
A Conceptual Model for Detecting Small-Scale Forest Disturbances Based on Ecosystem Morphological Traits

 , ,
, ,
Abstract
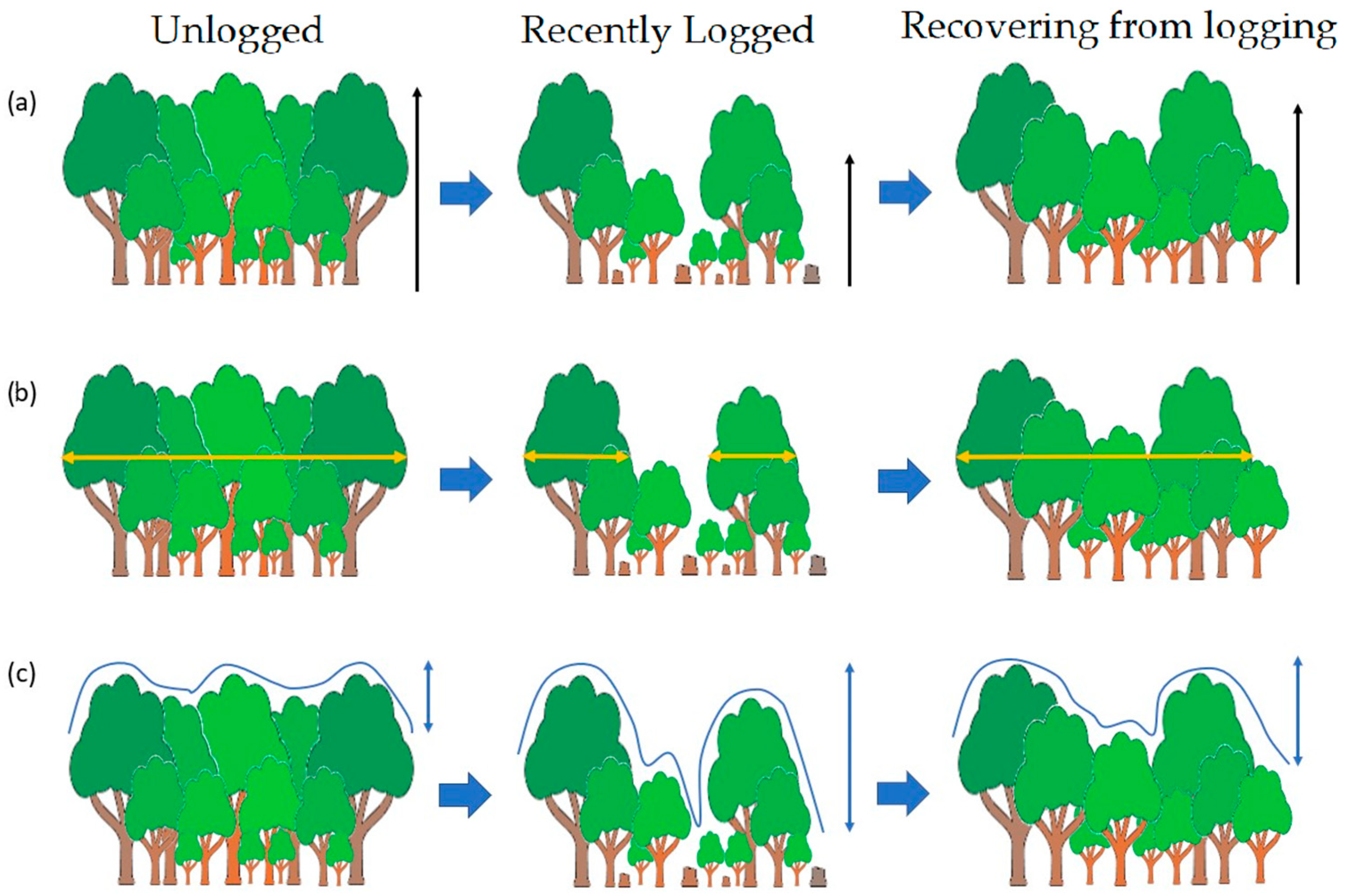
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Field Data
2.3. LiDAR Data Acquisition and Processing
2.4. AGB Modelling
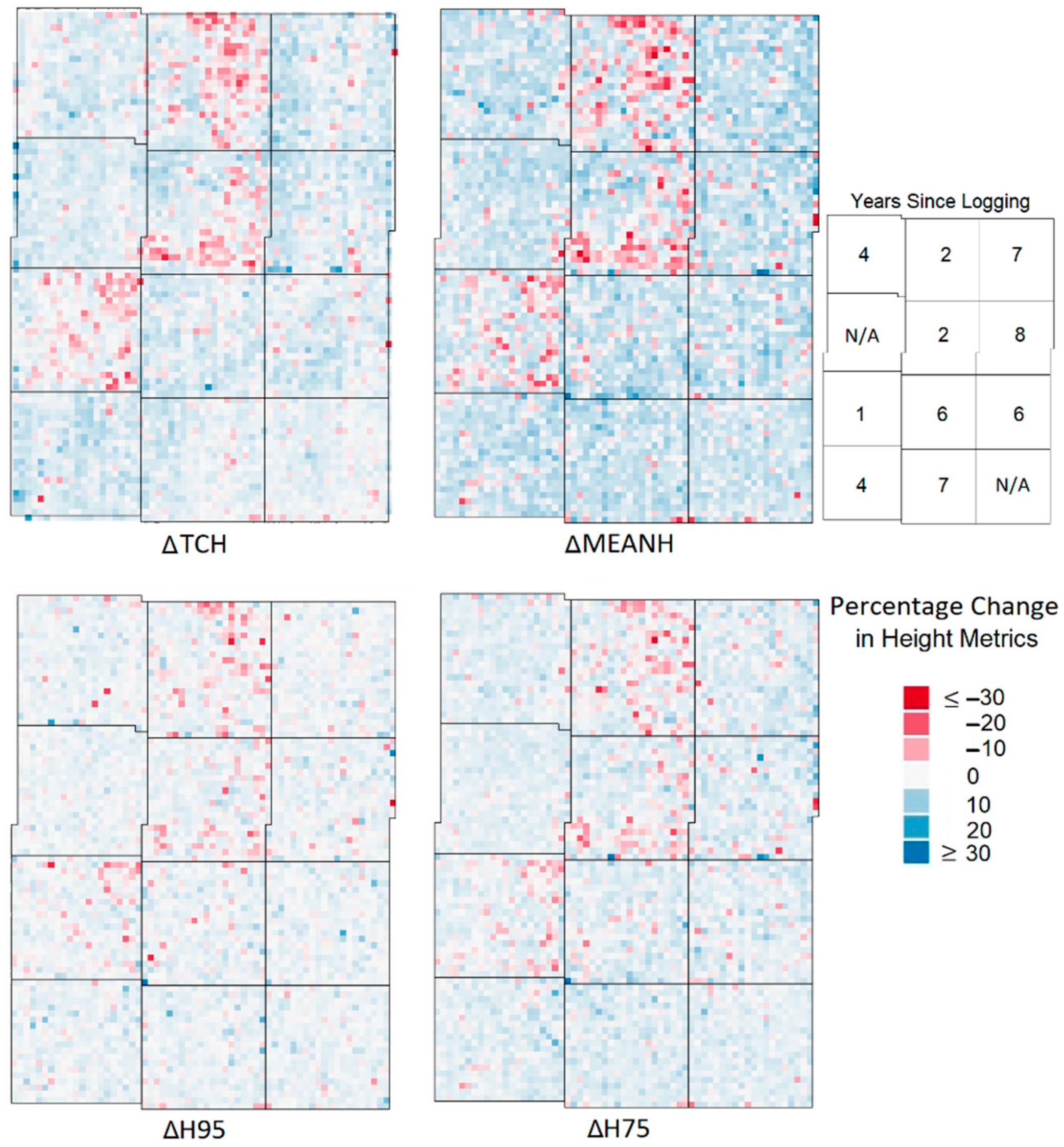
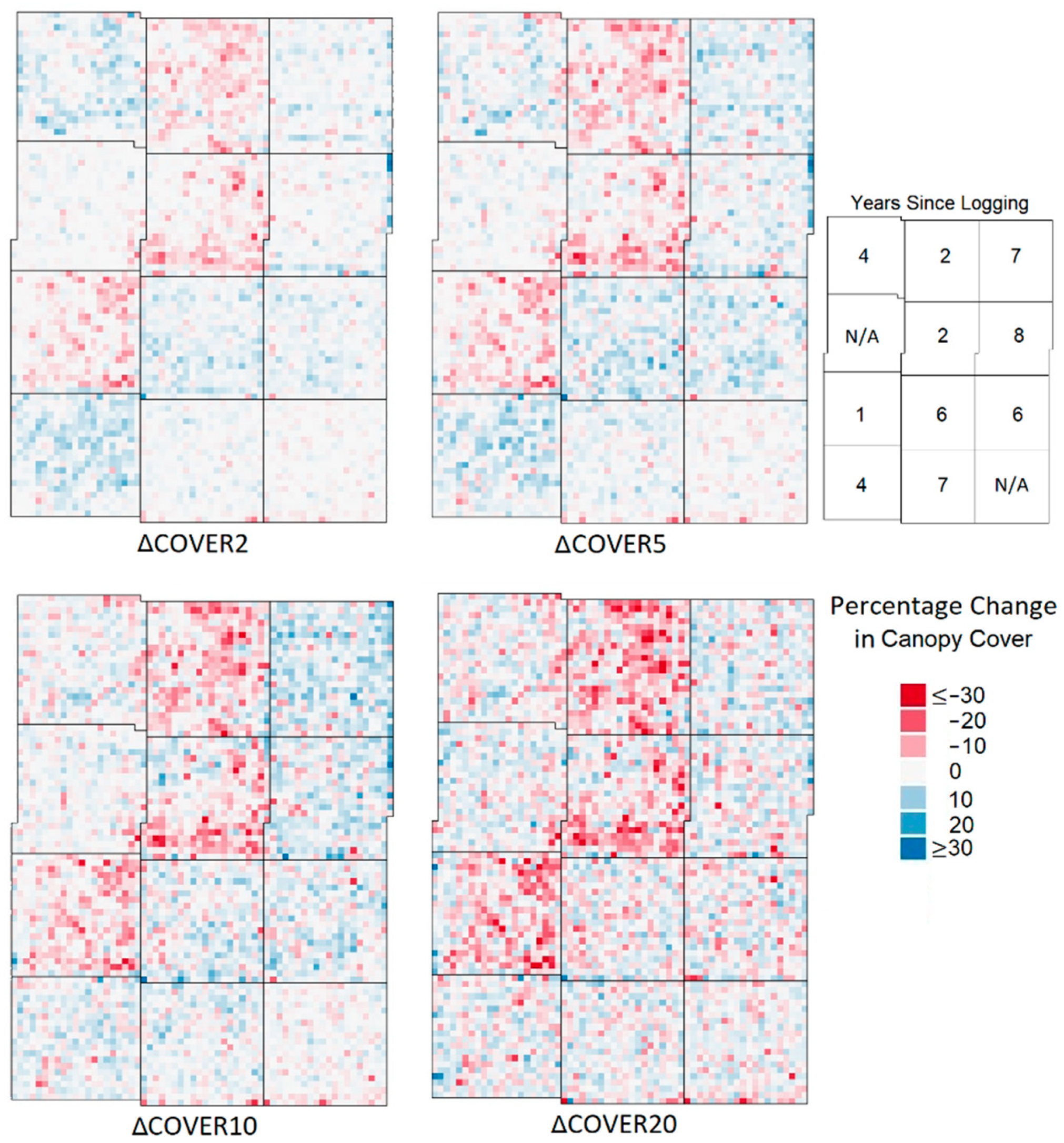
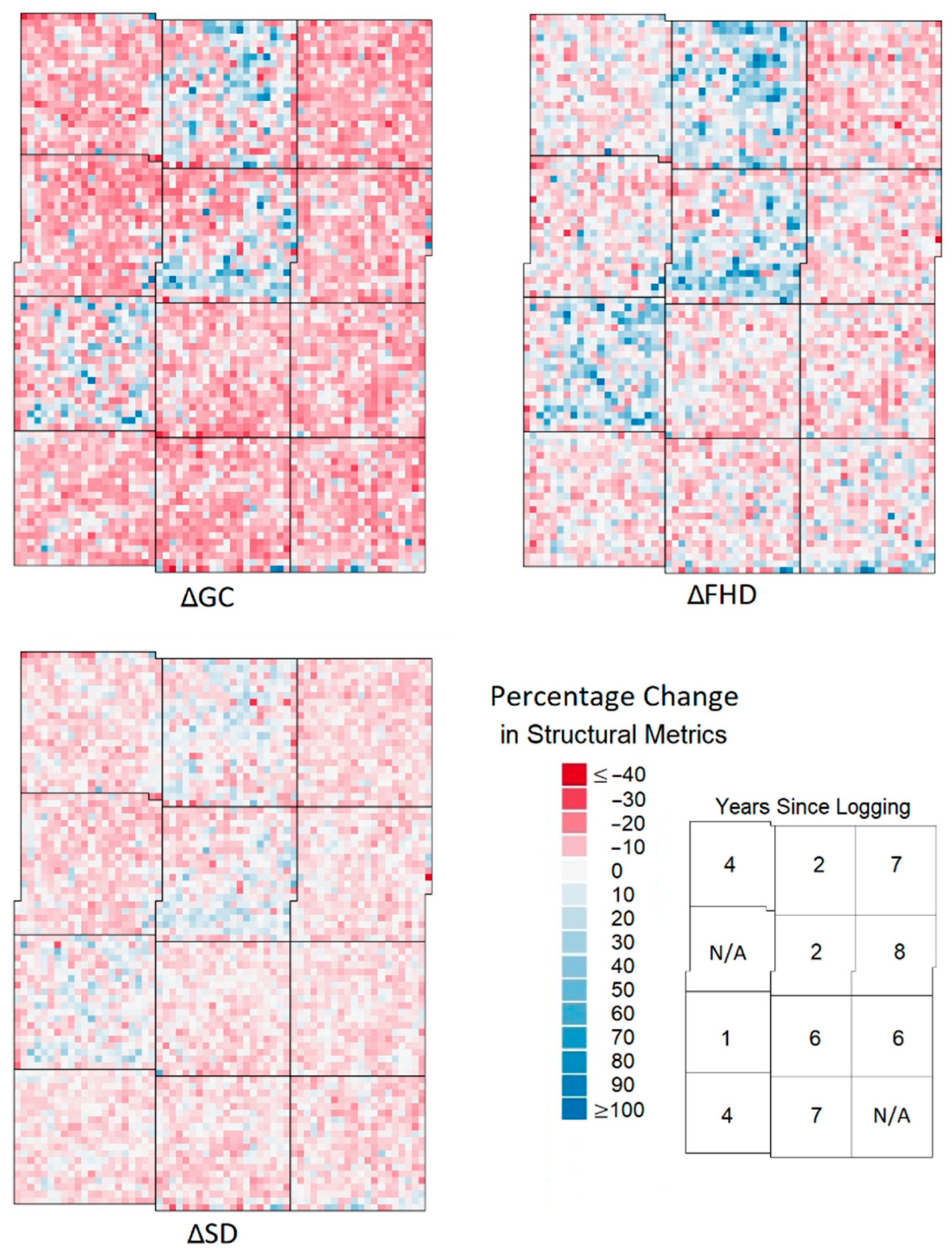
- Maps of percentage change in metrics (denoted as Δ in front of each metric). From the maps of change in the metrics it was possible to identify the metrics which showed the greatest percentage change in areas of recent and historical selective logging, and thus sensitivity to small-scale disturbance, and this was one factor that was used to select LiDAR proxies. One-tailed Wilcoxon rank tests, for data grouped by the logging year, were used to assess the significance of the increase or decrease in a given metric compared to the baseline defined by the unlogged areas.
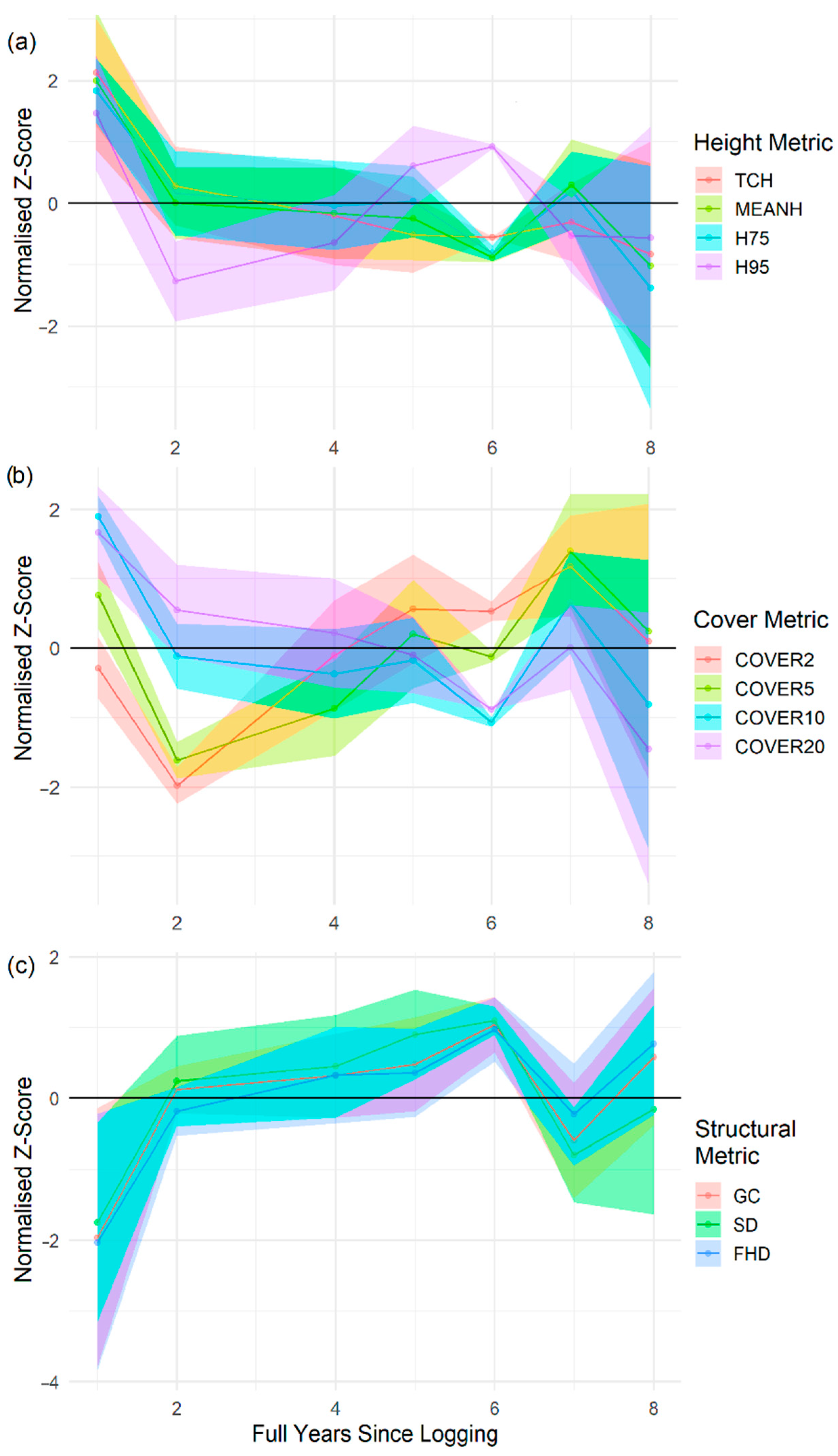
- Graphs of temporal dynamics of metrics in the years following logging. The graphs of normalised z-scores (in which the mean is equal to 0) were used to identify which metrics showed the greatest change in the years after selective logging and the graphs also highlighted how long it took metrics to stabilise and thus for how long after logging metrics were sensitive to the impacts of small-scale disturbance.
- Correlation plots and Pearson correlation coefficients. Plots of the correlation of change in metrics, with trendlines and inset correlation values. These identified relationships between metrics that allowed for informed decisions to be made with regards to the use of metrics which may explain the same variance.
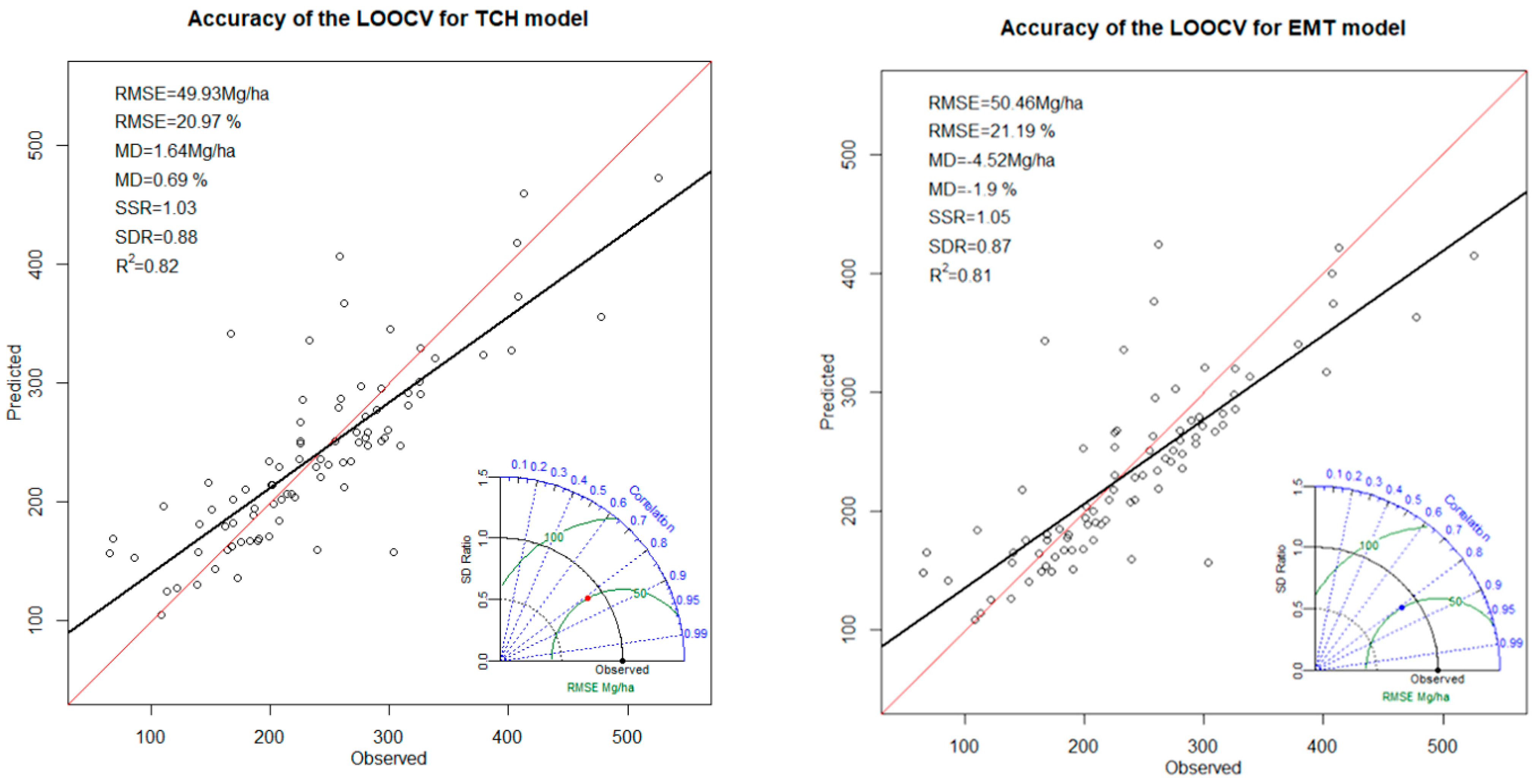
2.5. Accuracy Assessment
- (1)
- The precision of predictions given as the absolute and relative root mean squared error (RMSE) of predictions against the observed:where was the predicted sum of squares through LOOCV:
- (2)
- The degree of under- or over-prediction as the mean difference (MD) of the predictions minus the observed:
- (3)
- The agreement between observed and predicted, evaluated by the coefficient of determination (Valbuena et al., 2019):where was the sum of squared differences of each observation from :
- (4)
- The degree of overfitting, comparing the sums of squares obtained with (“cv”) and without (“fit”) cross-validation (Valbuena et al. 2017), calculated as the sum of squares ratio (SSR):where was the predicted sum of squares without LOOCV:
- (5)
- The model’s capacity to match the AGB variability originally observed. As the standard deviation ratio (SDR):where was the sum of squared differences of each observation from the mean predicted AGB ():
- (a)
- Correlation, denoted by the azimuthal angle (blue radial dashed lines and arc) which represents the Pearson correlation coefficient, evaluating similarity in patterns of distribution of the predicted and observed, which is also a measure of the similarity of the means. The Pearson correlation coefficient is a normalised measure of the linear correlation between two datasets, in this case the predictions of a leave-one-out model and the observed dataset [79]. Valbuena et al. (2019) [80] pointed out that only the squared correlation gives values comparable to the coefficient of determination in Equation (3), but correlation is added here as implemented in the package stats.
- (b)
- RMSE, proportional to the distance of a point from the reference for observed data, on the x-axis (which is shown by green arcs).
- (c)
- The standard deviation ratio of the cross-validation predictions to the observed is proportional to the radial distance from the origin (black arcs). The standard deviation ratio reflects how well the predictions reflect the variance of observed data.
3. Results
3.1. Vegetation Height
3.2. Vegetation Cover
3.3. Structural Complexity
3.4. TCH Model versus EMT Model
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brienen, R.J.; Phillips, O.L.; Feldpausch, T.R.; Gloor, E.; Baker, T.R.; Lloyd, J.; Lopez-Gonzalez, G.; Monteagudo-Mendoza, A.; Malhi, Y.; Lewis, S.L.; et al. Long-term decline of the Amazon carbon sink. Nature 2015, 519, 344–348. [Google Scholar] [CrossRef] [Green Version]
- Phillips, O.L.; Brienen, R.J.W.; The RAINFOR Collaboration. Carbon uptake by mature Amazon forests has mitigated Amazon nations’ carbon emissions. Carbon Balance Manag. 2017, 12, 1. [Google Scholar] [CrossRef] [Green Version]
- Reboredo, F. Socio-economic, environmental, and governance impacts of illegal logging. Environ. Syst. Decis. 2013, 33, 295–304. [Google Scholar] [CrossRef]
- Condé, M.; Higuchi, N.; Adriano, J.N. Illegal Selective Logging and Forest Fires in the Northern Brazilian Amazon. Forests 2019, 10, 61. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Skutsch, M.M.; Paneque-Gálvez, J.; Ghilardi, A. Remote sensing of forest degradation: A review. Environ. Res. Lett. 2020, 15, 103001. [Google Scholar] [CrossRef]
- Assunção, J.; Gandour, C.; Rocha, R. DETERing Deforestation in the Amazon: Environmental Monitoring and Law Enforcement. Climate Policy Initiative, Núcleo de Avaliação de Políticas Climáticas, Pontifica Universidade Católica (PUC), Rio de Janeiro, 2017. p. 36. Available online: http://climatepolicyinitiative.org/wp-content/uploads/2013/05/DETERring-Deforestation-in-the-Brazilian-Amazon-Environmental-Monitoring-and-Law-Enforcement-Technical-Paper.pdf (accessed on 16 June 2021).
- Coops, N.C.; Tompaski, P.; Nijland, W.; Rickbeil, G.J.; Nielsen, S.E.; Bater, C.W.; Stadt, J.J. A forest structure habitat index based on airborne laser scanning data. Ecol. Ind. 2016, 67, 346–357. [Google Scholar] [CrossRef]
- Schneider, F.D.; Morsdorf, F.; Schmid, B.; Petchey, O.L.; Hueni, A.; Schimel, D.S.; Schaepman, M.E. Mapping functional diversity from remotely sensed morphological and physiological forest traits. Nat. Commun. 2017, 8, 1441. [Google Scholar] [CrossRef] [Green Version]
- Fahey, R.T.; Atkins, J.W.; Gough, C.M.; Hardiman, B.S.; Nave, L.E.; Tallant, J.M.; Nadehoffer, K.J.; Vogel, C.; Scheuermann, C.M.; Stuart-Haëntjens, E.; et al. Defining a spectrum of integrative trait-based vegetation canopy structural types. Ecol. Lett. 2019, 22, 2049–2059. [Google Scholar] [CrossRef]
- Valbuena, R.; O’Connor, B.; Zellweger, F.; Simonson, W.; Vihervaara, P.; Maltamo, M.; Silva, C.A.; Almeida, D.R.A.; Danks, F.; Morsdorf, F.; et al. Standardizing Ecosystem Morphological Traits from 3D Information Sources. Trends Ecol. Evol. 2020, 35, 656–667. [Google Scholar] [CrossRef]
- Lefsky, M.A.; Hudak, A.T.; Cohen, W.B.; Acker, S.A. Patterns of covariance between forest stand and canopy structure in the Pacific Northwest. Remote. Sens. Environ. 2005, 95, 517–531. [Google Scholar] [CrossRef] [Green Version]
- Asner, G.P.; Mascaro, J.; Muller-Landau, H.C.; Vieilledent, G.; Vaudry, R.; Rasamoelina, M.; Hall, J.S.; Van Breugel, M. A universal airborne LiDAR approach for tropical forest carbon mapping. Oecologia 2012, 168, 1147–1160. [Google Scholar] [CrossRef] [PubMed]
- Locks, C.J.; Matricardi, E.A.T. Estimation of impacts of selective logging in the Amazon using LIDAR data. For. Sci. 2019, 29, 481–495. [Google Scholar] [CrossRef] [Green Version]
- Lefsky, M.A.; Cohen, W.B.; Parker, G.G.; Harding, D.J. Lidar Remote Sensing for Ecosystem Studies: Lidar, an emerging remote sensing technology that directly measures the three-dimensional distribution of plant canopies, can accurately estimate vegetation structural attributes and should be of particular interest to forest, landscape, and global ecologists. BioScience 2002, 52, 19–30. [Google Scholar] [CrossRef]
- Naesset, E. Predicting forest stand characteristics with airborne scanning laser using a practical two-stage procedure and field data. Remote. Sens. Environ. 2002, 80, 88–99. [Google Scholar] [CrossRef]
- Asner, G.P.; Rudel, T.K.; Aide, T.M.; Defries, R.; Emerson, R. A Contemporary Assessment of Change in Humid Tropical Forests. Conserv. Biol. 2009, 23, 1386–1395. [Google Scholar] [CrossRef]
- Zolkos, S.; Goetz, S.J.; Dubayah, R. A meta-analysis of terrestrial aboveground biomass estimation using lidar remote sensing. Remote Sens. Environ. 2013, 128, 289–298. [Google Scholar] [CrossRef]
- Valbuena, R.; Hernando, A.; Manzanera, J.A.; Görgens, E.B.; Almeida, D.R.A.; Mauro, F.; García-Abril, A.; Coomes, D.A. Enhancing of Accuracy Assessment for Forest Above-Ground Biomass Estimates Obtained from Remote Sensing via Hypothesis Testing and Overfitting Evaluation. Ecol. Model. 2017, 366, 15–26. [Google Scholar] [CrossRef] [Green Version]
- Houghton, R.A. Aboveground Forest Biomass and the Global Carbon Balance. Global Change Biology 2005, 11, 945–958. [Google Scholar] [CrossRef]
- Görgens, E.B.; Nunes, M.H.; Jackson, T.; Coomes, D.A.; Keller, M.; Reis, C.R.; Valbuena, R.; Rosette, J.; Almeida, D.R.A.; Gimenez, B.; et al. Resource Availability and Disturbance Shape Maximum Tree Height across the Amazon. Glob. Chang. Biol. 2020, 27, 177–189. [Google Scholar] [CrossRef]
- Clark, M.L.; Roberts, D.A.; Ewel, J.J.; Clark, D.B. Estimation of tropical rain forest aboveground biomass with small-footprint lidar and hyperspectral sensors. Remote Sens. Environ. 2011, 115, 2931–2942. [Google Scholar] [CrossRef]
- Asner, G.P.; Mascaro, J. Mapping tropical forest carbon: Calibrating plot estimates to a simple LiDAR metric. Remote Sens. Environ. 2014, 140, 614–624. [Google Scholar] [CrossRef]
- Latifi, H.; Heurich, M.; Hartig, F.; Müller, J.; Krzystek, P.; Jehl, H.; Dech, S. Estimating over and understorey canopy density of temperate mixed stands by airborne LiDAR data. Forestry 2015, 89, 69–81. [Google Scholar] [CrossRef] [Green Version]
- Latifi, H.; Fassnacht, F.E.; Muller, J.; Tharani, A.; Dech, S.; Heurich, M. Forest inventories by LiDAR data: A comparison of single tree segmentation and metric-based methods for inventories of a heterogeneous temperate forest. Int. J. Appl. Earth Obs. Geoinf. 2015, 42, 162–174. [Google Scholar] [CrossRef]
- Longo, M.; Keller, M.; dos-Santos, M.N.; Leitold, V.; Pinagé, E.R.; Baccini, A.; Saatchi, S.; Nogueira, E.M.; Batistella, M.; Morton, D.C. Aboveground biomass variability across intact and degraded forests in the Brazilian Amazon. Glob. Biogeochem. Cycles 2016, 30, 1639–1660. [Google Scholar] [CrossRef] [Green Version]
- Jucker, T.; Asner, G.P.; Dalponte, M.; Brodrick, P.G.; Philipson, C.D.; Vaughn, N.R.; Teh, Y.A.; Brelsford, C.; Burslem, D.F.R.P.; Deere, N.J.; et al. Estimating aboveground carbon density and its uncertainty in Borneo’s structurally complex tropical forests using airborne laser scanning. Biogeosciences 2018, 15, 3811–3830. [Google Scholar] [CrossRef]
- Almeida, D.R.A.; Stark, S.C.; Schietti, J.; Camargo, J.L.C.; Amazonas, N.T.; Görgens, E.B.; Rosa, D.M.; Smith, M.N.; Valbuena, R.; Saleska, S.; et al. Persistent Effects of Fragmentation on Tropical Rainforest Canopy Structure after 20 Years of Isolation. Ecol. Appl. 2019, 29, e01952. [Google Scholar] [CrossRef] [PubMed]
- Almeida, D.R.A.; Broadbent, E.; Wendt, A.; Wilkinson, B.; Papa, D.A.; Chazdon, R.; Meli, P.; Görgens, E.B.; Silva, C.A.; Stark, S.C.; et al. Detecting Successional Changes in Tropical Forest Structure Using Gatoreye Drone-Borne Lidar. Biotropica 2020, 52, 1155–1167. [Google Scholar] [CrossRef]
- Beland, M.; Parker, G.; Sparrow, B. Harding, D.; Chasmer, L.; Phinn, S.; Antonarakis, A.; Strahler, A. On promoting the use of lidar systems in forest ecosystem research. For. Ecol. Manag. 2019, 450, 117484. [Google Scholar] [CrossRef]
- Hernando, A.; Puerto, L.; Mola-Yudego, B.; Manzanera, J.A.; García-Abril, A.; Maltamo, M.; Valbuena, R. Estimation of Forest Biomass Components through Airborne LiDAR and Multispectral Sensors. iForests—Biogeosciences For. 2019, 12, 207–213. [Google Scholar] [CrossRef] [Green Version]
- Adnan, S.; Maltamo, M.; Mehtätalo, L.; Ammaturo, R.N.; Packalen, P.; Valbuena, R. Determining Maximum Entropy in 3D Remote Sensing Height Distributions and Using it to Improve Aboveground Biomass Modelling via Stratification. Remote. Sens. Environ. 2021, 260, 112464. [Google Scholar] [CrossRef]
- Costa, M.A.T.; Silva, C.A.; Broadbent, E.N.; Leite, R.V.; Mohan, M.; Liesenberg, V.; Stoddart, J.; Amaral, C.H.; Almeida, D.R.A.; da Silva, A.L.; et al. Beyond Trees: Mapping Total Aboveground Biomass Density in the Brazilian Savanna Using High-Density UAV-Lidar Data. For. Ecol. Manag. 2021, 491, 119155. [Google Scholar] [CrossRef]
- Rex, F.E.; Silva, C.A.; Dalla Corte, A.P.; Klauberg, C.; Mohan, M.; Cardil, A.; Silva, V.S.; Almeida, D.R.A.; Garcia, M.; Broadbent, E.N.; et al. Comparison of Statistical Modelling Approaches for Estimating Tropical Forest Aboveground Biomass Stock and Reporting Their Changes in Low-Intensity Logging Areas Using Multi-Temporal LiDAR Data. Remote. Sensing. 2020, 12, 1498. [Google Scholar] [CrossRef]
- Nunes, M.H.; Jucker, T.; Riutta, T.; Svátek, M.; Kvasnica, J.; Rejžek, M.; Matula, R.; Majalap, N.; Ewers, R.M.; Swinfield, T.; et al. Recovery of logged forest fragments in a human-modified tropical landscape during the 2015–2016 El Niño. Nat. Commun 2021, 12, 1526. [Google Scholar] [CrossRef] [PubMed]
- d’Oliveira, M.V.; Reutebuch, S.E.; McGaughey, R.; Andersen, H. Estimating forest biomass and identifying low-intensity logging areas using airborne scanning lidar in Antimary State Forest, Acre State, Western Brazilian Amazon. Remote. Sens. Environ. 2012, 124, 479–491. [Google Scholar] [CrossRef]
- Meyer, V.; Saatchi, S.S.; Chave, J.; Dalling, J.W.; Bohlman, S.; Fricker, G.A.; Robinson, C.; Neumann, M.; Hubbell, S. Detecting tropical forest biomass dynamics from repeated airborne lidar measurements. Biogeoscience 2013, 10, 5421–5438. [Google Scholar] [CrossRef] [Green Version]
- Kankare, V.; Vastaranta, M.; Holopainen, M.; Räty, M.; Yu, X.; Hyyppä, J.; Hyyppä, H.; Alho, P.; Viitala, R. Retrieval of Forest Aboveground Biomass and Stem Volume with Airborne Scanning LiDAR. Remote Sens. 2013, 5, 2257–2274. [Google Scholar] [CrossRef] [Green Version]
- Marchesan, J.; Alba, E.; Schuh, M.; Favarin, J.; Pereira, R. Aboveground Biomass Estimation in A Tropical Forest with Selective Logging Using Random Forest And Lidar Data. Floresta 2020, 50, 1873. [Google Scholar] [CrossRef]
- Valbuena, R.; Maltamo, M.; Mehtätalo, L.; Packalen, P. Key Structural Features of Boreal Forests May Be Detected Directly Using L-Moments from Airborne Lidar Data. Remote. Sens. Environ. 2017, 194, 437–446. [Google Scholar] [CrossRef] [Green Version]
- Garcia, M.; Saatchi, S.; Ustin, S.; Balzter, H. Modelling forest canopy height by integrating airborne LiDAR samples with satellite Radar and multispectral imagery. Int. J. Appl. Earth Obs. Geoinf. 2018, 66, 159–173. [Google Scholar] [CrossRef]
- Bouvier, M.; Durrieu, S.; Fournier, R.A.; Renaud, J.P. Generalizing predictive models of forest inventory attributes using an area-based approach with airborne LiDAR data. Remote. Sens. Environ. 2015, 156, 322–334. [Google Scholar] [CrossRef]
- Wang, Y.; Lehtomäki, M.; Liang, X.; Pyörälä, J.; Kukko, A.; Jaakkola, A.; Liu, J.; Feng, Z.; Chen, R.; Hyyppä, J. Is field-measured tree height as reliable as believed–a comparison study of tree height estimates from field measurement, airborne laser scanning and terrestrial laser scanning in a boreal forest. ISPRS J. Photogramm. Remote Sens. 2019, 147, 132–145. [Google Scholar] [CrossRef]
- Lefsky, M.A.; Harding, D.; Cohen, W.B.; Parker, G.; Shugart, H.H. Surface lidar remote sensing of basal area and biomass in deciduous forests of eastern Maryland, USA. Remote. Sens. Environ. 1999, 67, 83–98. [Google Scholar] [CrossRef]
- Silva, C.A.; Valbuena, R.; Pinagé, E.R.; Mohan, M.; de Almeida, D.R.; North Broadbent, E.; Jaafar, W.S.; de Almeida Papa, D.; Cardil, A.; Klauberg, C. ForestGapR: An r Package for forest gap analysis from canopy height models. Methods Ecol. Evol. 2019, 10, 1347–1356. [Google Scholar] [CrossRef] [Green Version]
- Hinsley, S.A.; Hill, R.A.; Fuller, R.J.; Bellamy, P.E.; Rothery, P. Bird species distributions cross woodland canopy structure gradients. Commun. Ecol. 2009, 10, 99–110. [Google Scholar] [CrossRef]
- Kane, V.R.; McGaughey, R.J.; Bakker, J.D.; Gersonde, R.F.; Lutz, J.A.; Franklin, J.F. Comparisons between field- and LiDAR-based measures of stand structural complexity. Can. J. For. Res. 2010, 40, 761–773. [Google Scholar] [CrossRef]
- Zellweger, F.; Baltensweiler, A.; Ginzler, C.; Roth, T.; Braunisch, V.; Bugmann, H.; Bollmann, K. Environmental predictors of species richness in forest landscapes: Abiotic factors versus vegetation structure. J. Biogeogr. 2016, 43, 1080–1090. [Google Scholar] [CrossRef]
- Bater, C.; Wulder, M.A.; Coops, N.C.; Nelson, R.; Hilker, T. Stability of sample-based scanning-LiDAR-derived vegetation metrics for forest monitoring. IEEE Trans. Geosci. Remote. Sens. 2011, 49, 2385–2392. [Google Scholar] [CrossRef]
- Nelson, R.; Krabill, W.; Tonelli, J. Estimating Forest Biomass and Volume Using Airborne Laser Data. Remote. Sens. Environ. 1988, 24, 247–267. [Google Scholar] [CrossRef]
- Morsdorf, F.; Kötz, B.; Meier, E.; Itten, K.I.; Allgöwer, B. Estimation of LAI and fractional cover from small footprint airborne laser scanning data based on gap fraction. Remote Sens Environ. 2006, 104, 50–61. [Google Scholar] [CrossRef]
- Solberg, S. Mapping gap fraction, LAI and defoliation using various ALS penetration variables. Int. J. Remote Sens. 2010, 31, 1227–1244. [Google Scholar] [CrossRef]
- Korhonen, L.; Korpela, I.; Heiskanen, J.; Maltamo, M. Airborne discrete-return LiDAR data in the estimation of vertical canopy cover, angular canopy closure and leaf area index. Remote. Sens Environ. 2011, 115, 1065–1080. [Google Scholar] [CrossRef]
- Gorgens, E.B.; Valbuena, R.; Rodriguez, L.C.E. A method for optimizing height threshold when computing airborne laser scanning metrics. Photogram. Eng. Remote Sens. 2017, 83, 343–350. [Google Scholar] [CrossRef]
- Wedeux, B.M.M.; Coomes, D.A. Landscape-scale changes in forest canopy structure across a partially logged tropical peat swamp. Biogeosciences 2015, 12, 6707–6719. [Google Scholar] [CrossRef] [Green Version]
- Clawges, R.; Vierling, K.; Vierling, L.; Rowell, E. The use of airborne lidar to assess avian species diversity, density, and occurrence in a pine/aspen forest. Remote. Sens. Environ. 2008, 112, 2064–2073. [Google Scholar] [CrossRef]
- Bergen, K.M.; Goetz, S.J.; Dubayah, R.O.; Henebry, G.M.; Hunsaker, C.T.; Imhoff, M.L.; Nelson, R.F.; Parker, G.G.; Radeloff, V.C. Remote sensing of vegetation 3-D structure for biodiversity and habitat: Review and implications for lidar and radar spaceborne missions. J. Geophys. Res. 2009, 114, G00E06. [Google Scholar] [CrossRef] [Green Version]
- Valbuena, R.; Packalen, P.; Martín-Fernández, S.; Maltamo, M. Diversity and equitability ordering profiles applied to the study of forest structure. For. Ecol. Manag. 2012, 276, 185–195. [Google Scholar] [CrossRef] [Green Version]
- Valbuena, R.; Maltamo, M.; Martín-Fernández, S.; Packalen, P.; Pascual, C.; Nabuurs, G.J. Patterns of covariance between airborne laser scanning metrics and Lorenz curve descriptors of tree size inequality. Can. J. Remote Sens. 2013, 39, S18–S31. [Google Scholar] [CrossRef]
- Pinagé, E.R.; Keller, M.; Duffy, P.; Longo, M.; dos-Santos, M.N.; Morton, D.C. Long-Term Impacts of Selective Logging on Amazon Forest Dynamics from Multi-Temporal Airborne LiDAR. Remote Sens. 2019, 11, 709. [Google Scholar] [CrossRef] [Green Version]
- Silva, C.A.; Hudak, A.T.; Vierling, L.A.; Klauberg, C.; Garcia, M.; Ferraz, A.; Keller, M.; Eitel, J.; Saatchi, S. Impacts of Airborne Lidar Pulse Density on Estimating Biomass Stocks and Changes in a Selectively Logged Tropical Forest. Remote Sens. 2017, 9, 1068. [Google Scholar] [CrossRef] [Green Version]
- Görgens, E.B.; Mund, J.P.; Cremer, T.; de Conto, T.; Krause, S.; Valbuena, R.; Rodríguez, L.C. Automated Operational Logging Plan Considering Multi-Criteria Optimization. Comput. Electron. Agric. 2020, 170, 105253. [Google Scholar] [CrossRef]
- Rex, F.E.; Corte, A.P.D.; Machado, S.D.A.; Silva, C.A.; Sanquetta, C.R. Estimating Above-Ground Biomass of Araucaria angustifolia (Bertol.) Kuntze Using LiDAR Data. Floresta E Ambiente 2019, 26. [Google Scholar] [CrossRef]
- Chave, J.; Réjou-Méchain, M.; Búrquez, A.; Chidumayo, E.; Colgan, M.S.; Delitti, W.B.; Duque, A.; Eid, T.; Fearnside, P.M.; Goodman, R.C.; et al. Improved allometric models to estimate the aboveground biomass of tropical trees. Glob. Chang. Biol. 2014, 20, 3177–3190. [Google Scholar] [CrossRef]
- McGaughey, R.J. FUSION/LDV: Software for LiDAR Data Analysis and Visualization; Version 3.80; USA Department of Agriculture Forest Service, Pacific Northwest Research Station, University of Washington: Seattle, WA, USA, 2019. [Google Scholar]
- Isenburg, M. LAStools—Efficient LiDAR Processing Software; Version 191018; Unlicensed; 2019. [Google Scholar]
- Roussel, J.R.; Auty, D.; Coops, N.C.; Tompalski, P.; Goodbody, T.R.H.; Sánchez Meador, A.; Bourdon, J.F.; De Boissieu, F.; Achim, A. lidR: An R package for analysis of Airborne Laser Scanning (ALS) data. Remote. Sens. Environ. 2020, 251, 112061. [Google Scholar] [CrossRef]
- McArthur, R.H.; McArthur, J.W. On bird species diversity. Ecology 1961, 42, 594–598. [Google Scholar] [CrossRef]
- R Core Team R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. 2021. Available online: https://www.R-project.org (accessed on 17 January 2021).
- QGIS.org QGIS Geographic Information System. QGIS Association. 2021. Available online: http://www.qgis.org (accessed on 16 May 2021).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Baskerville, G.L. Use of Logarithmic Regression in the Estimation of Plant Biomass. Can. J. For. Res. 1972, 2, 49–53. [Google Scholar] [CrossRef]
- Enquist, B.J.; Brown, J.H.; West, G.B. Allometric scaling of plant energetics and population density. Nature 1998, 395, 163–165. [Google Scholar] [CrossRef]
- Chave, J.; Andalo, C.; Brown, S.; Cairns, M.A.; Chamber, J.Q.; Eamus, D.; Folster, H.; Fromard, F.; Higuchi, N.; Kira, T.; et al. Tree allometry and improved estimation of carbon stocks and balance in tropical forests. Oecologia 2005, 145, 87–99. [Google Scholar] [CrossRef] [PubMed]
- Niklas, K.J. A phyletic perspective on the allometry of plant biomass-partitioning patterns and functionally equivalent organ categories. New Phytol. 2006, 171, 27–40. [Google Scholar] [CrossRef]
- Lipovetsky, S. How good is best? Multivariate case of Ehrenberg-Weisberg analysis of residual errors in competing regressions. J. Mod. Appl. Stat. Methods 2013, 12, 14. [Google Scholar] [CrossRef] [Green Version]
- Pineiro, G.; Perelman, S.; Guerschman, J.P.; Paruelo, J.M. How to evaluate models: Observed vs. predicted or predicted vs. observed? Ecol. Model. 2008, 216, 316–322. [Google Scholar] [CrossRef]
- Taylor, K.E. Summarizing multiple aspects of model performance in a single diagram. J. Geophys. Res. 2001, 106, 7183–7192. [Google Scholar] [CrossRef]
- Lemon, J. Plotrix: A package in the red light district of R. R-News 2006, 6, 8–12. [Google Scholar]
- Freedman, D.; Pisani, R.; Purves, R. Statistics (International Student Edition), 4th ed.; WW Norton & Company: New York, NY, USA, 2007. [Google Scholar]
- Hethcoat, M.G.; Edwards, D.P.; Carreiras, J.M.B.; Bryant, R.G.; França, F.M.; Quegan, S. A Machine Learning Approach to Map Tropical Selective Logging. Remote. Sens. Environ. 2019, 221, 569–582. [Google Scholar] [CrossRef]
- Su, H.; Shen, W.; Wang, J.; Ali, A.; Li, M. Machine learning and geostatistical approaches for estimating aboveground biomass in Chinese subtropical forests. For. Ecosyst. 2020, 7, 1–20. [Google Scholar] [CrossRef]
- St-Onge, B.; Vega, C.; Fournier, R.A.; Hu, Y. Mapping canopy height using a combination of digital stereo-photogrammetry and lidar. Int. J. Remote. Sens. 2008, 29, 3343–3364. [Google Scholar] [CrossRef]
- Kent, R.; Lindsell, J.; Laurin, G.; Valentini, R.; Coomes, D. Airborne LiDAR detects selectively logged tropical forest even in an advanced stage of recovery. Remote. Sens. 2015, 7, 8348–8367. [Google Scholar] [CrossRef] [Green Version]
- Valbuena, R.; Vauhkonen, J.; Packalen, P.; Pitkänen, J.; Maltamo, M. Comparison of airborne laser scanning methods for estimating forest structure indicators based on Lorenz curves. ISPRS J. Photogramm. Remote. Sens. 2014, 95, 23–33. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| EMT | Metrics | References | |
|---|---|---|---|
| Vegetation height | TCH | Lefsky et al., 1999 [43]; Asner and Mascaro, 2014 [22]; Fahey et al., 2019 [9]; Almeida et al., 2019 [27]; Silva et al., 2019 [44] | |
| MEANH | Næsset et al., 2002 [15]; Hinsley et al., 2009 [45]; Kane et al., 2010 [46]; Zellweger et al., 2016 [47]; Fahey et al., 2019 [9]; Rex et al., 2020 [33] | ||
| MODEH | Næsset et al., 2002 [15]; Fahey et al., 2019 [9]; Rex et al., 2020 [33] | ||
| MEDIANH | Næsset et al., 2002 [15]; Rex et al., 2020 [33] | ||
| Percentiles of height (H95, H75 etc.) | Næsset et al., 2002 [15]; Kane et al., 2010 [46]; Bater et al., 2011 [48]; Zellweger et al., 2016 [47] | ||
| Vegetation cover | Leaf area index/plant area index | COVER2 COVER5 COVER10 COVER20 | Nelson et al., 1988 [49]; Næsset et al., 2002 [15]; Morsdorf et al., 2006 [50]; Solberg, 2010 [51]; Korhonen et al., 2011 [52]; Görgens et al. 2017 [53]; Schneider et al., 2017 [8] |
| Gap fraction | Wedeux & Coomes, 2015 [54]; Jucker et al., 2018; Silva et al., 2019 [44] | ||
| Structural complexity of vegetation | Entropy | FHD | Clawges et al., 2008 [55]; Bergen et al., 2009 [56]; Valbuena et al. 2012 [57]; Zellweger et al., 2016 [47]; Schneider et al., 2017 [8] |
| Variability | SD | Kane et al., 2010 [46]; Bouvier et al., 2015 [41]; Coops et al., 2016 [7]; Zellweger et al., 2016 [47] | |
| GC | Kane et al., 2010 [46]; Valbuena et al., 2013; 2017b; 2020 [10,39,58]; Adnan et al., 2021 [31] | ||
| Adj.R2 | RMSE (Mg/ha) | RMSE (%) | MD (Mg/ha) | MD (%) | SSR | SDR | ||
|---|---|---|---|---|---|---|---|---|
| Height | a * × TCH * | 0.82 | 49.93 | 20.97 | 1.64 | 0.69 | 1.03 | 0.88 |
| MEANH * | 0.82 | 50.66 | 21.28 | −7.89 | 3.31 | 1.01 | 0.69 | |
| H95 * | 0.60 | 69.54 | 29.20 | −11.24 | −4.72 | 1.01 | 0.58 | |
| a * × H75 * | 0.56 | 56.73 | 23.83 | 1.02 | 0.43 | 0.91 | 0.76 | |
| Cover | COVER2 * | 0.01 | 87.77 | 36.86 | −15.84 | −6.65 | 1.03 | 0.10 |
| COVER5 * | 0.21 | 85.67 | 35.98 | −15.53 | −6.52 | 1.04 | 0.15 | |
| a * × COVER10 * | 0.50 | 74.72 | 31.38 | 1.41 | 0.59 | 1.02 | 0.52 | |
| COVER20 * | 0.75 | 57.54 | 24.16 | −5.85 | −2.46 | 1.01 | 0.75 | |
| Structural Complexity | a * × GC * | 0.13 | 80.79 | 33.93 | 1.92 | 0.81 | 1.02 | 0.46 |
| a * × FHD * | 0.37 | 70.31 | 29.53 | 2.64 | 1.11 | 1.05 | 0.80 | |
| a * × SD * | 0.07 | 83.06 | 34.88 | 1.62 | 0.68 | 1.03 | 0.32 | |
| Bi-variate models | ||||||||
| TCH * × COVER2 * | 0.82 | 50.07 | 21.03 | −5.47 | −2.30 | 1.03 | 0.83 | |
| TCH * × COVER5 * | 0.81 | 50.96 | 21.40 | −6.15 | −2.58 | 1.03 | 0.80 | |
| TCH * × COVER10 * | 0.80 | 52.95 | 22.24 | −7.79 | −3.27 | 1.03 | 0.72 | |
| TCH * × COVER20 * | 0.80 | 54.73 | 22.98 | −8.66 | −3.64 | 1.03 | 0.62 | |
| Tri-variate models | ||||||||
| TCH * × COVER2 * × GC * | 0.81 | 50.46 | 21.19 | −4.52 | −1.90 | 1.05 | 0.87 | |
| TCH * × COVER2 * × SD * | 0.81 | 51.17 | 21.49 | −4.71 | −1.98 | 1.05 | 0.86 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stoddart, J.; de Almeida, D.R.A.; Silva, C.A.; Görgens, E.B.; Keller, M.; Valbuena, R. A Conceptual Model for Detecting Small-Scale Forest Disturbances Based on Ecosystem Morphological Traits. Remote Sens. 2022, 14, 933. https://doi.org/10.3390/rs14040933
Stoddart J, de Almeida DRA, Silva CA, Görgens EB, Keller M, Valbuena R. A Conceptual Model for Detecting Small-Scale Forest Disturbances Based on Ecosystem Morphological Traits. Remote Sensing. 2022; 14(4):933. https://doi.org/10.3390/rs14040933
Chicago/Turabian StyleStoddart, Jaz, Danilo Roberti Alves de Almeida, Carlos Alberto Silva, Eric Bastos Görgens, Michael Keller, and Ruben Valbuena. 2022. "A Conceptual Model for Detecting Small-Scale Forest Disturbances Based on Ecosystem Morphological Traits" Remote Sensing 14, no. 4: 933. https://doi.org/10.3390/rs14040933