

Insights into the Relative Abundance, Life History, and Ecology of Oceanic Sharks in the Eastern Bahamas

 , , , , , , ,
, , , , , , ,
Abstract
:1. Introduction
2. Material and Methods
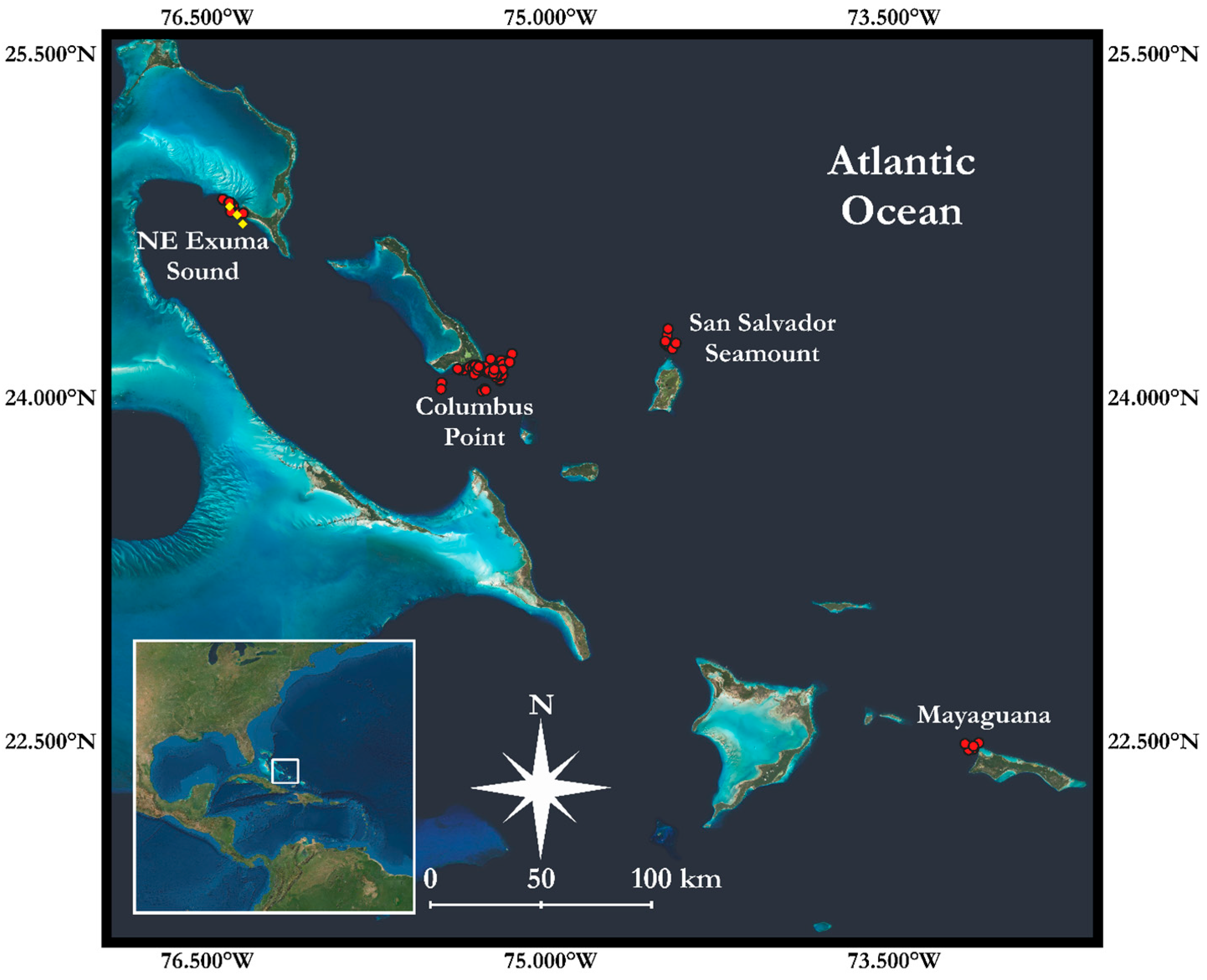
2.1. Study Area
2.2. Pelagic Longline Surveys
2.3. Targeted Baiting and Opportunistic Sightings
2.4. Shark Capture and Workup
2.5. Data Analysis
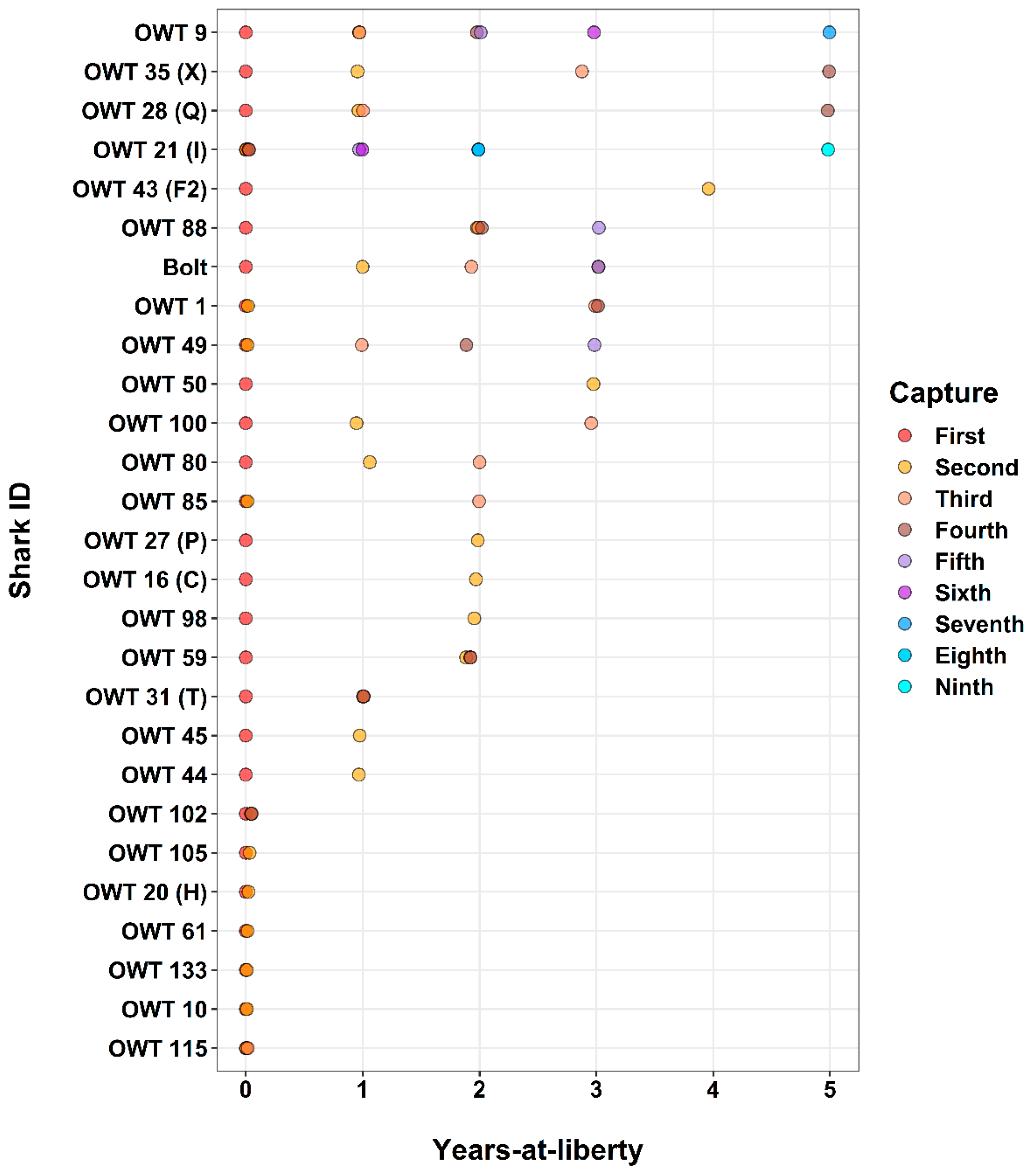
3. Results
3.1. North-Eastern Exuma Sound
3.1.1. Pelagic Longline Survey: Effort and Catch
3.1.2. Baiting and Other Techniques: Effort and Catch
3.1.3. Multispecies Associations
3.2. Columbus Point, Cat Island
3.2.1. Baiting: Effort and Catch
3.2.2. Multispecies Associations
3.3. San Salvador Seamount and Mayaguana
3.4. All Sites: Species-Specific Catch Characteristics
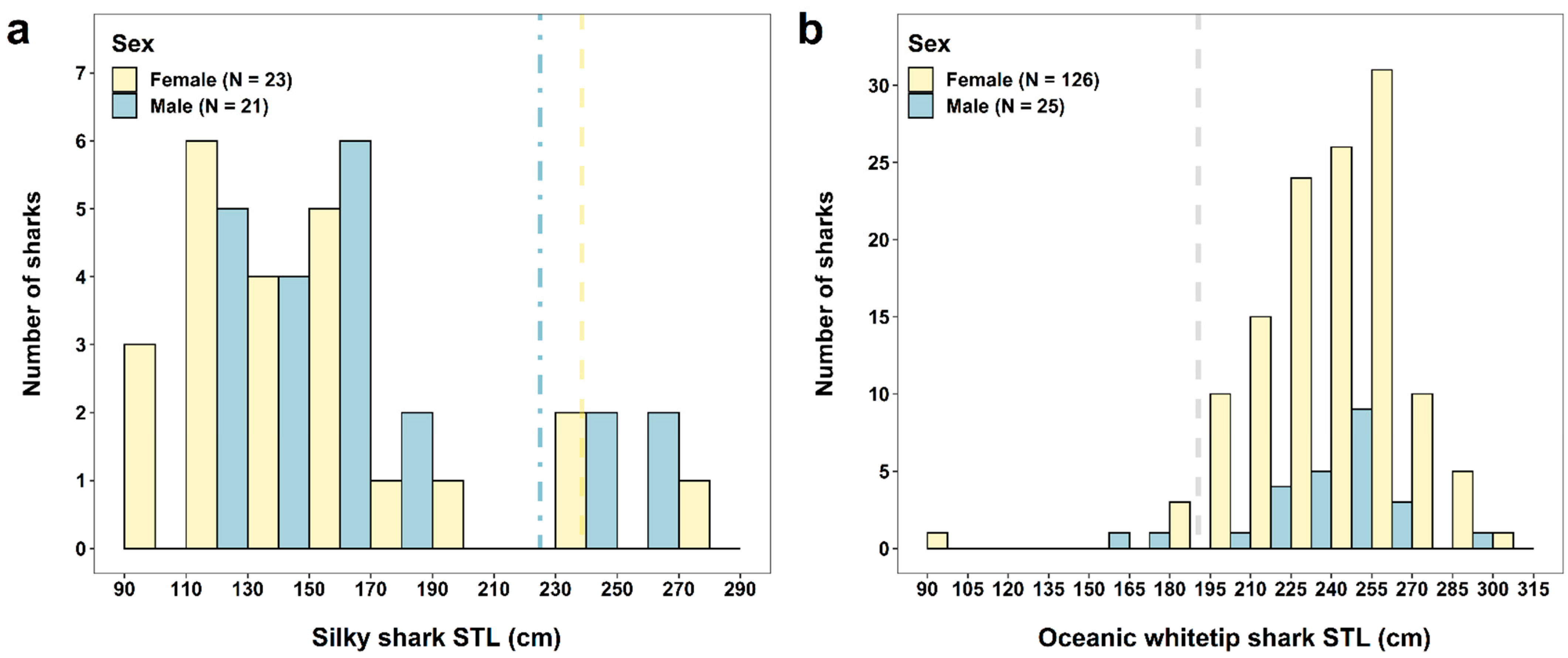
3.4.1. Silky Shark
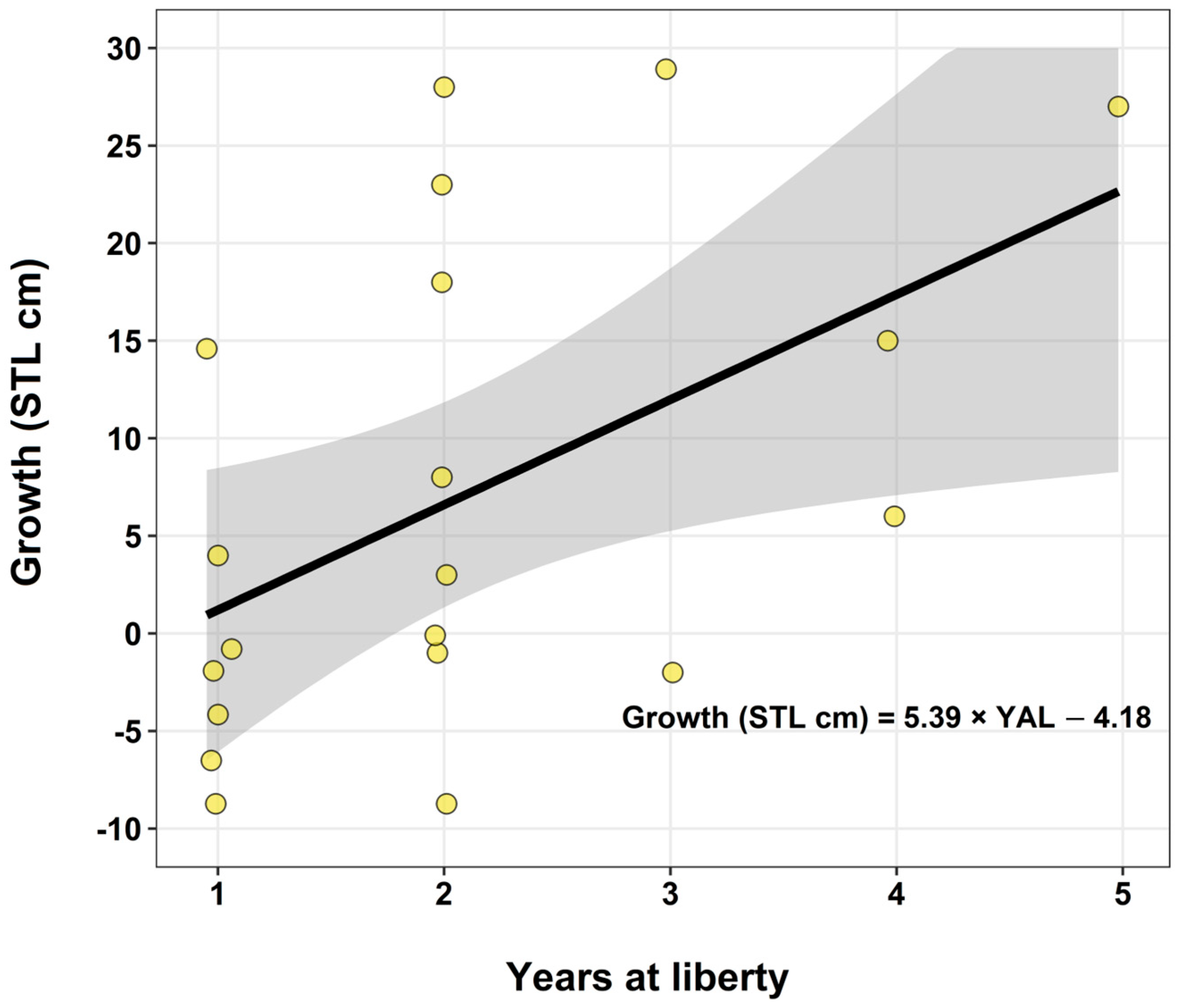
3.4.2. Oceanic Whitetip Shark
4. Discussion
4.1. Variable Species-Specific Conservation Value of Sites in the Eastern Bahamas
4.2. Multispecies Associations
4.3. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ruiz-Abierno, A.; Márquez-Farías, J.F.; Rojas-Corzo, A.; Miller, V.; Angulo-Valdés, J.A.; Hueter, R.E. Seasonal abundance and size structure of sharks taken in the pelagic longline fishery off northwestern Cuba. Mar. Coast. Fish. 2021, 13, 289–305. [Google Scholar] [CrossRef]
- Tagliafico, A.; Rangel, M.; Ehemann, N.; Rago, N.; Broadhurst, M. Reproductive aspects of seven threatened shark species captured by artisanal fisheries in the southern Caribbean Sea. Reg. Stud. Mar. Sci. 2021, 42, 101646. [Google Scholar] [CrossRef]
- Tavares, R.; Arocha, F. Species diversity, relative abundance and length structure of oceanic sharks caught by the Venezuelan longline fishery in the Caribbean Sea and western-Central Atlantic. Zootec. Trop. 2008, 26, 489–503. [Google Scholar]
- Cortes, E.; Brown, C.A.; Beerkircher, L.R. Relative abundance of pelagic sharks in the Western North Atlantic Ocean, including the Gulf of Mexico and Caribbean Sea. Gulf Caribb. Res. 2007, 19, 37–52. [Google Scholar] [CrossRef]
- Baum, J.K.; Blanchard, W. Inferring shark population trends from generalized linear mixed models of pelagic longline catch and effort data. Fish. Res. 2010, 102, 229–239. [Google Scholar] [CrossRef]
- Pacoureau, N.; Rigby, C.L.; Kyne, P.M.; Sherley, R.B.; Winker, H.; Carlson, J.K.; Fordham, S.V.; Barreto, R.; Fernando, D.; Francis, M.P.; et al. Half a century of global decline in oceanic sharks and rays. Nature 2021, 589, 567–571. [Google Scholar] [CrossRef] [PubMed]
- Cortés, E.; Arocha, F.; Beerkircher, L.; Carvalho, F.; Domingo, A.; Heupel, M.; Holtzhausen, H.; Santos, M.N.; Ribera, M.; Simpfendorfer, C. Ecological risk assessment of pelagic sharks caught in Atlantic pelagic longline fisheries. Aquat. Living Resour. 2009, 23, 25–34. [Google Scholar] [CrossRef]
- Stevens, J.D. Epipelagic oceanic elasmobranchs. In Sharks and Their Relatives II: Biodiversity, Adaptative Physiology and Conservation; Carrier, J.C., Musick, J.A., Heithaus, M.R., Eds.; CRC Press: Boca Raton, FL, USA, 2010; pp. 3–36. [Google Scholar]
- Garibaldi, L. The FAO global capture production database: A six-decade effort to catch the trend. Mar. Policy 2012, 36, 760–768. [Google Scholar] [CrossRef]
- ICCAT. Report of the 2019 Standing Committee on Research and Statistics (SCRS). Madrid, Spain, 30 September–4 October 2019. 2019. Available online: https://www.iccat.int/Documents/Meetings/Docs/2019/REPORTS/2019_SCRS_ENG.pdf (accessed on 9 October 2023).
- Juan-Jordá, M.J.; Murua, H.; Arrizabalaga, H.; Dulvy, N.K.; Restrepo, V. Report card on ecosystem-based fisheries management in tuna regional fisheries management organizations. Fish Fish. 2017, 19, 321–339. [Google Scholar] [CrossRef]
- Secor, D.H. The Unit Stock Concept: Bounded Fish and Fisheries. In Stock Identification Methods: Applications in Fishery Science; Cadrin, S.X., Kerr, L.A., Mariana, S., Eds.; Elsevier Science & Technology: San Diego, CA, USA, 2013. [Google Scholar]
- NOAA. Stock Assessment and Fishery Evaluation Report Atlantic Highly Migratory Species 2021; U.S. Department of Commerce, National Oceanic and Atmospheric Administration, National Marine Fisheries Service. 2021. Available online: https://media.fisheries.noaa.gov/2022-03/SAFE%20Report%202021%20Final_0.pdf (accessed on 9 October 2023).
- Young, C.N.; Carlson, J.K. The biology and conservation status of the oceanic whitetip shark (Carcharhinus longimanus) and future directions for recovery. Rev. Fish Biol. Fish. 2020, 30, 293–312. [Google Scholar] [CrossRef]
- Rigby, C.L.; Barreto, R.; Carlson, J.; Fernando, D.; Fordham, S.; Francis, M.P.; Herman, K.; Jabado, R.W.; Liu, K.M.; Marshall, A.; et al. Carcharhinus longimanus. The IUCN Red List of Threatened Species 2019. e.T39374A2911619. Available online: https://www.iucnredlist.org/species/39374/2911619 (accessed on 16 February 2022).
- Rigby, C.L.; Sherman, C.S.; Chin, A.; Simpfendorfer, C. Carcharhinus falciformis (Amended Version of 2017 Assessment). The IUCN Red List of Threatened Species 2021. 2021. e.T39370A205782570. Available online: https://www.iucnredlist.org/species/39370/205782570 (accessed on 16 May 2022).
- Ward-Paige, C.A.; Keith, D.M.; Worm, B.; Lotze, H.K. Recovery potential and conservation options for elasmobranchs. J. Fish Biol. 2012, 80, 1844–1869. [Google Scholar] [CrossRef]
- Chapman, D.D.; Feldheim, K.A.; Papastamatiou, Y.P.; Hueter, R.E. There and back again: A review of residency and return migrations in sharks, with implications for population structure and management. Annu. Rev. Mar. Sci. 2015, 7, 547–570. [Google Scholar] [CrossRef] [PubMed]
- Hyde, C.A.; di Sciara, G.N.; Sorrentino, L.; Boyd, C.; Finucci, B.; Fowler, S.L.; Kyne, P.M.; Leurs, G.; Simpfendorfer, C.A.; Tetley, M.J.; et al. Putting sharks on the map: A global standard for improving shark area-based conservation. Front. Mar. Sci. 2022, 9, 1–16. [Google Scholar] [CrossRef]
- Compagno, L.J. Pelagic elasmobranch diversity. In Sharks of the Open Ocean: Biology, Fisheries and Conservation, Book 15; Camhi, M.D., Pikitch, E.K., Babcock, E.A., Eds.; Blackwell Publishing: Oxford, UK, 2008; pp. 14–23. [Google Scholar]
- Madigan, D.; Brooks, E.; Bond, M.; Gelsleichter, J.; Howey, L.; Abercrombie, D.; Brooks, A.; Chapman, D. Diet shift and site-fidelity of oceanic whitetip sharks Carcharhinus longimanus along the Great Bahama Bank. Mar. Ecol. Prog. Ser. 2015, 529, 185–197. [Google Scholar] [CrossRef]
- Talwar, B.; Bond, M.; Williams, S.; Brooks, E.; Chapman, D.; Howey, L.; Knotek, R.; Gelsleichter, J. Reproductive timing and putative mating behavior of the oceanic whitetip shark Carcharhinus longimanus in the eastern Bahamas. Endanger. Species Res. 2023, 50, 181–194. [Google Scholar] [CrossRef]
- Howey-Jordan, L.A.; Brooks, E.J.; Abercrombie, D.L.; Jordan, L.K.B.; Brooks, A.; Williams, S.; Gospodarczyk, E.; Chapman, D.D. Complex movements, philopatry and expanded depth range of a severely threatened pelagic shark, the oceanic whitetip (Carcharhinus longimanus) in the Western North Atlantic. PLoS ONE 2013, 8, e56588. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.R.; Johnson, R.H.; Waldrop, L.G. Responses in Bahamian sharks and groupers to low-frequency, pulsed sounds. Bull. South. Calif. Acad. Sci. 1969, 68, 131–137. [Google Scholar]
- Myrberg, A.A., Jr.; Ha, S.J.; Walewski, S.; Banbury, J.C. Effectiveness of acoustic signals in attracting epipelagic sharks to an underwater sound source. Bull. Mar. Sci. 1972, 22, 926–949. [Google Scholar]
- Hueter, R.E.; Tyminski, J.P.; Amargós, F.P.; Morris, J.J.; Abierno, A.R.; Valdés, J.A.A.; Fernández, N.L. Movements of three female silky sharks (Carcharhinus falciformis) as tracked by satellite-linked tags off the Caribbean coast of Cuba. Bull. Mar. Sci. 2018, 94, 345–358. [Google Scholar] [CrossRef]
- Lezama-Ochoa, N.; Murua, H.; Chust, G.; Van Loon, E.; Ruiz, J.; Hall, M.; Chavance, P.; De Molina, A.D.; Villarino, E. Present and Future Potential Habitat Distribution of Carcharhinus falciformis and Canthidermis maculata By-Catch Species in the Tropical Tuna Purse-Seine Fishery under Climate Change. Front. Mar. Sci. 2016, 3, 1–16. [Google Scholar] [CrossRef]
- Morato, T.; Hoyle, S.D.; Allain, V.; Nicol, S.J. Seamounts are hotspots of pelagic biodiversity in the open ocean. Proc. Natl. Acad. Sci. USA 2010, 107, 9707–9711. [Google Scholar] [CrossRef] [PubMed]
- Lopez, J.; Alvarez-Berastegui, D.; Soto, M.; Murua, H. Using fisheries data to model the oceanic habitats of juvenile silky shark (Carcharhinus falciformis) in the tropical eastern Atlantic Ocean. Biodivers. Conserv. 2020, 29, 2377–2397. [Google Scholar] [CrossRef]
- Compagno, L.J.V. FAO Species Catalogue. Vol 4. Sharks of the World: An Annotated and Illustrated Catalogue of Shark Species Known to Date. Part 2. Carcharhiniformes; FAO Fish Synop (125); FAO: Rome, Italy, 1984; pp. 251–655. [Google Scholar]
- Stafford-Deitsch, J. Shark: A Photographer’s Story; Headline Book Publishing: London, UK, 1989. [Google Scholar]
- Syme, J.; Kiszka, J.J.; Parra, G.J. Dynamics of Cetacean Mixed-Species Groups: A Review and Conceptual Framework for Assessing Their Functional Significance. Front. Mar. Sci. 2021, 8, 678173. [Google Scholar] [CrossRef]
- Kiszka, J.J.; Heithaus, M.R.; Wirsing, A.J. Behavioural drivers of the ecological roles and importance of marine mammals. Mar. Ecol. Prog. Ser. 2015, 523, 267–281. [Google Scholar] [CrossRef]
- Talwar, B.S.; Anderson, B.; Avalos-Castillo, C.G.; Blanco-Parra, M.d.P.; Briones, A.; Cardeñosa, D.; Carlson, J.K.; Charvet, P.; Cotton, C.F.; Crysler, Z.; et al. Extinction risk, reconstructed catches and management of chondrichthyan fishes in the Western Central Atlantic Ocean. Fish Fish. 2022, 23, 1150–1179. [Google Scholar] [CrossRef]
- Knowles, J.; Green, A.L.; Dahlgren, C.; Arnett, F.; Knowles, L. Expanding The Bahamas Marine Protected Area Network to Protect 20% of the Marine and Coastal Environment by 2020: A Gap Analysis; The Nature Conservancy: Nassau, Bahamas, 2017; p. 66. [Google Scholar]
- Brooks, E.J.; Sloman, K.A.; Sims, D.W.; Danylchuk, A.J. Validating the use of baited remote underwater video surveys for assessing the diversity, distribution and abundance of sharks in the Bahamas. Endanger. Species Res. 2011, 13, 231–243. [Google Scholar] [CrossRef]
- Kessel, S.T.; Hansell, A.C.; Gruber, S.H.; Guttridge, T.L.; Hussey, N.E.; Perkins, R.G. Three decades of longlining in Bimini, Bahamas, reveals long-term trends in lemon shark Negaprion brevirostris (Carcharhinidae) catch per unit effort. J. Fish Biol. 2016, 88, 2144–2156. [Google Scholar] [CrossRef] [PubMed]
- Hansell, A.C.; Kessel, S.T.; Brewster, L.R.; Cadrin, S.X.; Gruber, S.H.; Skomal, G.B.; Guttridge, T.L. Local indicators of abundance and demographics for the coastal shark assemblage of Bimini, Bahamas. Fish. Res. 2018, 197, 34–44. [Google Scholar] [CrossRef]
- Talwar, B.S.; Stein, J.A.; Connett, S.M.; Liss, S.A.; Brooks, E.J. Results of a fishery-independent longline survey targeting coastal sharks in the eastern Bahamas between 1979 and 2013. Fish. Res. 2020, 230, 105683. [Google Scholar] [CrossRef]
- Smukall, M.J.; Carlson, J.; Kessel, S.T.; Guttridge, T.L.; Dhellemmes, F.; Seitz, A.C.; Gruber, S. Thirty-five years of tiger shark Galeocerdo cuvier relative abundance near Bimini, The Bahamas, and the Southeastern United States with a comparison across jurisdictional bounds. J. Fish Biol. 2022, 101, 13–25. [Google Scholar] [CrossRef]
- Sims, D.W.; Southall, E.J.; Humphries, N.E.; Hays, G.C.; Bradshaw, C.J.A.; Pitchford, J.W.; James, A.; Ahmed, M.Z.; Brierley, A.S.; Hindell, M.A.; et al. Scaling laws of marine predator search behaviour. Nature 2008, 451, 1098–1102. [Google Scholar] [CrossRef]
- Game, E.T.; Grantham, H.S.; Hobday, A.J.; Pressey, R.L.; Lombard, A.T.; Beckley, L.E.; Gjerde, K.; Bustamante, R.; Possingham, H.P.; Richardson, A.J. Pelagic protected areas: The missing dimension in ocean conservation. Trends Ecol. Evol. 2009, 24, 360–369. [Google Scholar] [CrossRef] [PubMed]
- Littler, M.M.; Littler, D.S.; Hanisak, M.D. Deep-water rhodolith distribution, productivity, and growth history at sites of formation and subsequent degradation. J. Exp. Mar. Biol. Ecol. 1991, 150, 163–182. [Google Scholar] [CrossRef]
- Marshall, H.; Skomal, G.; Ross, P.G.; Bernal, D. At-vessel and post-release mortality of the dusky (Carcharhinus obscurus) and sandbar (C. plumbeus) sharks after longline capture. Fish. Res. 2015, 172, 373–384. [Google Scholar] [CrossRef]
- Priede, I.G.; Merrett, N.R. Estimation of abundance of abyssal demersal fishes; a comparison of data from trawls and baited cameras. J. Fish Biol. 1996, 49, 207–216. [Google Scholar] [CrossRef]
- Schneider, E.V.C.; Brooks, E.J.; Cortina, M.P.; Bailey, D.M.; Killen, S.S.; Van Leeuwen, T.E. Design, construction, and deployment of an affordable and long-lasting moored deep-water fish aggregation device. Caribb. Nat. 2021, 83, 1–16. [Google Scholar]
- Francis, M.P. Morphometric minefields—Towards a measurement standard for chondrichthyan fishes. Environ. Biol. Fishes 2006, 77, 407–421. [Google Scholar] [CrossRef]
- Clark, E.; von Schmidt, K. Sharks of the Central Gulf Coast of Florida. Bull. Mar. Sci. 1965, 15, 13–83. [Google Scholar]
- Bonfil, R.; Mena, R.; de Anda, D. Biological parameters of commercially exploited silky shark from the Campeche Bank, Mexico. NOAA Tech. Rep. NMFS 1993, 115, 73–86. [Google Scholar]
- Tambourgi, M.R.d.S.; Hazin, F.H.; Oliveira, P.G.; Coelho, R.; Burgess, G.; Roque, P.C.G. Reproductive aspects of the oceanic whitetip shark, Carcharhinus longimanus (Elasmobranchii: Carcharhinidae), in the equatorial and southwestern Atlantic Ocean. Braz. J. Oceanogr. 2013, 61, 161–168. [Google Scholar] [CrossRef]
- Natanson, L.J.; Casey, J.G.; Kohler, N.E. Age and growth estimates for the dusky shark, Carcharhinus obscurus, in the western North Atlantic Ocean. Fish. Bull. 1995, 93, 116–126. [Google Scholar]
- Branstetter, S.; Musick, J.A.; Colvocoresses, J.A. A comparison of the age and growth of the tiger shark, Galeocerdo cuvier, from off Virginia and from the northwestern Gulf of Mexico. Fish. Bull. 1987, 85, 269–279. [Google Scholar]
- Pratt, H.L. Reproduction in the blue shark, Prionace glauca. Fish. Bull. 1979, 77, 445–470. [Google Scholar]
- Castro, J.I. The shark nursery of Bulls Bay, South Carolina, with a review of the shark nurseries of the southeastern coast of the United States. Environ. Biol. Fishes 1993, 38, 37–48. [Google Scholar] [CrossRef]
- Barker, M.J.; Gruber, S.H.; Newman, S.P.; Schluessel, V. Spatial and ontogenetic variation in growth of nursery-bound juvenile lemon sharks, Negaprion brevirostris: A comparison of two age-assigning techniques. Environ. Biol. Fishes 2005, 72, 343–355. [Google Scholar] [CrossRef]
- Grubbs, R.D.; Musick, J.A.; Conrath, C.L.; Romine, J.G. Long- term movements, migrations, and temporal delineation of a summer nursery for juvenile sandbar sharks in the Chesapeake Bay region. In Shark Nursery Grounds of the Gulf of Mexico and East Coast Waters of the United States; McCandless, C.T., Pratt, H.L., Jr., Kohler, N.E., Eds.; American Fisheries Society: Bethesda, MD, USA, 2007; Volume 50, pp. 87–107. [Google Scholar]
- Rue, H.; Martino, S.; Chopin, N. Approximate Bayesian Inference for Latent Gaussian models by using Integrated Nested Laplace Approximations. J. R. Stat. Soc. Ser. B Stat. Methodol. 2009, 71, 319–392. [Google Scholar] [CrossRef]
- Duncan, K.; Holland, K. Habitat use, growth rates and dispersal patterns of juvenile scalloped hammerhead sharks Sphyrna lewini in a nursery habitat. Mar. Ecol. Prog. Ser. 2006, 312, 211–221. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 9 October 2023).
- Passerotti, M.S.; Andrews, A.H.; Natanson, L.J. Inferring Life History Characteristics of the Oceanic Whitetip Shark Carcharhinus longimanus From Vertebral Bomb Radiocarbon. Front. Mar. Sci. 2020, 7, 1–10. [Google Scholar] [CrossRef]
- Howey, L.A.; Wetherbee, B.M.; Tolentino, E.R.; Shivji, M.S. Biogeophysical and physiological processes drive movement patterns in a marine predator. Mov. Ecol. 2017, 5, 1–16. [Google Scholar] [CrossRef]
- Norman, B.M.; Stevens, J.D. Size and maturity status of the whale shark (Rhincodon typus) at Ningaloo Reef in Western Australia. Fish. Res. 2007, 84, 81–86. [Google Scholar] [CrossRef]
- Berkeley, S.A.; Campos, W.L. Relative abundance and fishery potential of pelagic sharks along Florida’s east coast. Mar. Fish. Rev. 1988, 50, 9–16. [Google Scholar]
- Beerkircher, L.R.; Cortés, E.; Shivji, M. Characteristics of shark bycatch observed on pelagic longlines off the Southeastern United States, 1992–2000. Mar. Fish. Rev. 2002, 64, 40–49. [Google Scholar]
- Pauly, D.; Zeller, D. Catch Reconstruction: Concepts, Methods and Data Sources; Sea Around Us (www.seaaroundus.org); University of British Columbia: Vancouver, BC, Canada, 2015. [Google Scholar]
- Vaudo, J.; Wetherbee, B.; Wood, A.; Weng, K.; Howey-Jordan, L.; Harvey, G.; Shivji, M.S. Vertical movements of shortfin mako sharks Isurus oxyrinchus in the western North Atlantic Ocean are strongly influenced by temperature. Mar. Ecol. Prog. Ser. 2016, 547, 163–175. [Google Scholar] [CrossRef]
- Hayes, C.G.; Jiao, Y.; Cortés, E. Stock Assessment of Scalloped Hammerheads in the Western North Atlantic Ocean and Gulf of Mexico. N. Am. J. Fish. Manag. 2009, 29, 1406–1417. [Google Scholar] [CrossRef]
- Carlson, J.; Charvet, P.; Blanco-Parra, M.P.; Briones Bell-lloch, A.; Cardenosa, D.; Crysler, Z.; Espinoza, E.; Herman, K.; Morales-Saldaña, J.M.; Naranjo-Elizondo, B.; et al. Carcharhinus signatus. The IUCN Red List of Threatened Species. 2021. e.T60219A3094326. Available online: https://www.iucnredlist.org/species/60219/3094326 (accessed on 9 October 2023).
- McCandless, C.T.; Conn, P.; Cooper, P.; Cortés, E.; Laporte, S.W.; Nammack, M. Status Review Report: Northwest Atlantic Dusky Shark (Carcharhinus obscurus); Report to National Marine Fisheries Service; Office of Protected Resources: Silver Spring, MD, USA, 2014.
- SEDAR. Update Assessment to SEDAR 21 HMS Dusky Shark; SEDAR: North Charleston, SC, USA, 2016. [Google Scholar]
- Rigby, C.L.; Barreto, R.; Carlson, J.; Fernando, D.; Fordham, S.; Francis, M.P.; Herman, K.; Jabado, R.W.; Liu, K.M.; Marshall, A.; et al. Carcharhinus obscurus. The IUCN Red List of Threatened Species 2019. e.T3852A2872747. Available online: https://www.iucnredlist.org/species/3852/2872747 (accessed on 16 February 2022).
- Kroetz, A.M.; Gulak, S.J.B.; Carlson, J.K. Horizontal and vertical movements of immature dusky sharks Carcharhinus obscurus in relation to commercial longline fisheries in the western North Atlantic Ocean. Anim. Biotelemetry 2021, 9, 1–12. [Google Scholar] [CrossRef]
- Hacohen-Domené, A.; Polanco-Vásquez, F.; Estupiñan-Montaño, C.; Graham, R.T. Description and characterization of the artisanal elasmobranch fishery on Guatemala’s Caribbean coast. PLoS ONE 2020, 15, e0227797. [Google Scholar] [CrossRef] [PubMed]
- Branstetter, S. Age, growth and reproductive biology of the silky shark, Carcharhinus falciformis, and the scalloped hammerhead, Sphyrna lewini, from the northwestern Gulf of Mexico. Environ. Biol. Fishes 1987, 19, 161–173. [Google Scholar] [CrossRef]
- Gulak, S.J.B.; Enzenauer, M.P.; Carlson, J.K. Characterization of the Shark and Reef Fish Bottom Longline Fisheries: 2012; NOAA Technical Memorandum NMFS-SEFSC-652; NOAA Fisheries: Panama City, FL, USA, 2013; 42p. [Google Scholar]
- Springer, S. Social organization of shark populations. In Sharks, Skates and Rays; Gilbert, P., Mathewson, R., Rall, D., Eds.; John Hopkins Press: Baltimore, MD, USA, 1967; pp. 149–174. [Google Scholar]
- Bonfil, R. Status of shark resources in the southern Gulf of Mexico and Caribbean: Implications for management. Fish. Res. 1997, 29, 101–117. [Google Scholar] [CrossRef]
- Yokota, L.; Lessa, R.P. A nursery Area for Sharks and Rays in Northeastern Brazil. Environ. Biol. Fishes 2006, 75, 349–360. [Google Scholar] [CrossRef]
- Rogers, A.D. The biology of seamounts: 25 years on. Adv. Mar. Biol. 2018, 79, 137–224. [Google Scholar]
- Branstetter, S. Early life-history implications of selected Carcharhinoid and Lamnoid sharks of the northwest Atlantic. NOAA Tech. Rep. NMFS 1990, 90, 17–28. [Google Scholar]
- Heithaus, M. Nursery areas as essential shark habitats: A theoretical perspective. Am. Fish. Soc. Symp. 2007, 50, 3–13. [Google Scholar]
- Bonfil, R. The biology and ecology of the silky shark, Carcharhinus falciformis. In Sharks of the Open Ocean: Biology, Fisheries and Conservation; Camhi, M., Pickitch, E.K., Babcock, E.A., Eds.; Oxford Blackwell Publishing Ltd.: Oxford, UK, 2008. [Google Scholar]
- Gelsleichter, J.; Sparkman, G.; Howey, L.; Brooks, E.; Shipley, O. Elevated accumulation of the toxic metal mercury in the Critically Endangered oceanic whitetip shark Carcharhinus longimanus from the northwestern Atlantic Ocean. Endanger. Species Res. 2020, 43, 267–279. [Google Scholar] [CrossRef]
- Backus, R.H.; Springer, S.; Arnold, E.L., Jr. A contribution to the natural history of the white-tip shark, Pterolamiops longimanus (Poey). Deep. Sea Res. 1956, 3, 178–188. [Google Scholar] [CrossRef]
- Lessa, R.; Santana, F.M.; Paglerani, R. Age, growth and stock structure of the oceanic whitetip shark, Carcharhinus longimanus, from the southwestern equatorial Atlantic. Fish. Res. 1999, 42, 21–30. [Google Scholar] [CrossRef]
- D’Alberto, B.M.; Chin, A.; Smart, J.J.; Baje, L.; White, W.T.; Simpfendorfer, C.A. Age, growth and maturity of oceanic whitetip shark (Carcharhinus longimanus) from Papua New Guinea. Mar. Freshw. Res. 2017, 68, 1118. [Google Scholar] [CrossRef]
- Bass, A.J.; D’Aubrey, J.D.; Kistnasamy, N. Sharks of the east coast of southern Africa. In The Genus Carcharhinus (Carcharhinidae); Investigational Report No. 33; South African Association for Marine Biological Research, The Oceanographic Institute: Durban, South Africa, 1973. [Google Scholar]
- Camargo, S.M.; Coelho, R.; Chapman, D.; Howey-Jordan, L.; Brooks, E.J.; Fernando, D.; Mendes, N.J.; Hazin, F.H.V.; Oliveira, C.; Santos, M.N.; et al. Structure and Genetic Variability of the Oceanic Whitetip Shark, Carcharhinus longimanus, Determined Using Mitochondrial DNA. PLoS ONE 2016, 11, e0155623. [Google Scholar] [CrossRef]
- Castro, J. Sharks of North America; Oxford University Press: New York, NY, USA, 2011. [Google Scholar]
- Bond, M. Department of Biological Sciences, Institute of Environment, Florida International University: North Miami, FL, USA, 2019; unpublished data.
- Hazen, E.; Suryan, R.; Santora, J.; Bograd, S.; Watanuki, Y.; Wilson, R. Scales and mechanisms of marine hotspot formation. Mar. Ecol. Prog. Ser. 2013, 487, 177–183. [Google Scholar] [CrossRef]
- Scales, K.L.; Miller, P.I.; Hawkes, L.A.; Ingram, S.N.; Sims, D.W.; Votier, S.C. REVIEW: On the Front Line: Frontal zones as priority at-sea conservation areas for mobile marine vertebrates. J. Appl. Ecol. 2014, 51, 1575–1583. [Google Scholar] [CrossRef]
- Gove, J.M.; McManus, M.A.; Neuheimer, A.M.L.B.; Polovina, J.J.; Drazen, J.C.; Smith, C.R.; Merrifield, M.A.; Friedlander, A.M.; Ehses, J.S.; Young, C.W.; et al. Near-island biological hotspots in barren ocean basins. Nat. Commun. 2016, 7, 10581. [Google Scholar] [CrossRef]
- Benoit-Bird, K.J.; Au, W.W.L. Prey dynamics affect foraging by a pelagic predator (Stenella longirostris) over a range of spatial and temporal scales. Behav. Ecol. Sociobiol. 2003, 53, 364–373. [Google Scholar] [CrossRef]
- Papastamatiou, Y.P.; Meyer, C.G.; Carvalho, F.; Dale, J.J.; Hutchinson, M.R.; Holland, K.N. Telemetry and random-walk models reveal complex patterns of partial migration in a large marine predator. Ecology 2013, 94, 2595–2606. [Google Scholar] [CrossRef] [PubMed]
- Nadon, M.O.; Baum, J.K.; Williams, I.D.; Mcpherson, J.M.; Zgliczynski, B.J.; Richards, B.L.; Schroeder, R.E.; Brainard, R.E. Re-Creating Missing Population Baselines for Pacific Reef Sharks. Conserv. Biol. 2012, 26, 493–503. [Google Scholar] [CrossRef] [PubMed]
- Williams, I.D.; Baum, J.K.; Heenan, A.; Hanson, K.M.; Nadon, M.O.; Brainard, R.E. Human, Oceanographic and Habitat Drivers of Central and Western Pacific Coral Reef Fish Assemblages. PLoS ONE 2015, 10, e0120516. [Google Scholar] [CrossRef] [PubMed]
- Reid, S.B.; Hirota, J.; Young, R.E.; Hallacher, L.E. Mesopelagic-boundary community in Hawaii: Micronekton at the interface between neritic and oceanic ecosystems. Mar. Biol. 1991, 109, 427–440. [Google Scholar] [CrossRef]
- McManus, M.; Benoit-Bird, K.; Woodson, C.B. Behavior exceeds physical forcing in the diel horizontal migration of the midwater sound-scattering layer in Hawaiian waters. Mar. Ecol. Prog. Ser. 2008, 365, 91–101. [Google Scholar] [CrossRef]
- Santana-Garcon, J.; Braccini, M.; Langlois, T.J.; Newman, S.J.; McAuley, R.B.; Harvey, E.S. Calibration of pelagic stereo-BRUVs and scientific longline surveys for sampling sharks. Methods Ecol. Evol. 2014, 5, 824–833. [Google Scholar] [CrossRef]
- Migura, K.A.; Meadows, D.W. Short-finned pilot whales (Globicephala macrorhynchus) interact with melon-headed whales (Peponocephala electra) in Hawaii. Aquat. Mamm. 2002, 28, 294–297. [Google Scholar]
- Baird, R.W. The Lives of Hawai’i’s Dolphins and Whales: Natural history and Conservation; University of Hawaii Press: Honolulu, HI, USA, 2016. [Google Scholar]
- Papastamatiou, Y.P.; Verbeck, D.; Hutchinson, M.; Bracken-Grissom, H.D.; Chapman, D. An encounter between a pelagic shark and giant cephalopod. J. Fish Biol. 2020, 97, 588–589. [Google Scholar] [CrossRef]
- Clua, E.E.G.; Demarchi, S.; Meyer, C.G. Suspected predatory bites on a snorkeler by an oceanic whitetip shark Carcharhinus longimanus off Moorea island (French Polynesia). J. Forensic Sci. 2021, 66, 2493–2498. [Google Scholar] [CrossRef]
- Fox, S.; Foisy, I.; Venegas, R.D.L.P.; Pastoriza, B.E.G.; Graham, R.T.; Hoffmayer, E.R.; Holmberg, J.; Pierce, S.J. Population structure and residency of whale sharks Rhincodon typus at Utila, Bay Islands, Honduras. J. Fish Biol. 2013, 83, 574–587. [Google Scholar] [CrossRef] [PubMed]
- Heithaus, M.R. Predator–prey and competitive interactions between sharks (order Selachii) and dolphins (suborder Odontoceti): A review. J. Zoo. 2001, 253, 53–68. [Google Scholar] [CrossRef]
- Filmalter, J.D.; Cowley, P.D.; Potier, M.; Ménard, F.; Smale, M.J.; Cherel, Y.; Dagorn, L. Feeding ecology of silky sharks Carcharhinus falciformis associated with floating objects in the western Indian Ocean. J. Fish Biol. 2016, 90, 1321–1337. [Google Scholar] [CrossRef] [PubMed]
- Au, D.W. Polyspecific nature of tuna schools: Shark, dolphin, and seabird associates. Fish. Bull. 1991, 89, 343–354. [Google Scholar]
- Kiszka, J.J.; Woodstock, M.S.; Heithaus, M.R. Functional Roles and Ecological Importance of Small Cetaceans in Aquatic Ecosystems. Front. Mar. Sci. 2022, 9, 1–7. [Google Scholar] [CrossRef]
- Hoffmayer, E.R.; Franks, J.S.; Driggers, W.B.; McKinney, J.A.; Hendon, J.M.; Quattro, J.M. Habitat, movements and environmental preferences of dusky sharks, Carcharhinus obscurus, in the northern Gulf of Mexico. Mar. Biol. 2014, 161, 911–924. [Google Scholar] [CrossRef]
- Howey, L.A.; Tolentino, E.R.; Papastamatiou, Y.P.; Brooks, E.J.; Abercrombie, D.L.; Watanabe, Y.Y.; Williams, S.; Brooks, A.; Chapman, D.D.; Jordan, L.K. Into the deep: The functionality of mesopelagic excursions by an oceanic apex predator. Ecol. Evol. 2016, 6, 5290–5304. [Google Scholar] [CrossRef]
- Quick, N.J.; Isojunno, S.; Sadykova, D.; Bowers, M.; Nowacek, D.P.; Read, A.J. Hidden Markov models reveal complexity in the diving behaviour of short-finned pilot whales. Sci. Rep. 2017, 7, srep45765. [Google Scholar] [CrossRef]
- Haas, A.R.; Fedler, T.; Brooks, E.J. The contemporary economic value of elasmobranchs in The Bahamas: Reaping the rewards of 25 years of stewardship and conservation. Biol. Conserv. 2017, 207, 55–63. [Google Scholar] [CrossRef]
- Anderson, L.; Dahlgren, C.; Knowles, L.; Jupp, L.; Cant-Woodside, S.; Albury-Smith, S.; McKinney-Lambert, C.; Lundy, A. Bahamas Protected Marine Protection Plan for Expanding The Bahamas Marine Protected Areas Network to Meet The Bahamas 2020 Declaration; Bahamas National Trust, Perry Institute for Marine Science, The Nature Conservancy, Bahamas Reef Environmental Educational Foundation: Nassau, Bahamas, 2018. [Google Scholar]
- Sherman, K.D.; Shultz, A.D.; Dahlgren, C.P.; Thomas, C.; Brooks, E.; Brooks, A.; Brumbaugh, D.R.; Gittens, L.; Murchie, K.J. Contemporary and emerging fisheries in The Bahamas—Conservation and management challenges, achievements and future directions. Fish. Manag. Ecol. 2018, 25, 319–331. [Google Scholar] [CrossRef]
- Sadovy, Y.; Domeier, M. Are aggregation-fisheries sustainable? Reef fish fisheries as a case study. Coral Reefs 2005, 24, 254–262. [Google Scholar] [CrossRef]
- Rollin, B.E.; Kessel, M.L. Guidelines for the treatment of animals in behavioural research and teaching. Anim. Behav. 1998, 55, 251–257. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Methods and Effort | Common Name | N (% Catch and Observations) | N Female (N Mature) | N Male (N Mature) | % Female | Male STL (cm) Range | Female STL (cm) Range | Mean STL ± SD (cm) |
|---|---|---|---|---|---|---|---|---|---|
| Northeastern Exuma Sound | Pelagic longline surveys (n = 73 sets, September 2018–March 2020), targeted baiting (35 occasions, September 2018–March 2020), opportunistic encounters (2014–2020) | Silky shark | 20 (59) | 11 (1) | 5 (1) | 69 | 110–240 | 94–240 | 134 ± 39 |
| Dusky shark | 5 (15) | 2 (1) | 3 (3) | 40 | 260–300 | 222–310 | 277 ± 36 | ||
| Oceanic whitetip shark | 3 (9) | 2 (1) | 1 (1) | 67 | 210 | 92–190 | 164 ± 63 | ||
| Lemon shark | 1 (3) | 0 | 1 (1) | 0 | 263 | - | 263 | ||
| Tiger shark | 5 (15) | 2 (1) | 1 (0) | 67 | 179 | 202–327 | 252 ± 63 | ||
| Total | 34 (100) | 61 | 179 ± 74 | ||||||
| Columbus Point, Cat Island | Targeted baiting (62 occasions in April–May from 2011–2018, 15–16 July 2019, 16–18 and 22 June 2020, 29 July 2020), opportunistic encounters (16–18 and 22–23 June 2020) | Silky shark | 15 (7) | 5 (0) | 8 (2) | 38 | 114–260 | 99–199 | 167 ± 45 |
| Dusky shark | 3 (1) | - | 1 (1) | 0 | 238 | - | 238 | ||
| Oceanic whitetip shark | 187 (90) | 167 (162) * | 20 (20) | 89 | 207–285 | 194–307 | 245 ± 23 | ||
| Blue shark | 2 (1) | 2 (1) | - | 100 | - | 208–260 | 234 ± 37 | ||
| Tiger shark | 1 (0.5) | - | 1 (0) | 0 | 232 | - | 232 | ||
| Total | 208 (100) | 85 | 237 ± 34 | ||||||
| San Salvador Seamount | Targeted baiting (6 occasions, 22 January–5 February 2019; 1 occasion, 1 September 2020) | Silky shark | 14 (61) | 7 (2) | 7 (0) | 50 | 136–170 | 110–281 | 167 ± 45 |
| Oceanic whitetip shark | 9 (39) | 3 (3) | 2 (1) | 60 | 158–258 | 189–264 | 226 ± 49 | ||
| Total | 23 (100) | 53 | 182 ± 52 | ||||||
| Mayaguana | Pelagic longline (n = 1 set, 12 July 2019), targeted baiting (2 occasions, 12–13 July 2019) | Silky shark | 1 (17) | - | 1 (1) | 0 | 262 | - | 262 |
| Oceanic whitetip shark | 5 (83) | 3(3) | 2 (1) | 60 | 178–244 | 207–240 | 221 ± 28 | ||
| Total | 6 (100) | 50 | 228 ± 30 |
| Species | y | a | b | x | r2 | N | STL Range | |||
|---|---|---|---|---|---|---|---|---|---|---|
| Oceanic whitetip shark | PCL | = | −6.65 (2.55) | + | 0.93 (0.01) | × | FL | 0.97 | 158 | 92–307 |
| PCL | 14.31 (13.73) | 0.69 (0.06) | TLnat | 0.88 | 24 | 201–285 | ||||
| PCL | −16.78 (3.45) | 0.80 (0.01) | STL | 0.96 | 147 | 92–307 | ||||
| FL | 12.58 (2.56) | 1.04 (0.01) | PCL | 0.97 | 158 | 92–307 | ||||
| FL | 32.10 (13.15) | 0.70 (0.05) | TLnat | 0.89 | 24 | 201–285 | ||||
| FL | −8.65 (3.51) | 0.85 (0.01) | STL | 0.96 | 147 | 92–307 | ||||
| TLnat | 12.57 (18.99) | 1.28 (0.10) | PCL | 0.88 | 24 | 201–285 | ||||
| TLnat | −13.16 (19.76) | 1.27 (0.10) | FL | 0.89 | 24 | 201–285 | ||||
| TLnat | −40.97 (21.93) | 1.13 (0.09) | STL | 0.94 | 13 | 201–285 | ||||
| STL | 30.56 (3.77) | 1.19 (0.02) | PCL | 0.96 | 147 | 92–307 | ||||
| STL | 19.45 (3.80) | 1.13 (0.02) | FL | 0.96 | 147 | 92–307 | ||||
| STL | 49.50 (15.62) | 0.83 (0.06) | TLnat | 0.94 | 13 | 201–285 | ||||
| PCL | = | −2.84 (0.99) | + | 0.93 (0.01) | × | FL | 0.99 | 46 | 94–281 | |
| Silky shark | PCL | −1.12 (1.23) | 0.74 (0.01) | STL | 0.99 | 46 | ||||
| FL | 3.42 (1.05) | 1.08 (0.01) | PCL | 0.99 | 46 | |||||
| FL | 2.08 (1.42) | 0.8 (0.01) | STL | 0.99 | 46 | |||||
| STL | 2.21 (1.64) | 1.35 (0.01) | PCL | 0.99 | 46 | |||||
| STL | −1.79 (1.80) | 1.25 (0.01) | FL | 0.99 | 46 |
| Common Name | Scientific Name | N Shallow | N Deep | % Total Catch | % Female | Mean STL ± SD (cm) | Shallow NES CPUE (n/hook/h) | Deep NES CPUE (n/hook/h) | NES CPUE 2018–2020 (n/1000 hooks) |
|---|---|---|---|---|---|---|---|---|---|
| Silky shark | Carcharhinus falciformis | 1 | 3 | 20 | 25 | 115.9 ± 5.9 | 0.000249 | 0.000239 | 0.77 |
| Tiger shark | Galeocerdo cuvier | 1 | 3 | 20 | 67 | 252 ± 72.5 | 0.000249 | 0.000239 | 0.77 |
| Dusky shark | C. obscurus | 1 | 2 | 15 | 67 | 274.7 ± 46.5 | 0.000249 | 0.000159 | 0.58 |
| Oceanic whitetip shark | C. longimanus | 0 | 1 | 5 | 100 | 92 | 0 | 0.000080 | 0.19 |
| Sharks | 3 | 9 | 60 | 54.5 | 199 ± 89.3 | 0.000746 | 0.000716 | 2.32 | |
| Great barracuda | Sphyraena barracuda | 1 | 2 | 15 | 98.7 ± 7.8 | 0.000249 | 0.000159 | 0.58 | |
| Mahi-mahi | Coryphaena hippurus | 1 | 1 | 10 | 108.5 ± 3.5 | 0.000249 | 0.000080 | 0.39 | |
| Snake mackerel | Gempylus serpens | 0 | 1 | 5 | 70.2 | 0 | 0.000080 | 0.19 | |
| Oilfish | Ruvettus pretiosus | 0 | 1 | 5 | 43.5 | 0 | 0.000080 | 0.19 | |
| Cottonmouth jack | Uraspis secunda | 0 | 1 | 5 | - | 0 | 0.000080 | 0.19 | |
| Teleosts | 2 | 6 | 40 | 89.5 ± 24.5 | 0.000497 | 0.000477 | 1.55 | ||
| Total | 5 | 15 | 100 | 158.6 ± 89.5 | 0.001243 | 0.001193 | 3.87 |
| Site | Shark Species 1 | Shark Species 2 | Other Species 1 | Other Species 2 |
|---|---|---|---|---|
| Northeast-ern Exuma Sound | Oceanic whitetip shark (Carcharhinus longimanus), n = 2: 1 mature male (~210 cm), 1 mature female (~190 cm) | Dusky shark n = 1, mature male, ~300 cm | Short-finned pilot whale (Globicephala macrorhynchus) n > 15 | - |
| Silky shark (C. falciformis) n = 1, immature (~110 cm) | - | Short-finned pilot whale n = 22 | - | |
| Silky shark n = 1, immature female (124 cm) | - | Short-finned pilot whale n = 6 | Pantropical spotted dolphin (Stenella attenuata) n = 8 | |
| Silky shark n = 2: 1 mature female (~240 cm), 1 mature male (~240 cm) | - | Tunas (Thunnini) n > 25 | - | |
| Dusky shark (C. obscurus) n = 1, mature male (~260 cm) | - | Tunas n > 25 | - | |
| Columbus Point, Cat Island | Silky shark n = 2: 1 immature (160 cm), 1 immature (220 cm) | Whale shark (Rhincodon typus) n = 1, mature male (>9 m) | Skipjack tuna (Katsuwonus pelamis) n > 20, Blackfin tuna (Thunnus atlanticus) n > 20, Yellowfin tuna (Thunnus albacares) n > 20 | Anchovies (Engraulidae) n > 1000 |
| Silky shark n = 1, mature male (260 cm) | - | Tunas n > 25 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Talwar, B.S.; Brooks, E.J.; Abercrombie, D.L.; Anderson, B.; Bond, M.E.; Brooks, A.M.L.; Chapman, D.D.; Clementi, G.M.; Fields, C.Y.A.; Gelsleichter, J.; et al. Insights into the Relative Abundance, Life History, and Ecology of Oceanic Sharks in the Eastern Bahamas. Sustainability 2024, 16, 200. https://doi.org/10.3390/su16010200
Talwar BS, Brooks EJ, Abercrombie DL, Anderson B, Bond ME, Brooks AML, Chapman DD, Clementi GM, Fields CYA, Gelsleichter J, et al. Insights into the Relative Abundance, Life History, and Ecology of Oceanic Sharks in the Eastern Bahamas. Sustainability. 2024; 16(1):200. https://doi.org/10.3390/su16010200
Chicago/Turabian StyleTalwar, Brendan S., Edward J. Brooks, Debra L. Abercrombie, Brenda Anderson, Mark E. Bond, Annabelle M. L. Brooks, Demian D. Chapman, Gina M. Clementi, Candace Y. A. Fields, Jim Gelsleichter, and et al. 2024. "Insights into the Relative Abundance, Life History, and Ecology of Oceanic Sharks in the Eastern Bahamas" Sustainability 16, no. 1: 200. https://doi.org/10.3390/su16010200