verrelli, B.C.; McGlade, C.J.; Rawls, J.A.; Wilson-Rawls, J. Numb Regulates Notch1, but Not Notch3, during Myogenesis. Mech. Dev. 2011, 128, 247–257. [Google Scholar] [CrossRef]
- Luo, Z.; Mu, L.; Zheng, Y.; Shen, W.; Li, J.; Xu, L.; Zhong, B.; Liu, Y.; Zhou, Y. NUMB Enhances Notch Signaling by Repressing Ubiquitination of NOTCH1 Intracellular Domain. J. Mol. Cell Biol. 2020, 12, 345–358. [Google Scholar] [CrossRef]
- Givogri, M.I.; Schonmann, V.; Cole, R.; de Vellis, J.; Bongarzone, E.R. Notch1 and Numb Genes Are Inversely Expressed as Oligodendrocytes Differentiate. Dev. Neurosci. 2003, 25, 50–64. [Google Scholar] [CrossRef]
- Miao, L.; Lu, Y.; Nusrat, A.; Abdelnasser, H.Y.; Datta, S.; Zhou, B.; Schwartz, R.J.; Wu, M. The Spatiotemporal Expression of Notch1 and Numb and Their Functional Interaction during Cardiac Morphogenesis. Cells 2021, 10, 2192. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Bücker, S.; Jungblut, B.; Böttger, T.; Cinnamon, Y.; Tchorz, J.; Müller, M.; Bettler, B.; Harvey, R.; Sun, Q.Y.; et al. Inhibition of Notch2 by Numb/Numblike Controls Myocardial Compaction in the Heart. Cardiovasc. Res. 2012, 96, 276–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kikuchi, H.; Sakakibara-Konishi, J.; Furuta, M.; Kikuchi, E.; Kikuchi, J.; Oizumi, S.; Hida, Y.; Kaga, K.; Kinoshita, I.; Dosaka-Akita, H.; et al. Numb Has Distinct Function in Lung Adenocarcinoma and Squamous Cell Carcinoma. Oncotarget 2018, 9, 29379–29391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kikuchi, H.; Sakakibara-Konishi, J.; Furuta, M.; Yokouchi, H.; Nishihara, H.; Yamazaki, S.; Uramoto, H.; Tanaka, F.; Harada, M.; Akie, K.; et al. Expression of Notch1 and Numb in Small Cell Lung Cancer. Oncotarget 2017, 8, 10485–10497. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.H.; Wu, Y.; Yao, S.; Levine, A.C.; Kirschenbaum, A.; Collier, L.; Bauman, W.A.; Cardozo, C.P. Androgens Up-Regulate Transcription of the Notch Inhibitor Numb in C2C12 Myoblasts via Wnt/β-Catenin Signaling to t Cell Factor Elements in the Numb Promoter. J. Biol. Chem. 2013, 288, 17990–17998. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Lanner, F.; Lendahl, U.; Das, D. Numblike and Numb Differentially Affect P53 and Sonic Hedgehog Signaling. Biochem. Biophys. Res. Commun. 2011, 413, 426–431. [Google Scholar] [CrossRef]
- di Marcotullio, L.; Greco, A.; Mazzà, D.; Canettieri, G.; Pietrosanti, L.; Infante, P.; Coni, S.; Moretti, M.; de Smaele, E.; Ferretti, E.; et al. Numb Activates the E3 Ligase Itch to Control Gli1 Function through a Novel Degradation Signal. Oncogene 2011, 30, 65–76. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Chen, A.; Jamieson, C.H.; Fereshteh, M.; Abrahamsson, A.; Blum, J.; Kwon, H.Y.; Kim, J.; Chute, J.P.; Rizzieri, D.; et al. Hedgehog Signalling Is Essential for Maintenance of Cancer Stem Cells in Myeloid Leukaemia. Nature 2009, 458, 776–779. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Zhang, K.; Cheng, C.; Ji, Z.; Wang, X.; Wang, M.; Chu, M.; Tang, D.G.; Zhu, H.H.; Gao, W.Q. Numb-/Low Enriches a Castration-Resistant Prostate Cancer Cell Subpopulation Associated with Enhanced Notch and Hedgehog Signaling. Clin. Cancer Res. 2017, 23, 6744–6756. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Sheng, W.; Tang, J.; Li, X.; Zhou, J.; Dong, M. Cooperation of SRPK2, Numb and P53 in the Malignant Biology and Chemosensitivity of Colorectal Cancer. Biosci. Rep. 2020, 40, BSR20191488. [Google Scholar] [CrossRef]
- Sheng, W.; Dong, M.; Chen, C.; Wang, Z.; Li, Y.; Wang, K.; Li, Y.; Zhou, J. Cooperation of Musashi-2, Numb, MDM2, and P53 in Drug Resistance and Malignant Biology of Pancreatic Cancer. FASEB J. 2017, 31, 2429–2438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xian, J.; Cheng, Y.; Qin, X.; Cao, Y.; Luo, Y.; Cao, Y. Progress in the Research of P53 Tumour Suppressor Activity Controlled by Numb in Triple-Negative Breast Cancer. J. Cell. Mol. Med. 2020, 24, 7451–7459. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Ronai, Z.A. Rewired Notch/P53 by Numb’ing Mdm2. J. Cell Biol. 2018, 217, 445–446. [Google Scholar] [CrossRef] [Green Version]
- Sheng, W.; Dong, M.; Zhou, J.; Li, X.; Liu, Q.; Dong, Q.; Li, F. Cooperation among Numb, MDM2 and P53 in the Development and Progression of Pancreatic Cancer. Cell Tissue Res. 2013, 354, 521–532. [Google Scholar] [CrossRef] [PubMed]
- Carter, S.; Vousden, K.H. A Role for Numb in P53 Stabilization. Genome Biol. 2008, 9, 221. [Google Scholar] [CrossRef] [Green Version]
- Faraldo, M.M.; Glukhova, M.A. Regulating the Regulator: Numb Acts Upstream of P53 to Control Mammary Stem and Progenitor Cell. J. Cell Biol. 2015, 211, 737–739. [Google Scholar] [CrossRef]
- Chou, C.H.; Tu, H.F.; Kao, S.Y.; Chiang, C.Y.F.; Liu, C.J.; Chang, K.W.; Lin, S.C. Targeting of MiR-31/96/182 to the Numb Gene during Head and Neck Oncogenesis. Head Neck 2018, 40, 808–817. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Zhu, H.C.; Liu, X.H.; Wang, L.; Ning, J.Z.; Xiao, C.C. MiR-543 Promotes Proliferation and Epithelial-Mesenchymal Transition in Prostate Cancer via Targeting RKIP. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2017, 41, 1135–1146. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zheng, W.; Li, D.; Zheng, J. MiR-146a-5p Promotes Chondrocyte Apoptosis and Inhibits Autophagy of Osteoarthritis by Targeting NUMB. Cartilage 2021, 13, 1467S–1477S. [Google Scholar] [CrossRef]
- Wu, Y.C.; Lee, K.S.; Song, Y.; Gehrke, S.; Lu, B. The Bantam MicroRNA Acts through Numb to Exert Cell Growth Control and Feedback Regulation of Notch in Tumor-Forming Stem Cells in the Drosophila Brain. PLoS Genet. 2017, 13, e1006785. [Google Scholar] [CrossRef]
- Brennecke, J.; Hipfner, D.R.; Stark, A.; Russell, R.B.; Cohen, S.M. Bantam Encodes a Developmentally Regulated MicroRNA That Controls Cell Proliferation and Regulates the Proapoptotic Gene Hid in Drosophila. Cell 2003, 113, 25–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, W.L.; Jiang, J.K.; Yang, S.H.; Huang, T.S.; Lan, H.Y.; Teng, H.W.; Yang, C.Y.; Tsai, Y.P.; Lin, C.H.; Wang, H.W.; et al. MicroRNA-146a Directs the Symmetric Division of Snail-Dominant Colorectal Cancer Stem Cells. Nat. Cell Biol. 2014, 16, 268–280. [Google Scholar] [CrossRef] [PubMed]
- Hung, P.S.; Liu, C.J.; Chou, C.S.; Kao, S.Y.; Yang, C.C.; Chang, K.W.; Chiu, T.H.; Lin, S.C. MiR-146a Enhances the Oncogenicity of Oral Carcinoma by Concomitant Targeting of the IRAK1, TRAF6 and NUMB Genes. PLoS ONE 2013, 8, e79926. [Google Scholar] [CrossRef] [PubMed]
- Kuang, W.; Tan, J.; Duan, Y.; Duan, J.; Wang, W.; Jin, F.; Jin, Z.; Yuan, X.; Liu, Y. Cyclic Stretch Induced MiR-146a Upregulation Delays C2C12 Myogenic Differentiation through Inhibition of Numb. Biochem. Biophys. Res. Commun. 2009, 378, 259–263. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.J.; Ma, P.; Ping, S.; Hong, S.; Yunjia, L. MicroRNA-146a contributes to CD4+ T lymphocyte differentiation in patients with thyroid ophthalmopathy. Am. J. Transl. Res. 2017, 94, 1801–1809. [Google Scholar]
- Ma, X.L.; Zhang, X.J.; Du, Q.; Zhang, X.N.; Zhang, S.Y.; Zhao, H.F. MicroRNA-146b Promotes Neuroblastoma Cell Growth through Targeting NUMB. Exp. Ther. Med. 2020, 19, 3531–3536. [Google Scholar] [CrossRef] [Green Version]
- Peng, H.; Wang, L.; Su, Q.; Yi, K.; Du, J.; Wang, Z. MiR-31-5p Promotes the Cell Growth, Migration and Invasion of Colorectal Cancer Cells by Targeting NUMB. Biomed. Pharmacother. 2019, 109, 208–216. [Google Scholar] [CrossRef]
- Wang, X.; Cai, J.; Zhao, L.; Zhang, D.; Xu, G.; Hu, J.; Zhang, T.; Jin, M. NUMB Suppression by MiR-9-5P Enhances CD44+ Prostate Cancer Stem Cell Growth and Metastasis. Sci. Rep. 2021, 11, 11210. [Google Scholar] [CrossRef]
- Dou, X.Q.; Chen, X.J.; Zhou, Q.; Wen, M.X.; Zhang, S.Z.; Zhang, S.Q. MiR-335 Modulates Numb Alternative Splicing via Targeting RBM10 in Endometrial Cancer. Kaohsiung J. Med. Sci. 2020, 36, 171–177. [Google Scholar] [CrossRef] [Green Version]
- Zheng, L.; Song, J.; Tang, R.; Chen, X.; Wang, L.; Wu, D.; Cen, H.; Shi, L. MicroRNA-524-5p Regulates the Proliferation and Invasion of HTR-8/SVneo Trophoblasts by Targeting NUMB in the Notch Signaling Pathway. Mol. Med. Rep. 2021, 23, 436. [Google Scholar] [CrossRef] [PubMed]
- Ianni, A.; Hofmann, M.; Kumari, P.; Tarighi, S.; Al-Tamari, H.M.; Görgens, A.; Giebel, B.; Nolte, H.; Krüger, M.; Salwig, I.; et al. Depletion of Numb and Numblike in Murine Lung Epithelial Cells Ameliorates Bleomycin-Induced Lung Fibrosis by Inhibiting the β-Catenin Signaling Pathway. Front. Cell Dev. Biol. 2021, 9, 639162. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.; Liu, W.; Li, T.; Wan, J.; Tian, J.; Zhou, Z.; Li, H.; Liu, Y.; Hou, F.F.; Nie, J. Numb Contributes to Renal Fibrosis by Promoting Tubular Epithelial Cell Cycle Arrest at G2/M. Oncotarget 2016, 7, 25604–25619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, X.; Cao, Y. Prognostic Biomarker NUMB Is Inhibited by Breast Cancer Cell Exosomes to Promote Breast Cancer Progression. J. Immunol. Res. 2022, 2022, 6032076. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Kang, Y.; Ma, R.; Chen, F.; Chen, F.; Dong, X. Expression of Numb and Gli1 in Malignant Pleural Mesothelioma and Their Clinical Significance. J. Cancer Res. Ther. 2018, 14, 970–976. [Google Scholar] [PubMed]
- Wang, C.; Feng, W.; Zhang, C. The Expression and Function of NUMB in Endometrial Cancer and the Interaction with HDM2 and P53. J. Cancer 2015, 6, 1030–1040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Shao, X.; Sun, H.; Liu, K.; Ding, Z.; Chen, J.; Fang, L.; Su, W.; Hong, Y.; Li, H.; et al. NUMB Negatively Regulates the Epithelial-Mesenchymal Transition of Triple-Negative Breast Cancer by Antagonizing Notch Signaling. Oncotarget 2016, 7, 61036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonulcu, S.C.; Unal, B.; Bassorgun, I.C.; Ozcan, M.; Coskun, H.S.; Elpek, G.O. Expression of Notch Pathway Components (Numb, Itch, and Siah-1) in Colorectal Tumors: A Clinicopathological Study. World J. Gastroenterol. 2020, 26, 3814–3833. [Google Scholar] [CrossRef]
- Bi, Y.L.; Min, M.; Shen, W.; Liu, Y. Numb/Notch Signaling Pathway Modulation Enhances Human Pancreatic Cancer Cell Radiosensitivity. Tumor Biol. 2016, 37, 15145–15155. [Google Scholar] [CrossRef] [PubMed]
- Flores, A.N.; McDermott, N.; Meunier, A.; Marignol, L. NUMB Inhibition of NOTCH Signalling as a Therapeutic Target in Prostate Cancer. Nat. Rev. Urol. 2014, 11, 499–507. [Google Scholar]
- Hristova, D.M.; Fukumoto, T.; Takemori, C.; Gao, L.; Hua, X.; Wang, J.X.; Li, L.; Beqiri, M.; Watters, A.; Vultur, A.; et al. NUMB as a Therapeutic Target for Melanoma. J. Investig. Dermatol. 2022, 142, 1882–1892.e5. [Google Scholar] [CrossRef]
- Shu, Y.; Xu, Q.; Xu, Y.; Tao, Q.; Shao, M.; Cao, X.; Chen, Y.; Wu, Z.; Chen, M.; Zhou, Y.; et al. Loss of Numb Promotes Hepatic Progenitor Expansion and Intrahepatic Cholangiocarcinoma by Enhancing Notch Signaling. Cell Death Dis. 2021, 12, 966. [Google Scholar] [CrossRef]
- Jiang, X.; Xing, H.; Kim, T.M.; Jung, Y.; Huang, W.; Yang, H.W.; Song, S.; Park, P.J.; Carroll, R.S.; Johnson, M.D. Numb Regulates Glioma Stem Cell Fate and Growth by Altering Epidermal Growth Factor Receptor and Skp1-Cullin-F-Box Ubiquitin Ligase Activity. Stem Cells 2012, 30, 1313–1326. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Shen, S.L.; Chen, B.; Nie, J.; Peng, B.G. Numb Promotes Cell Proliferation and Correlates with Poor Prognosis in Hepatocellular Carcinoma. PLoS ONE 2014, 9, e95849. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Luo, C.; Yang, W.; Wang, K.; Hu, C.; Zou, J.; Zhu, H.; Fu, L.; Nie, J.; Cheng, C. Increased Numb Protein Expression Predicts Poor Clinical Outcomes in Esophageal Squamous Cell Carcinoma Patients. Cancer Biol. Ther. 2018, 19, 34–41. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Xu, W.; Ji, J.; Feng, D.; Sourbier, C.; Yang, Y.; Qu, J.; Zeng, Z.; Wang, C.; Chang, X.; et al. Alternative Splicing of the Cell Fate Determinant Numb in Hepatocellular Carcinoma. Hepatology 2015, 62, 1122–1131. [Google Scholar] [CrossRef] [Green Version]
- Hong, J.; Liu, Z.; Zhu, H.; Zhang, X.; Liang, Y.; Yao, S.; Wang, F.; Xie, X.; Zhang, B.; Tan, T.; et al. The Tumor Suppressive Role of NUMB Isoform 1 in Esophageal Squamous Cell Carcinoma. Oncotarget 2014, 5, 5602–5614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, Y.; Ding, M.; Tian, G.; Guo, H.; Wan, Y.; Yao, Z.; Li, B.; Lin, D. Overexpression of Numb Suppresses Tumor Cell Growth and Enhances Sensitivity to Cisplatin in Epithelioid Malignant Pleural Mesothelioma. Oncol. Rep. 2013, 30, 313–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Alegría, E.; Carmen Lafita-Navarro, M.; Aguado, R.; Garcia-Gutierrez, L.; Sarnataro, K.; Ruiz-Herguido, C.; Martín, F.; Bigas, A.; Canelles, M.; León, J. NUMB Inactivation Confers Resistance to Imatinib in Chronic Myeloid Leukemia Cells. Cancer Lett. 2016, 375, 92–99. [Google Scholar] [CrossRef] [Green Version]
- Shen, E.D.; Zeng, Q. Inhibition of the Numb/Notch Signaling Pathway Increases Radiation Sensitivity in Human Nasopharyngeal Carcinoma Cells. Kaohsiung J. Med. Sci. 2019, 35, 474–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shan, G.P.; Zhang, P.; Li, P.; Du, F.L.; Yang, Y.W. Numb Gene Enhances Radiation Sensitivity of Nonsmall Cell Lung Cancer Stem Cells. Cancer Biother. Radiopharm. 2016, 31, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Dong, Y.; Maeda, U.; Xia, W.; Tanzi, R.E. RNAi-Mediated Knock-down of Dab and Numb Attenuate Aβ Levels via γ-Secretase Mediated APP Processing. Transl. Neurodegener. 2012, 1, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chigurupati, S.; Madan, M.; Okun, E.; Wei, Z.; Pattisapu, J.v.; Mughal, M.R.; Mattson, M.P.; Chan, S.L. Evidence for Altered Numb Isoform Levels in Alzheimer’s Disease Patients and a Triple Transgenic Mouse Model. J. Alzheimer’s Dis. 2011, 24, 349–361. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.; Ma, M.; Teng, J.; Shao, F.; Wu, E.; Wang, X. Numb Protects Human Renal Tubular Epithelial Cells from Bovine Serum Albumin-Induced Apoptosis Through Antagonizing CHOP/PERK Pathway. J. Cell. Biochem. 2016, 117, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Wu, Y.; Yu, F.; Hu, W.; Fang, X.; Hao, W. The Implication of Numb-Induced Notch Signaling in Endothelial-Mesenchymal Transition of Diabetic Nephropathy. J. Diabetes Complicat. 2018, 32, 889–899. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Li, Y.; He, Y.; Wang, J. Numb Promotes Autophagy through P53 Pathway in Acute Kidney Injury Induced by Cisplatin. Anal. Cell. Pathol. 2022, 2022, 8213683. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.; Li, H.; Long, T.; Zhou, M.; Wan, J.; Tian, J.; Zhou, Z.; Hu, Z.; Nie, J. Tubular Numb Promotes Renal Interstitial Fibrosis via Modulating HIF-1α Protein Stability. Biochim. Biophys. Acta Mol. Basis Dis. 2021, 1867, 166081. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Li, H.; Su, J.; Xu, S.; Zhu, F.; Ai, J.; Hu, Z.; Zhou, M.; Tian, J.; Su, Z.; et al. Numb Depletion Promotes Drp1-Mediated Mitochondrial Fission and Exacerbates Mitochondrial Fragmentation and Dysfunction in Acute Kidney Injury. Antioxid. Redox Signal. 2019, 30, 1797–1816. [Google Scholar] [CrossRef] [PubMed]
- Meng, H.; Li, L.; Ruan, J.; Chen, Y.; Yan, Z.; Liu, J.; Li, X.; Mao, C.; Yang, P. Association of Low Expression of NUMB in Peripheral Blood with Acute Myocardial Infarction. Cardiol. Res. Pract. 2022, 2022, 7981637. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Guo, H.; Li, J.; Myint, T.; Pittman, W.; Yang, L.; Zhong, W.; Schwartz, R.J.; Schwarz, J.J.; Singer, H.A.; et al. Numb Family Proteins Are Essential for Cardiac Morphogenesis and Progenitor Differentiation. Development 2014, 141, 281–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- George, R.M.; Biressi, S.; Beres, B.J.; Rogers, E.; Mulia, A.K.; Allen, R.E.; Rawls, A.; Rando, T.A.; Wilson-Rawls, J. Numb-Deficient Satellite Cells Have Regeneration and Proliferation Defects. Proc. Natl. Acad. Sci. USA 2013, 110, 18549–18555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, M.; Wang, X.; Ding, X.; Teng, J.; Shao, F.; Zhang, J. Numb/Notch Signaling Plays an Important Role in Cerebral Ischemia-Induced Apoptosis. Neurochem. Res. 2013, 38, 254–261. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
NUMB isoforms are generated by alternative splicing, being the PTB and PRR domains involved in this process. The resulting isoforms can be classified as having long or short PTB (PTBL/S) and long or short PRR (PRRL/S) domains. Please note that the sizes of exons and domains in the diagram are not to scale and are simplified for clarity.
Figure 1.
NUMB isoforms are generated by alternative splicing, being the PTB and PRR domains involved in this process. The resulting isoforms can be classified as having long or short PTB (PTBL/S) and long or short PRR (PRRL/S) domains. Please note that the sizes of exons and domains in the diagram are not to scale and are simplified for clarity.

Figure 2.
(A) NUMB and NOTCH are asymmetrically distributed to daughter cells during SOPs in Drosophila. (B) NUMB and NOTCH are involved in determining cell fate in Drosophila neuroblasts. The cell that inherits NOTCH (Neuroblast) has self-renewal properties, while the cell that inherits NUMB (GMC: ganglion mother cell) is able to differentiate into neurons. (C) The balance of NUMB and NOTCH is also thought to be involved in EMT in mice and humans tumors. Increased NUMB expression relative to NOTCH has been shown to reduce EMT, while increased NOTCH levels can increase EMT.
Figure 2.
(A) NUMB and NOTCH are asymmetrically distributed to daughter cells during SOPs in Drosophila. (B) NUMB and NOTCH are involved in determining cell fate in Drosophila neuroblasts. The cell that inherits NOTCH (Neuroblast) has self-renewal properties, while the cell that inherits NUMB (GMC: ganglion mother cell) is able to differentiate into neurons. (C) The balance of NUMB and NOTCH is also thought to be involved in EMT in mice and humans tumors. Increased NUMB expression relative to NOTCH has been shown to reduce EMT, while increased NOTCH levels can increase EMT.

Figure 3.
NUMB acts as an adaptor protein in multiple signaling pathways. NUMB inhibits NOTCH signaling by interacting with NICD. Additionally, the interaction between NUMB and MDM2 increases p53 stability, allowing transcription of genes related to DNA-damage repair. Furthermore, the PRRL NUMB isoform promotes ERK/MAPK signaling and the formation of a SMAD complex, leading to transcription of genes related to EMT. In addition, NUMB promotes the ubiquitination and subsequent degradation of both NUMB and ITCH through GLI1.
Figure 3.
NUMB acts as an adaptor protein in multiple signaling pathways. NUMB inhibits NOTCH signaling by interacting with NICD. Additionally, the interaction between NUMB and MDM2 increases p53 stability, allowing transcription of genes related to DNA-damage repair. Furthermore, the PRRL NUMB isoform promotes ERK/MAPK signaling and the formation of a SMAD complex, leading to transcription of genes related to EMT. In addition, NUMB promotes the ubiquitination and subsequent degradation of both NUMB and ITCH through GLI1.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ortega-Campos, S.M.; García-Heredia, J.M. The Multitasker Protein: A Look at the Multiple Capabilities of NUMB. Cells 2023, 12, 333. https://doi.org/10.3390/cells12020333
AMA Style
Ortega-Campos SM, García-Heredia JM. The Multitasker Protein: A Look at the Multiple Capabilities of NUMB. Cells. 2023; 12(2):333. https://doi.org/10.3390/cells12020333
Chicago/Turabian StyleOrtega-Campos, Sara M., and José Manuel García-Heredia. 2023. "The Multitasker Protein: A Look at the Multiple Capabilities of NUMB" Cells 12, no. 2: 333. https://doi.org/10.3390/cells12020333
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.