
GABAA and Glycine Receptor-Mediated Inhibitory Synaptic Transmission onto Adult Rat Lamina IIi PKCγ-Interneurons: Pharmacological but Not Anatomical Specialization

 , , ,
, , ,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Electrophysiology Experiments
2.2.1. Slice Preparation
2.2.2. Patch-Clamp Recordings
2.2.3. Drug Application
2.2.4. Immunofluorescence Detection of PKCγ/Neurobiotin Interneurons
2.2.5. Data Analysis
2.3. Histological Procedures
2.3.1. Immunofluorescence Detection of Inhibitory Receptors
2.3.2. GABAergic and Glycinergic Boutons on PKCγ Interneurons
2.3.3. Confocal Microscopy and Analysis
2.3.4. Characterization of Antibodies
2.4. Statistical Analysis
3. Results
3.1. Electrophysiological Analysis
3.1.1. Three Kinetically Distinct Populations of mIPSCs onto MDH Lamina IIi PKCγ+ Interneurons
3.1.2. Co-Occurrence of Fast Monoexponential, Slow Monoexponential, and Biexponential Spontaneous IPSCs in All MDH Lamina IIi PKCγ+ Interneurons
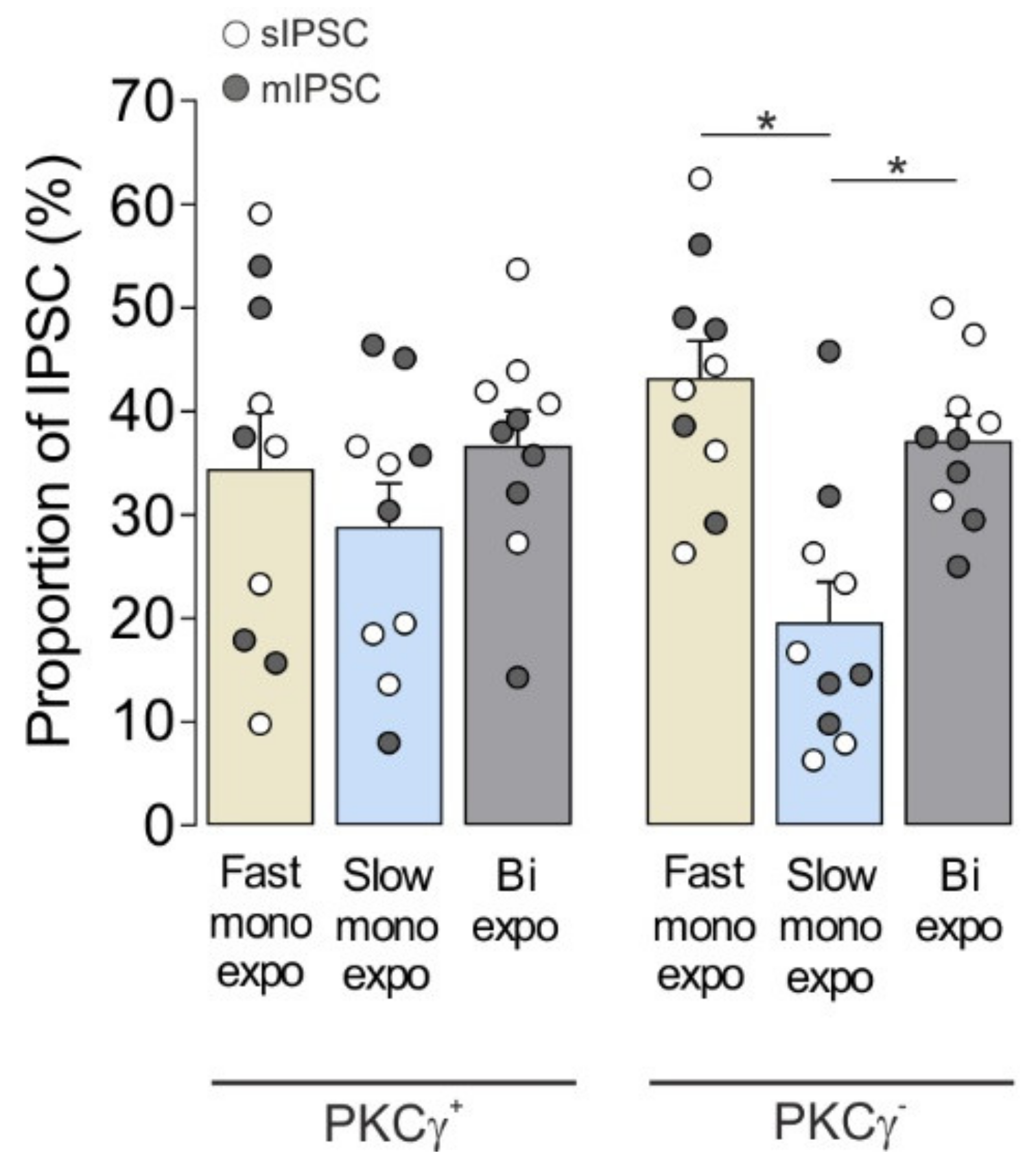
3.1.3. Co-Occurrence of Fast Monoexponential, Slow Monoexponential, and Biexponential IPSCs in MDH Lamina IIi PKCγ+ and PKCγ− Interneurons
3.1.4. GABAAR- and GlyR-Mediated sIPSCs in MDH Lamina IIi PKCγ+ and PKCγ− Interneurons
3.2. Inhibitory Receptors and Synapses onto PKCγ+ Interneurons
4. Discussion
4.1. GABAAR-Only- or GlyR-Only-Mediated IPSCs onto MDH lamina IIi PKCγ+ and PKCγ− Interneurons
4.2. PKCγ+ Interneurons Co-Express GABAAR and GlyR at the Same Inhibitory Synapses
4.3. Can Anatomical Data Account for the Functional Specialization of Inhibitory Synapses onto MDH Lamina IIi PKCγ+ Interneurons?
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bouhassira, D.; Attal, N.; Alchaar, H.; Boureau, F.; Brochet, B.; Bruxelle, J.; Cunin, G.; Fermanian, J.; Ginies, P.; Grun-Overdyking, A.; et al. Comparison of pain syndromes associated with nervous or somatic lesions and development of a new neuropathic pain diagnostic questionnaire (DN4). Pain 2005, 114, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Jensen, T.S.; Finnerup, N.B. Allodynia and hyperalgesia in neuropathic pain: Clinical manifestations and mechanisms. Lancet Neurol. 2014, 13, 924–935. [Google Scholar] [CrossRef]
- Li, L.; Rutlin, M.; Abraira, V.E.; Cassidy, C.; Kus, L.; Gong, S.; Jankowski, M.P.; Luo, W.; Heintz, N.; Koerber, H.R.; et al. The functional organization of cutaneous low-threshold mechanosensory neurons. Cell 2011, 147, 1615–1627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seal, R.P.; Wang, X.; Guan, Y.; Raja, S.N.; Woodbury, C.J.; Basbaum, A.I.; Edwards, R.H. Injury-induced mechanical hypersensitivity requires C-low threshold mechanoreceptors. Nature 2009, 462, 651–655. [Google Scholar] [CrossRef] [PubMed]
- Miraucourt, L.S.; Dallel, R.; Voisin, D.L. Glycine inhibitory dysfunction turns touch into pain through PKCgamma interneurons. PLoS ONE 2007, 2, e1116. [Google Scholar] [CrossRef] [PubMed]
- Miraucourt, L.S.; Moisset, X.; Dallel, R.; Voisin, D.L. Glycine inhibitory dysfunction induces a selectively dynamic, morphine-resistant, and neurokinin 1 receptor- independent mechanical allodynia. J. Neurosci. 2009, 29, 2519–2527. [Google Scholar] [CrossRef] [PubMed]
- Todd, A.J. Neuronal circuitry for pain processing in the dorsal horn. Nat. Rev. Neurosci. 2010, 11, 823–836. [Google Scholar] [CrossRef] [Green Version]
- Torsney, C.; MacDermott, A.B. Disinhibition opens the gate to pathological pain signaling in superficial neurokinin 1 receptor-expressing neurons in rat spinal cord. J. Neurosci. 2006, 26, 1833–1843. [Google Scholar] [CrossRef] [Green Version]
- Artola, A.; Voisin, D.; Dallel, R. PKCgamma interneurons, a gateway to pathological pain in the dorsal horn. J. Neural. Transm. 2020, 127, 527–540. [Google Scholar] [CrossRef]
- Malmberg, A.B.; Chen, C.; Tonegawa, S.; Basbaum, A.I. Preserved acute pain and reduced neuropathic pain in mice lacking PKCgamma. Science 1997, 278, 279–283. [Google Scholar] [CrossRef] [Green Version]
- Polgar, E.; Fowler, J.H.; McGill, M.M.; Todd, A.J. The types of neuron which contain protein kinase C gamma in rat spinal cord. Brain Res. 1999, 833, 71–80. [Google Scholar] [CrossRef]
- Mao, J.; Price, D.D.; Phillips, L.L.; Lu, J.; Mayer, D.J. Increases in protein kinase C gamma immunoreactivity in the spinal cord dorsal horn of rats with painful mononeuropathy. Neurosci. Lett. 1995, 198, 75–78. [Google Scholar] [CrossRef]
- Martin, W.J.; Liu, H.; Wang, H.; Malmberg, A.B.; Basbaum, A.I. Inflammation-induced up-regulation of protein kinase Cgamma immunoreactivity in rat spinal cord correlates with enhanced nociceptive processing. Neuroscience 1999, 88, 1267–1274. [Google Scholar] [CrossRef]
- Peirs, C.; Bourgois, N.; Artola, A.; Dallel, R. Protein Kinase C gamma Interneurons Mediate C-fiber-induced Orofacial Secondary Static Mechanical Allodynia, but Not C-fiber-induced Nociceptive Behavior. Anesthesiology 2016, 124, 1136–1152. [Google Scholar] [CrossRef]
- Petitjean, H.; Pawlowski, S.A.; Fraine, S.L.; Sharif, B.; Hamad, D.; Fatima, T.; Berg, J.; Brown, C.M.; Jan, L.Y.; Ribeiro-da-Silva, A.; et al. Dorsal Horn Parvalbumin Neurons Are Gate-Keepers of Touch-Evoked Pain after Nerve Injury. Cell Rep. 2015, 13, 1246–1257. [Google Scholar] [CrossRef] [Green Version]
- Pham-Dang, N.; Descheemaeker, A.; Dallel, R.; Artola, A. Activation of medullary dorsal horn gamma isoform of protein kinase C interneurons is essential to the development of both static and dynamic facial mechanical allodynia. Eur. J. Neurosci. 2016, 43, 802–810. [Google Scholar] [CrossRef]
- Mermet-Joret, N.; Alba-Delgado, C.; Descheemaeker, A.; Gabrielli, F.; Dallel, R.; Antri, M. Postnatal development of inner lamina II interneurons of the rat medullary dorsal horn. Pain, 2021; in press. [Google Scholar] [CrossRef]
- Zhao, C.; Leitges, M.; Gereau, R.W.t. Isozyme-specific effects of protein kinase C in pain modulation. Anesthesiology 2011, 115, 1261–1270. [Google Scholar] [CrossRef] [Green Version]
- Abraira, V.E.; Kuehn, E.D.; Chirila, A.M.; Springel, M.W.; Toliver, A.A.; Zimmerman, A.L.; Orefice, L.L.; Boyle, K.A.; Bai, L.; Song, B.J.; et al. The Cellular and Synaptic Architecture of the Mechanosensory Dorsal Horn. Cell 2017, 168, 295–310. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Dong, H.; Gao, Y.; Gong, Y.; Ren, Y.; Gu, N.; Zhou, S.; Xia, N.; Sun, Y.Y.; Ji, R.R.; et al. A feed-forward spinal cord glycinergic neural circuit gates mechanical allodynia. J. Clin. Investig. 2013, 123, 4050–4062. [Google Scholar] [CrossRef] [Green Version]
- Neumann, S.; Braz, J.M.; Skinner, K.; Llewellyn-Smith, I.J.; Basbaum, A.I. Innocuous, not noxious, input activates PKCgamma interneurons of the spinal dorsal horn via myelinated afferent fibers. J. Neurosci. Off. J. Soc. Neurosci. 2008, 28, 7936–7944. [Google Scholar] [CrossRef] [PubMed]
- Peirs, C.; Patil, S.; Bouali-Benazzouz, R.; Artola, A.; Landry, M.; Dallel, R. Protein kinase C gamma interneurons in the rat medullary dorsal horn: Distribution and synaptic inputs to these neurons, and subcellular localization of the enzyme. J. Comp. Neurol. 2014, 522, 393–413. [Google Scholar] [CrossRef] [PubMed]
- Alba-Delgado, C.; Mountadem, S.; Mermet-Joret, N.; Monconduit, L.; Dallel, R.; Artola, A.; Antri, M. 5-HT2A Receptor-Induced Morphological Reorganization of PKCgamma-Expressing Interneurons Gates Inflammatory Mechanical Allodynia in Rat. J. Neurosci. Off. J. Soc. Neurosci. 2018, 38, 10489–10504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peirs, C.; Dallel, R.; Todd, A.J. Recent advances in our understanding of the organization of dorsal horn neuron populations and their contribution to cutaneous mechanical allodynia. J. Neural. Transm. 2020, 127, 505–525. [Google Scholar] [CrossRef] [Green Version]
- Boyle, K.A.; Gradwell, M.A.; Yasaka, T.; Dickie, A.C.; Polgar, E.; Ganley, R.P.; Orr, D.P.H.; Watanabe, M.; Abraira, V.E.; Kuehn, E.D.; et al. Defining a Spinal Microcircuit that Gates Myelinated Afferent Input: Implications for Tactile Allodynia. Cell Rep. 2019, 28, 526–540.e6. [Google Scholar] [CrossRef]
- Wang, Q.; Zhang, X.; He, X.; Du, S.; Jiang, Z.; Liu, P.; Qi, L.; Liang, C.; Gu, N.; Lu, Y. Synaptic Dynamics of the Feed-forward Inhibitory Circuitry Gating Mechanical Allodynia in Mice. Anesthesiology 2020, 132, 1212–1228. [Google Scholar] [CrossRef]
- Chery, N.; de Koninck, Y. Junctional versus extrajunctional glycine and GABA(A) receptor-mediated IPSCs in identified lamina I neurons of the adult rat spinal cord. J. Neurosci. Off. J. Soc. Neurosci. 1999, 19, 7342–7355. [Google Scholar] [CrossRef] [Green Version]
- Jonas, P.; Bischofberger, J.; Sandkuhler, J. Corelease of two fast neurotransmitters at a central synapse. Science 1998, 281, 419–424. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, J.A.; Berger, A.J. Cotransmission of GABA and glycine to brain stem motoneurons. J. Neurophysiol. 1999, 82, 1638–1641. [Google Scholar] [CrossRef] [Green Version]
- Todd, A.J.; Watt, C.; Spike, R.C.; Sieghart, W. Colocalization of GABA, glycine, and their receptors at synapses in the rat spinal cord. J. Neurosci. 1996, 16, 974–982. [Google Scholar] [CrossRef] [Green Version]
- Anderson, W.B.; Graham, B.A.; Beveridge, N.J.; Tooney, P.A.; Brichta, A.M.; Callister, R.J. Different forms of glycine- and GABA(A)-receptor mediated inhibitory synaptic transmission in mouse superficial and deep dorsal horn neurons. Mol. Pain 2009, 5, 65. [Google Scholar] [CrossRef] [Green Version]
- Grudt, T.J.; Henderson, G. Glycine and GABAA receptor-mediated synaptic transmission in rat substantia gelatinosa: Inhibition by mu-opioid and GABAB agonists. J. Physiol. 1998, 507, 473–483. [Google Scholar] [CrossRef]
- Inquimbert, P.; Rodeau, J.L.; Schlichter, R. Regional differences in the decay kinetics of GABA(A) receptor-mediated miniature IPSCs in the dorsal horn of the rat spinal cord are determined by mitochondrial transport of cholesterol. J. Neurosci. Off. J. Soc. Neurosci. 2008, 28, 3427–3437. [Google Scholar] [CrossRef] [Green Version]
- Keller, A.F.; Breton, J.D.; Schlichter, R.; Poisbeau, P. Production of 5alpha-reduced neurosteroids is developmentally regulated and shapes GABA(A) miniature IPSCs in lamina II of the spinal cord. J. Neurosci. Off. J. Soc. Neurosci. 2004, 24, 907–915. [Google Scholar] [CrossRef]
- Keller, A.F.; Coull, J.A.; Chery, N.; Poisbeau, P.; De Koninck, Y. Region-specific developmental specialization of GABA-glycine cosynapses in laminas I–II of the rat spinal dorsal horn. J. Neurosci. 2001, 21, 7871–7880. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, E.A.; Gentet, L.J.; Dempster, J.; Belelli, D. GABAA and glycine receptor-mediated transmission in rat lamina II neurones: Relevance to the analgesic actions of neuroactive steroids. J. Physiol. 2007, 583, 1021–1040. [Google Scholar] [CrossRef]
- Poisbeau, P.; Patte-Mensah, C.; Keller, A.F.; Barrot, M.; Breton, J.D.; Luis-Delgado, O.E.; Freund-Mercier, M.J.; Mensah-Nyagan, A.G.; Schlichter, R. Inflammatory pain upregulates spinal inhibition via endogenous neurosteroid production. J. Neurosci. 2005, 25, 11768–11776. [Google Scholar] [CrossRef] [Green Version]
- Zimmermann, M. Ethical guidelines for investigations of experimental pain in conscious animals. Pain 1983, 16, 109–110. [Google Scholar] [CrossRef]
- Alba-Delgado, C.; El Khoueiry, C.; Peirs, C.; Dallel, R.; Artola, A.; Antri, M. Subpopulations of PKCgamma interneurons within the medullary dorsal horn revealed by electrophysiologic and morphologic approach. Pain 2015, 156, 1714–1728. [Google Scholar] [CrossRef]
- Nagy, G.G.; Al-Ayyan, M.; Andrew, D.; Fukaya, M.; Watanabe, M.; Todd, A.J. Widespread expression of the AMPA receptor GluR2 subunit at glutamatergic synapses in the rat spinal cord and phosphorylation of GluR1 in response to noxious stimulation revealed with an antigen-unmasking method. J. Neurosci. 2004, 24, 5766–5777. [Google Scholar] [CrossRef] [Green Version]
- Wisden, W.; Gundlach, A.L.; Barnard, E.A.; Seeburg, P.H.; Hunt, S.P. Distribution of GABAA receptor subunit mRNAs in rat lumbar spinal cord. Mol. Brain Res. 1991, 10, 179–183. [Google Scholar] [CrossRef]
- Haring, M.; Zeisel, A.; Hochgerner, H.; Rinwa, P.; Jakobsson, J.E.T.; Lonnerberg, P.; La Manno, G.; Sharma, N.; Borgius, L.; Kiehn, O.; et al. Neuronal atlas of the dorsal horn defines its architecture and links sensory input to transcriptional cell types. Nat. Neurosci. 2018, 21, 869–880. [Google Scholar] [CrossRef] [PubMed]
- Sathyamurthy, A.; Johnson, K.R.; Matson, K.J.E.; Dobrott, C.I.; Li, L.; Ryba, A.R.; Bergman, T.B.; Kelly, M.C.; Kelley, M.W.; Levine, A.J. Massively Parallel Single Nucleus Transcriptional Profiling Defines Spinal Cord Neurons and Their Activity during Behavior. Cell Rep. 2018, 22, 2216–2225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorenzo, L.E.; Godin, A.G.; Ferrini, F.; Bachand, K.; Plasencia-Fernandez, I.; Labrecque, S.; Girard, A.A.; Boudreau, D.; Kianicka, I.; Gagnon, M.; et al. Enhancing neuronal chloride extrusion rescues alpha2/alpha3 GABAA-mediated analgesia in neuropathic pain. Nat. Commun. 2020, 11, 869. [Google Scholar] [CrossRef]
- Ralvenius, W.T.; Neumann, E.; Pagani, M.; Acuna, M.A.; Wildner, H.; Benke, D.; Fischer, N.; Rostaher, A.; Schwager, S.; Detmar, M.; et al. Itch suppression in mice and dogs by modulation of spinal alpha2 and alpha3GABAA receptors. Nat. Commun. 2018, 9, 3230. [Google Scholar] [CrossRef]
- Llewellyn-Smith, I.J.; Minson, J.B. Complete penetration of antibodies into vibratome sections after glutaraldehyde fixation and ethanol treatment: Light and electron microscopy for neuropeptides. J. Histochem. Cytochem. 1992, 40, 1741–1749. [Google Scholar] [CrossRef] [Green Version]
- Sardella, T.C.; Polgar, E.; Watanabe, M.; Todd, A.J. A quantitative study of neuronal nitric oxide synthase expression in laminae I-III of the rat spinal dorsal horn. Neuroscience 2011, 192, 708–720. [Google Scholar] [CrossRef] [Green Version]
- Kralic, J.E.; Korpi, E.R.; O’Buckley, T.K.; Homanics, G.E.; Morrow, A.L. Molecular and pharmacological characterization of GABA(A) receptor alpha1 subunit knockout mice. J. Pharmacol. Exp. Ther. 2002, 302, 1037–1045. [Google Scholar] [CrossRef] [Green Version]
- Schroder, S.; Hoch, W.; Becker, C.M.; Grenningloh, G.; Betz, H. Mapping of antigenic epitopes on the alpha 1 subunit of the inhibitory glycine receptor. Biochemistry 1991, 30, 42–47. [Google Scholar] [CrossRef]
- Zafra, F.; Aragon, C.; Olivares, L.; Danbolt, N.C.; Gimenez, C.; Storm-Mathisen, J. Glycine transporters are differentially expressed among CNS cells. J. Neurosci. 1995, 15, 3952–3969. [Google Scholar] [CrossRef]
- Chang, Y.C.; Gottlieb, D.I. Characterization of the proteins purified with monoclonal antibodies to glutamic acid decarboxylase. J. Neurosci. 1988, 8, 2123–2130. [Google Scholar] [CrossRef] [Green Version]
- Yoshimura, M.; Nishi, S. Primary afferent-evoked glycine- and GABA-mediated IPSPs in substantia gelatinosa neurones in the rat spinal cord in vitro. J. Physiol. 1995, 482, 29–38. [Google Scholar] [CrossRef]
- Salin, P.A.; Prince, D.A. Spontaneous GABAA receptor-mediated inhibitory currents in adult rat somatosensory cortex. J. Neurophysiol. 1996, 75, 1573–1588. [Google Scholar] [CrossRef]
- Hughes, B.A.; Bohnsack, J.P.; O’Buckley, T.K.; Herman, M.A.; Morrow, A.L. Chronic Ethanol Exposure and Withdrawal Impair Synaptic GABAA Receptor-Mediated Neurotransmission in Deep-Layer Prefrontal Cortex. Alcohol. Clin. Exp. Res. 2019, 43, 822–832. [Google Scholar] [CrossRef]
- Todd, A.J.; Spike, R.C.; Chong, D.; Neilson, M. The relationship between glycine and gephyrin in synapses of the rat spinal cord. Eur. J. Neurosci. 1995, 7, 1–11. [Google Scholar] [CrossRef]
- Usoskin, D.; Furlan, A.; Islam, S.; Abdo, H.; Lonnerberg, P.; Lou, D.; Hjerling-Leffler, J.; Haeggstrom, J.; Kharchenko, O.; Kharchenko, P.V.; et al. Unbiased classification of sensory neuron types by large-scale single-cell RNA sequencing. Nat. Neurosci. 2015, 18, 145–153. [Google Scholar] [CrossRef]
- Zeisel, A.; Hochgerner, H.; Lonnerberg, P.; Johnsson, A.; Memic, F.; van der Zwan, J.; Haring, M.; Braun, E.; Borm, L.E.; La Manno, G.; et al. Molecular Architecture of the Mouse Nervous System. Cell 2018, 174, 999–1014.e22. [Google Scholar] [CrossRef] [Green Version]
- Lorenzo, L.E.; Godin, A.G.; Wang, F.; St-Louis, M.; Carbonetto, S.; Wiseman, P.W.; Ribeiro-da-Silva, A.; De Koninck, Y. Gephyrin clusters are absent from small diameter primary afferent terminals despite the presence of GABA(A) receptors. J. Neurosci. 2014, 34, 8300–8317. [Google Scholar] [CrossRef] [Green Version]
- Bohlhalter, S.; Weinmann, O.; Mohler, H.; Fritschy, J.M. Laminar compartmentalization of GABAA-receptor subtypes in the spinal cord: An immunohistochemical study. J. Neurosci. 1996, 16, 283–297. [Google Scholar] [CrossRef] [Green Version]
- Waldvogel, H.J.; Baer, K.; Eady, E.; Allen, K.L.; Gilbert, R.T.; Mohler, H.; Rees, M.I.; Nicholson, L.F.; Faull, R.L. Differential localization of gamma-aminobutyric acid type A and glycine receptor subunits and gephyrin in the human pons, medulla oblongata and uppermost cervical segment of the spinal cord: An immunohistochemical study. J. Comp. Neurol. 2010, 518, 305–328. [Google Scholar] [CrossRef]
- Zeilhofer, H.U.; Wildner, H.; Yevenes, G.E. Fast synaptic inhibition in spinal sensory processing and pain control. Physiol. Rev. 2012, 92, 193–235. [Google Scholar] [CrossRef] [PubMed]
- Harvey, R.J.; Depner, U.B.; Wassle, H.; Ahmadi, S.; Heindl, C.; Reinold, H.; Smart, T.G.; Harvey, K.; Schutz, B.; Abo-Salem, O.M.; et al. GlyR alpha3: An essential target for spinal PGE2-mediated inflammatory pain sensitization. Science 2004, 304, 884–887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spike, R.C.; Todd, A.J.; Johnston, H.M. Coexistence of NADPH diaphorase with GABA, glycine, and acetylcholine in rat spinal cord. J. Comp. Neurol. 1993, 335, 320–333. [Google Scholar] [CrossRef] [PubMed]
- Todd, A.J.; Spike, R.C. The localization of classical transmitters and neuropeptides within neurons in laminae I-III of the mammalian spinal dorsal horn. Prog. Neurobiol. 1993, 41, 609–645. [Google Scholar] [CrossRef]
- Todd, A.J.; Sullivan, A.C. Light microscope study of the coexistence of GABA-like and glycine-like immunoreactivities in the spinal cord of the rat. J. Comp. Neurol. 1990, 296, 496–505. [Google Scholar] [CrossRef]
- Watanabe, M.; Fukaya, M.; Sakimura, K.; Manabe, T.; Mishina, M.; Inoue, Y. Selective scarcity of NMDA receptor channel subunits in the stratum lucidum (mossy fibre-recipient layer) of the mouse hippocampal CA3 subfield. Eur. J. Neurosci. 1998, 10, 478–487. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Figures | Analysis (Post-Hoc Test) | Factor Analyzed | F-Ratios | p-Values |
|---|---|---|---|---|
| Figure 1D | One-way ANOVA (Tukey) | IPSC type | IPSC type F(2,12) = 18.4 | <0.001 |
| Figure 1E | One-way ANOVA (Tukey) | IPSC type | IPSC type F(2,12) < 0.1 | 1.0 |
| Figure 1F | One-way ANOVA (Tukey) | IPSC type | IPSC type F(2,12) = 3.1 | 0.08 |
| Figure 2 | Two-way ANOVA (Tukey) | Cell type × IPSC type | Cell type F(1,54) < 0.1 | 1.0 |
| IPSC type F(2,54) = 8.2 | <0.001 | |||
| Interaction F(2,54) = 2.6 | 0.08 |
| Variables | Analysis (Post-Hoc Test) | Factor Analyzed | F-Ratios | p-Values |
|---|---|---|---|---|
| Decay τ of mIPSC (ms) | Two-way ANOVA (Tukey) | Cell type × IPSC type | Cell type F(1,24) = 0.4 | 0.5 |
| IPSC type F(2,24) = 42.4 | <0.001 | |||
| Interaction F(2,24) = 1.3 | 0.3 | |||
| Decay τ of sIPSC (ms) | Two-way ANOVA (Tukey) | Cell type × IPSC type | Cell type F(1,24) = 0.3 | 0.6 |
| IPSC type F(2,24) = 46.8 | <0.001 | |||
| Interaction F(2,24) = 0.1 | 0.5 | |||
| Rise τ of mIPSC (ms) | Two-way ANOVA (Tukey) | Cell type × IPSC type | Cell type F(1,24) = 2.5 | 0.1 |
| IPSC type F(2,24) < 0.1 | 1.0 | |||
| Interaction F(2,24) = 0.2 | 0.8 | |||
| Instantaneous frequency of mIPSC (Hz) | Two-way ANOVA (Tukey) | Cell type × IPSC type | Cell type F(1,24) = 6.6 | <0.05 |
| IPSC type F(2,24) = 0.2 | 0.8 | |||
| Interaction F(2,24) = 0.2 | 0.8 | |||
| Instantaneous frequency of sIPSC (Hz) | Two-way ANOVA (Tukey) | Cell type × IPSC type | Cell type F(1,24) < 0.1 | 0.8 |
| IPSC type F(2,24) = 0.5 | 0.6 | |||
| Interaction F(2,24) = 0.6 | 0.6 | |||
| Amplitude of mIPSC (pA) | Two-way ANOVA (Tukey) | Cell type × IPSC type | Cell type F(1,24) < 0.1 | 0.9 |
| IPSC type F(2,24) = 5.1 | <0.05 | |||
| Interaction F(2,24) = 0.3 | 0.8 | |||
| Amplitude of sIPSC (pA) | Two-way ANOVA (Tukey) | Cell type × IPSC type | Cell type F(1,24) = 0.4 | 0.5 |
| IPSC type F(2,24) = 0.8 | 0.4 | |||
| Interaction F(2,24) < 0.1 | 0.9 |
| PKCγ+ | PKCγ− | |||
|---|---|---|---|---|
| mIPSC (5) | sIPSC (5) | mIPSC (5) | sIPSC (5) | |
| Decay τ (ms) | ||||
| Fast monoexpo | 8.9 ± 0.9 a,b | 10.0 ± 1.0 | 11.7 ± 0.6 | 11.8 ± 1.4 |
| Slow monoexpo | 31.7 ± 1.3 c | 31.9 ± 2.0 | 30.1 ± 0.7 | 30.4 ± 2.4 |
| Biexpo | 20.6 ± 4.3 | 19.6 ± 2.7 | 16.0 ± 2.9 | 16.6 ± 2.6 |
| Rise τ (ms) | ||||
| Fast monoexpo | 3.6 ± 0.6 | ~ | 2.4 ± 0.6 | ~ |
| Slow monoexpo | 3.4 ± 0.7 | ~ | 3.0 ± 0.6 | ~ |
| Biexpo | 3.6 ± 0.5 | ~ | 2.6 ± 0.9 | ~ |
| Instantaneous frequency (Hz) | ||||
| Fast monoexpo | 0.2 ± 0.1 | 0.5 ± 0.2 | 1.1 ± 0.5 | 0.3 ± 0.1 |
| Slow monoexpo | 0.3 ± 0.1 | 0.2 ± 0.1 | 0.7 ± 0.5 | 0.3 ± 0.1 |
| Biexpo | 0.3 ± 0.1 | 0.3 ± 0.1 | 1.0 ± 0.4 | 0.6 ± 0.1 |
| Amplitude (pA) | ||||
| Fast monoexpo | 39.9 ± 5.7 | 37.1 ± 13.2 | 48.5 ± 7.2 | 33.1 ± 6.0 |
| Slow monoexpo | 49.5 ± 9.8 | 37.5 ± 8.9 | 46.9 ± 5.9 | 35.4 ± 8.1 |
| Biexpo | 72.4 ± 11.9 | 51.7 ± 12.2 | 69.5 ± 11.7 | 42.7 ± 11.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El Khoueiry, C.; Alba-Delgado, C.; Antri, M.; Gutierrez-Mecinas, M.; Todd, A.J.; Artola, A.; Dallel, R. GABAA and Glycine Receptor-Mediated Inhibitory Synaptic Transmission onto Adult Rat Lamina IIi PKCγ-Interneurons: Pharmacological but Not Anatomical Specialization. Cells 2022, 11, 1356. https://doi.org/10.3390/cells11081356
El Khoueiry C, Alba-Delgado C, Antri M, Gutierrez-Mecinas M, Todd AJ, Artola A, Dallel R. GABAA and Glycine Receptor-Mediated Inhibitory Synaptic Transmission onto Adult Rat Lamina IIi PKCγ-Interneurons: Pharmacological but Not Anatomical Specialization. Cells. 2022; 11(8):1356. https://doi.org/10.3390/cells11081356
Chicago/Turabian StyleEl Khoueiry, Corinne, Cristina Alba-Delgado, Myriam Antri, Maria Gutierrez-Mecinas, Andrew J. Todd, Alain Artola, and Radhouane Dallel. 2022. "GABAA and Glycine Receptor-Mediated Inhibitory Synaptic Transmission onto Adult Rat Lamina IIi PKCγ-Interneurons: Pharmacological but Not Anatomical Specialization" Cells 11, no. 8: 1356. https://doi.org/10.3390/cells11081356