
Bioactive Compounds from Plant Origin as Natural Antimicrobial Agents for the Treatment of Wound Infections

Abstract
:1. Introduction
2. Results and Discussion
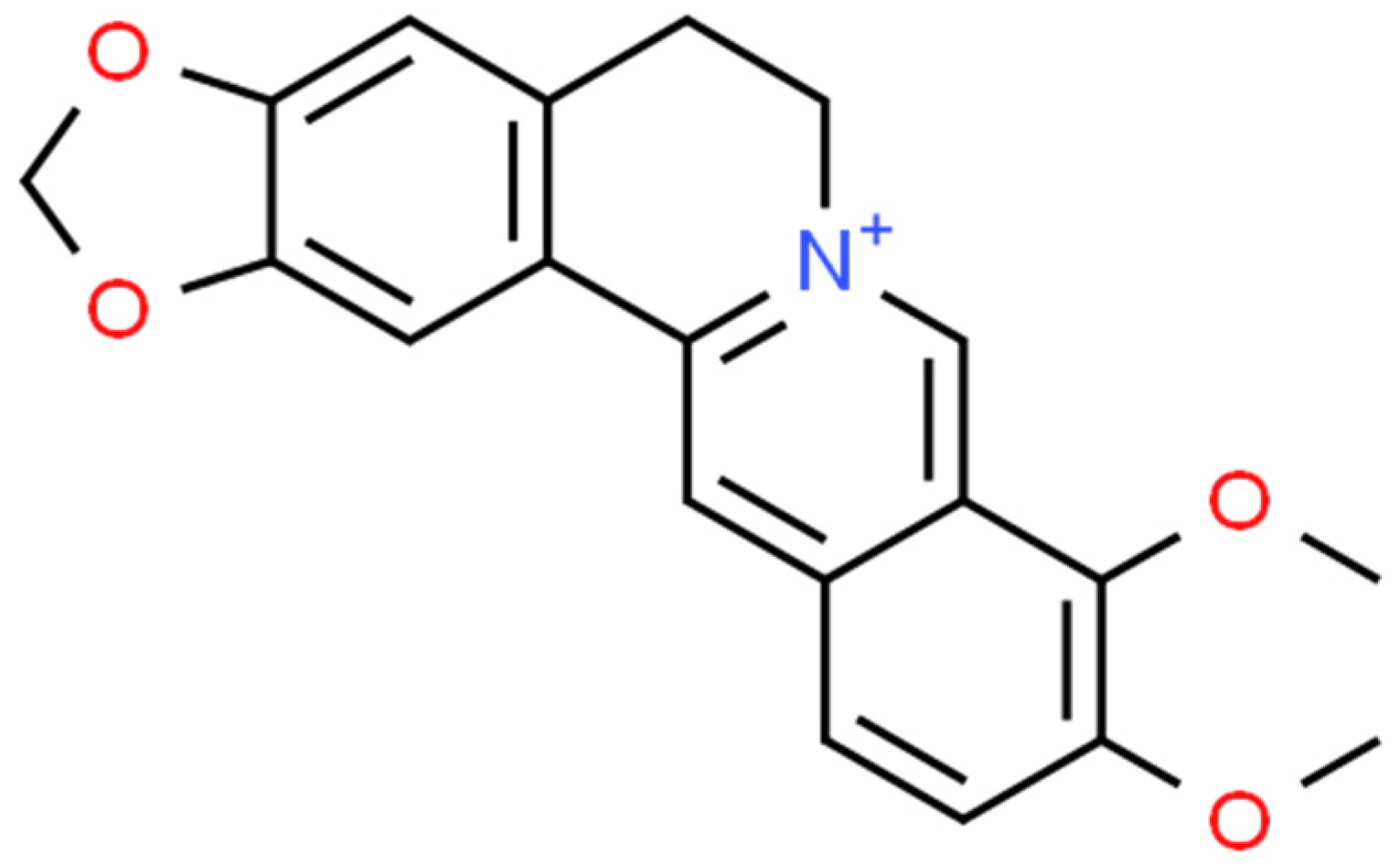
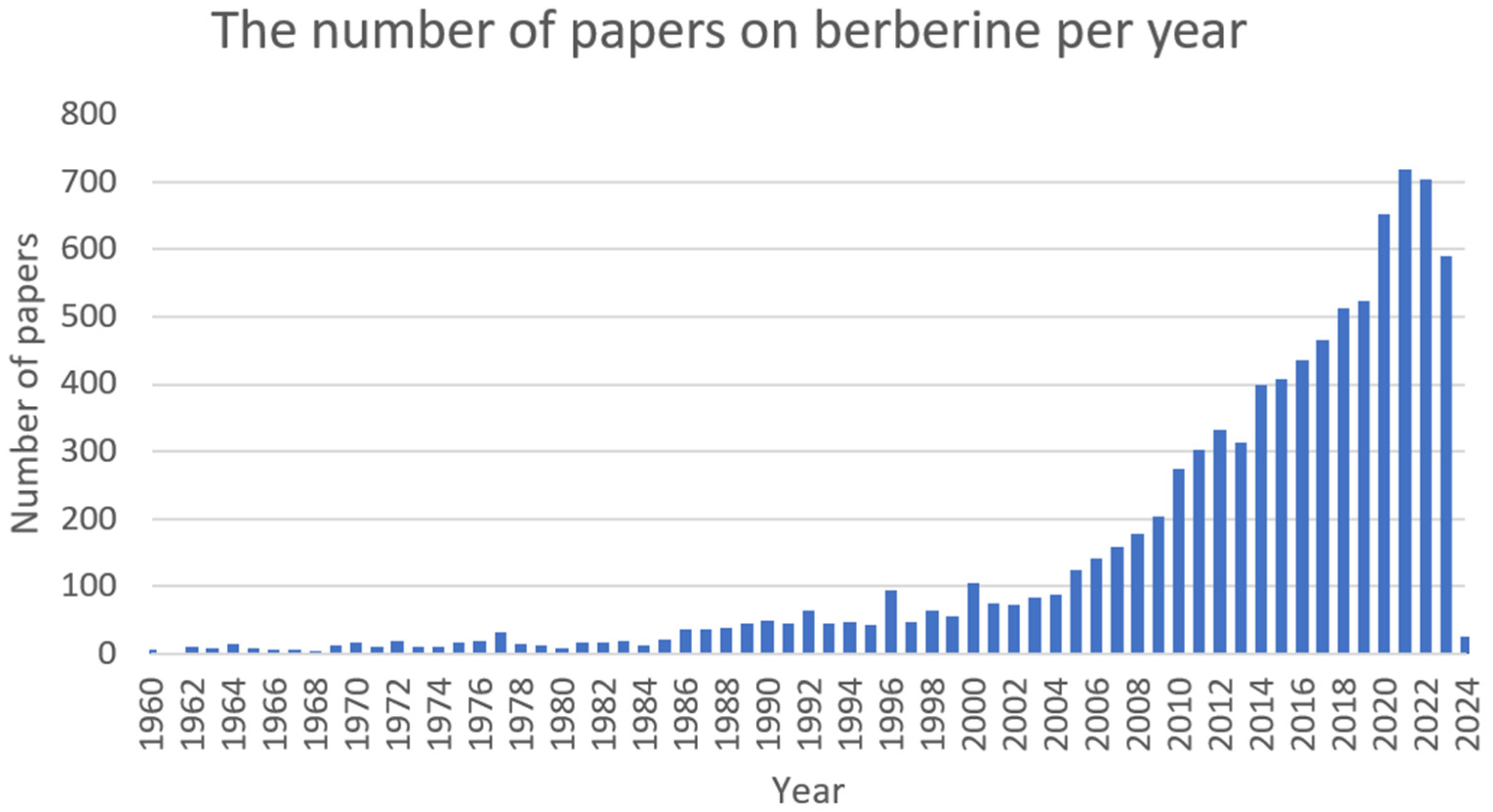
2.1. Berberine (BRB)
2.1.1. Antimicrobial Activity
Staphylococcus aureus
CoNS (Coagulase-Negative Staphylococcus)
Enterococcus spp.
Klebsiella pneumoniae
Acinetobacter baumannii
Escherichia coli
Pseudomonas aeruginosa
2.1.2. Biological Activity
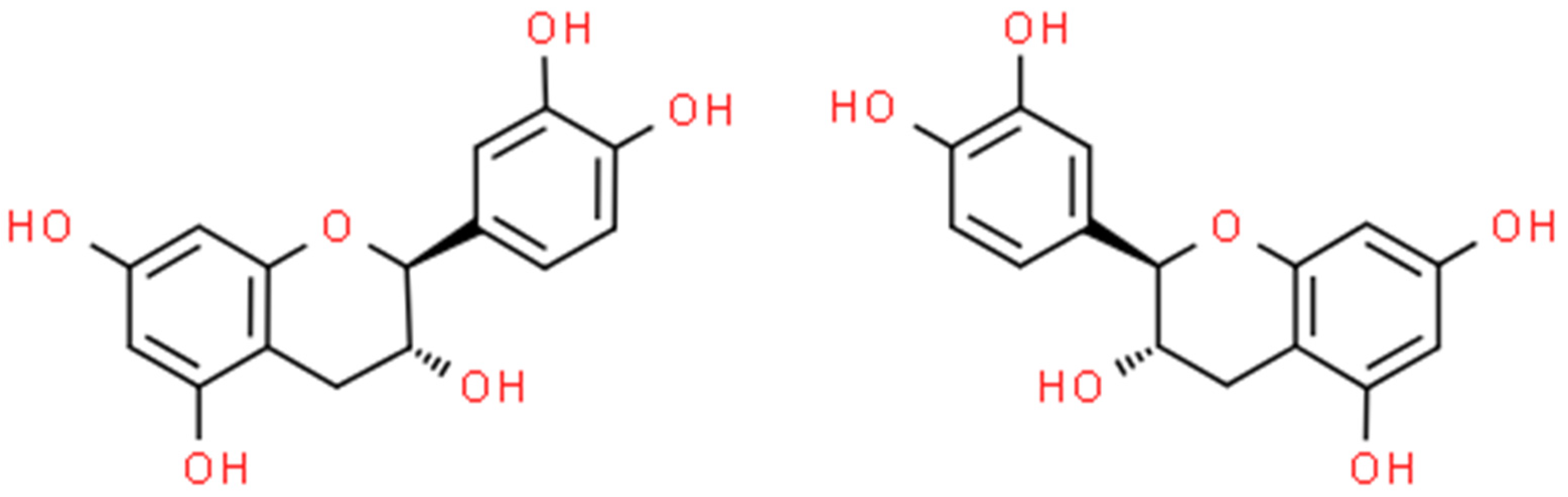
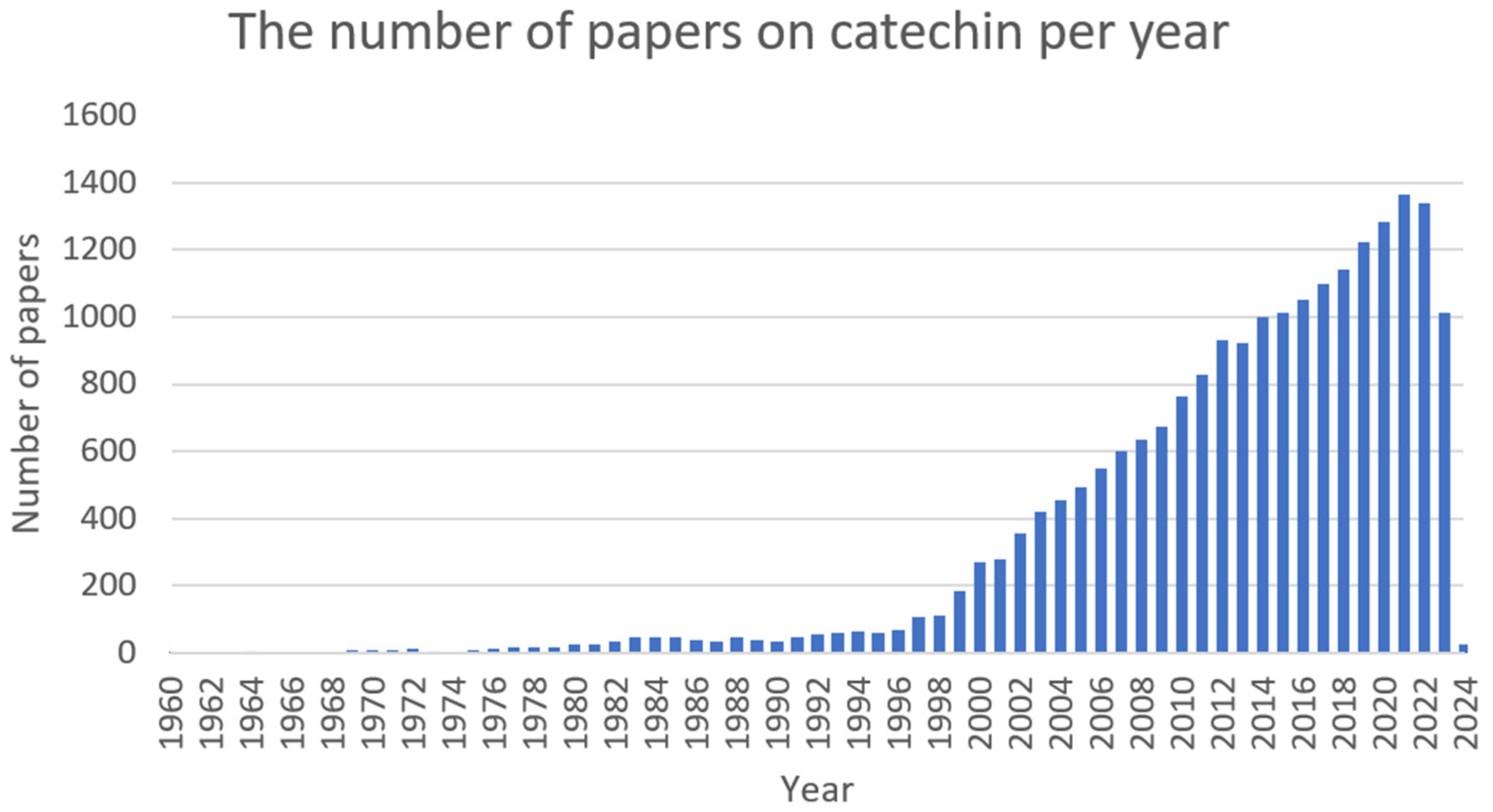
2.2. Catechin (CT)
2.2.1. Antimicrobial Activity
Staphylococccus aureus
Staphylococcus epidermidis
Enterococcus spp.
Klebsiella pneumoniae
Acinetobacter baumannii
Pseudomonas aeruginosa
Escherichia coli
2.2.2. Biological Activity
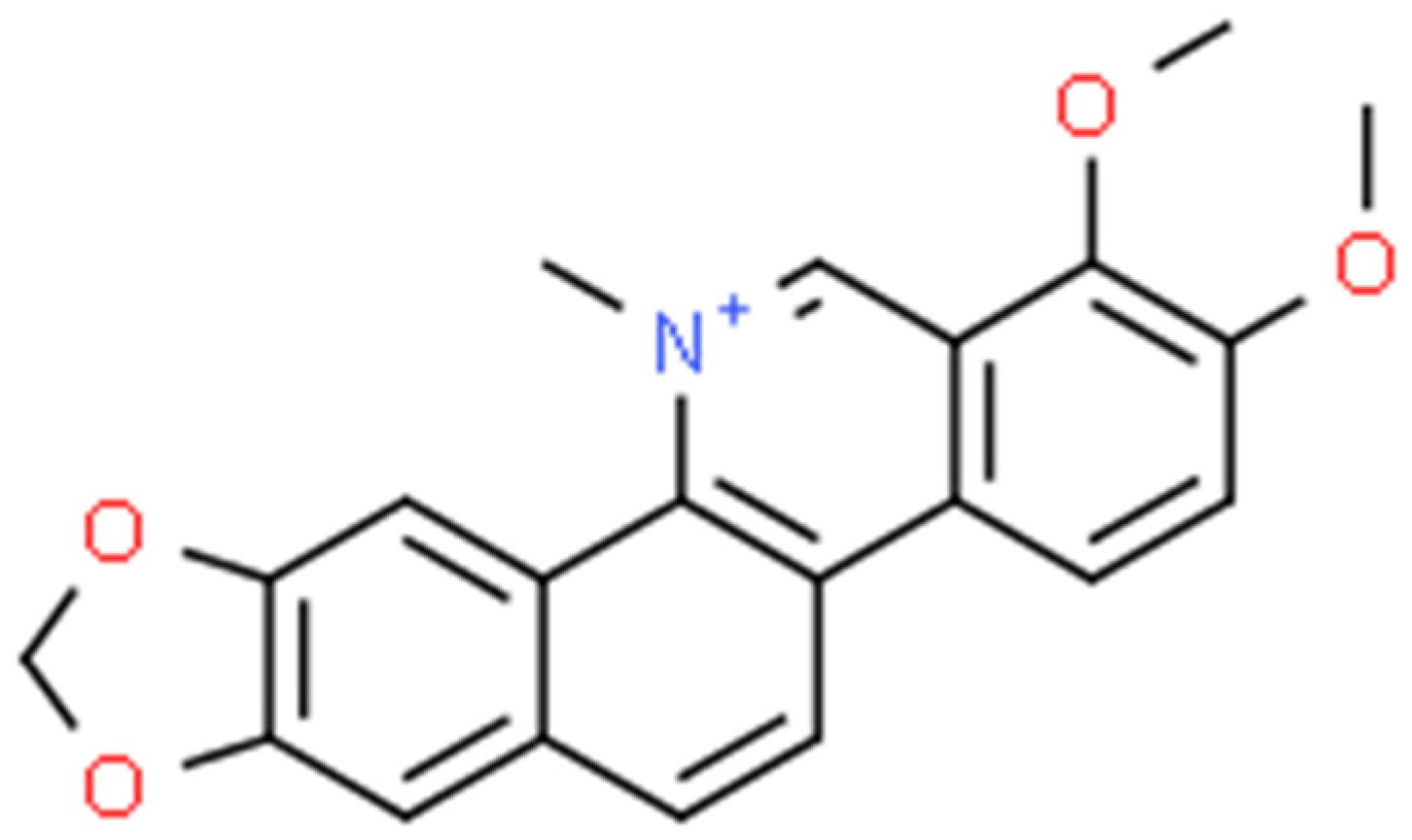
2.3. Chelerythrine (CHE)
2.3.1. Antimicrobial Activity
Staphylococcus spp.
Escherichia coli
Pseudomonas aeruginosa
Serratia marcescens
2.3.2. Biological Activity
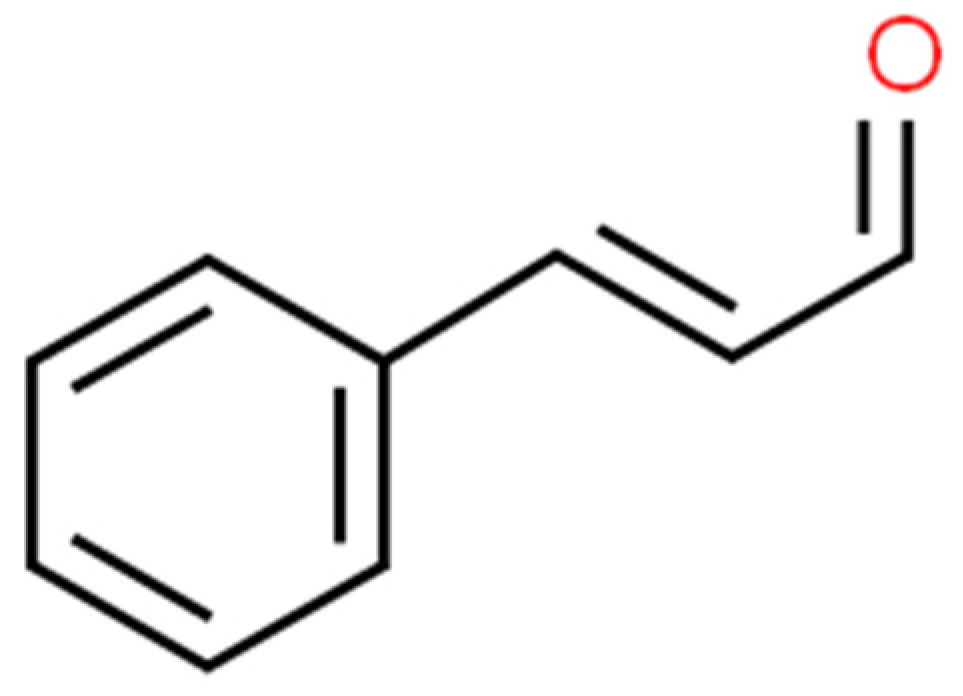
2.4. Cinnamaldehyde (CAN)
2.4.1. Antimicrobial Activity
Staphylococcus aureus
Staphylococcus epidermidis
Enterococcus spp.
Klebsiella pneumoniae
Acinetobacter baumannii
Pseudomonas aeruginosa
Escherichia coli
2.4.2. Biological Activity
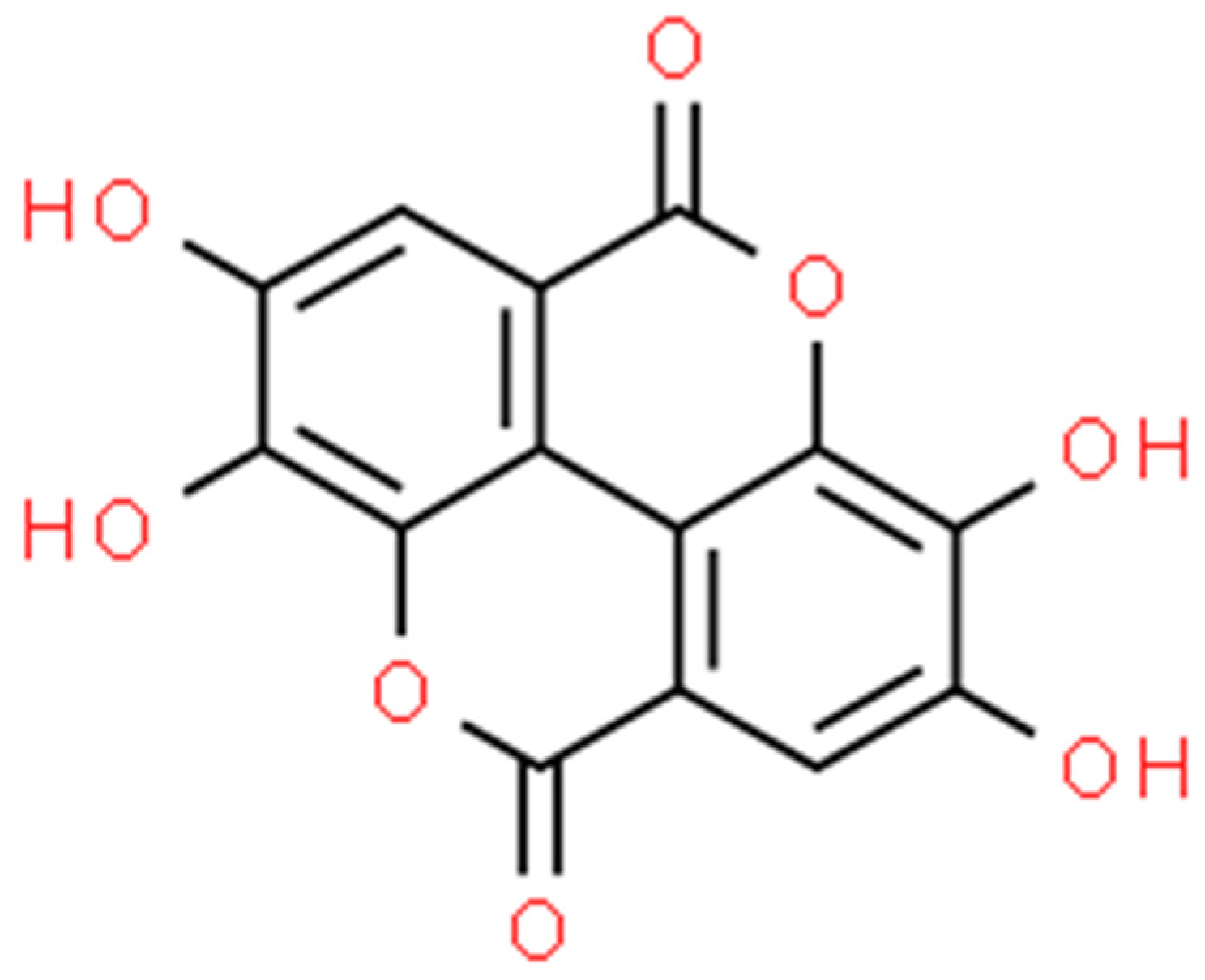
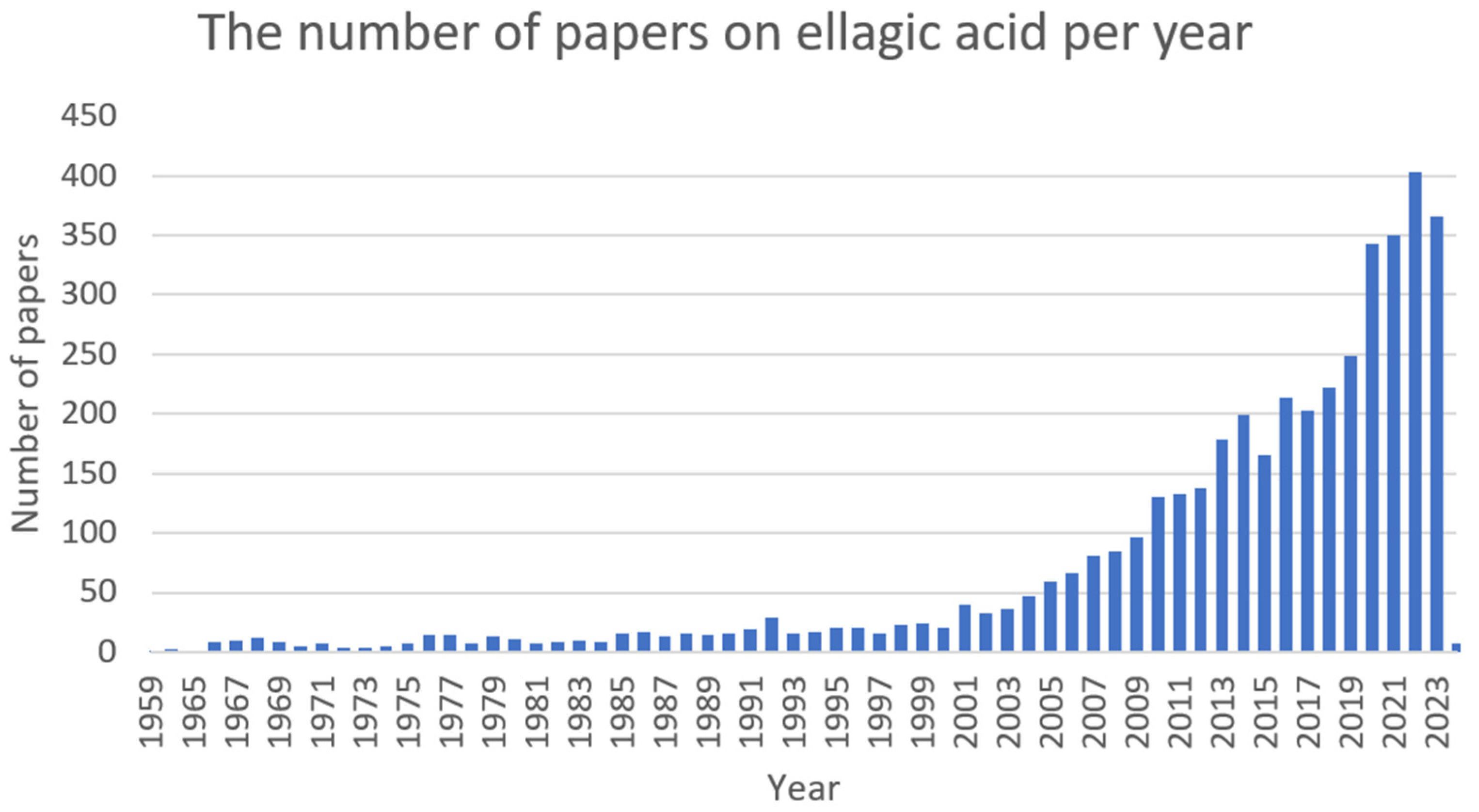
2.5. Ellagic Acid (EA)
2.5.1. Antimicrobial Activity
Staphylococcus aureus
Enterococcus spp.
Klebsiella pneumoniae
Acinetobacter baumannii
Pseudomonas aeruginosa
Escherichia coli
2.5.2. Biological Activity
2.6. Proanthocyanidin (PAC)
2.6.1. Antimicrobial Activity
Staphylococcus spp.
Enterococcus spp.
Escherichia coli
Pseudomonas aeruginosa
2.6.2. Biological Activity
2.7. Sanguinarine (SG)
2.7.1. Antimicrobial Activity
Staphylococcus spp.
Enterococcus spp.
Escherichia coli
Acinetobacter baumannii
2.7.2. Biological Activity
3. Materials and Methods
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Ohshima, H.; Tatemichi, M.; Sawa, T. Chemical Basis of Inflammation-Induced Carcinogenesis. Arch. Biochem. Biophys. 2003, 417, 3–11. [Google Scholar] [CrossRef]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory Responses and Inflammation-Associated Diseases in Organs. Oncotarget 2018, 9, 7204–7218. [Google Scholar] [CrossRef]
- Sans, M.; Panes, J.; Ardite, E.; Elizalde, J.I.; Arce, Y.; Elena, M.; Palacin, A.; Fernandez-Checa, J.C.; Anderson, D.C.; Lobb, R.; et al. VCAM-1 and ICAM-1 Mediate Leukocyte-Endothelial Cell Adhesion in Rat Experimental Colitis. Gastroenterology 1999, 116, 874–883. [Google Scholar] [CrossRef]
- Singh, V.; Kaur, R.; Kumari, P.; Pasricha, C.; Singh, R. ICAM-1 and VCAM-1: Gatekeepers in Various Inflammatory and Cardiovascular Disorders. Clin. Chim. Acta 2023, 548, 117487. [Google Scholar] [CrossRef] [PubMed]
- Abdulkhaleq, L.A.; Assi, M.A.; Abdullah, R.; Zamri-Saad, M.; Taufiq-Yap, Y.H.; Hezmee, M.N.M. The Crucial Roles of Inflammatory Mediators in Inflammation: A Review. Vet. World 2018, 11, 627–635. [Google Scholar] [CrossRef] [PubMed]
- Steinman, L. Blocking Adhesion Molecules as Therapy for Multiple Sclerosis: Natalizumab. Nat. Rev. Drug Discov. 2005, 4, 510–518. [Google Scholar] [CrossRef] [PubMed]
- Ansar, W.; Ghosh, S. Inflammation and Inflammatory Diseases, Markers, and Mediators: Role of CRP in Some Inflammatory Diseases. In Biology of C Reactive Protein in Health and Disease; Springer: Berlin/Heidelberg, Germany, 2016; ISBN 9788132226802. [Google Scholar] [CrossRef]
- Libby, P. Insight Review Articles 868, Inflammation in Atherosclerosis. Nature 2002, 420, 868–874. [Google Scholar] [CrossRef] [PubMed]
- Packard, R.R.S.; Libby, P. Inflammation in Atherosclerosis: From Vascular Biology to Biomarker Discovery and Risk Prediction. Clin. Chem. 2008, 54, 24–38. [Google Scholar] [CrossRef] [PubMed]
- Manohar, M.; Verma, A.K.; Venkateshaiah, S.U.; Sanders, N.L.; Mishra, A. Pathogenic Mechanisms of Pancreatitis. World J. Gastrointest. Pharmacol. Ther. 2017, 8, 10. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Xie, K.; Wolff, R.; Abbruzzese, J.L. Pancreatic Cancer. Lancet 2004, 363, 1049–1057. [Google Scholar] [CrossRef] [PubMed]
- Goggins, M.; Hruban, R.H.; Kern, S.E. BRCA2 Is Inactivated Late in the Development of Pancreatic Intraepithelial Neoplasia: Evidence and Implications. Am. J. Pathol. 2000, 156, 1767–1771. [Google Scholar] [CrossRef]
- Kubes, P.; Mehal, W.Z. Sterile Inflammation in the Liver. Gastroenterology 2012, 143, 1158–1172. [Google Scholar] [CrossRef]
- Walford, H.H.; Doherty, T.A. STAT6 and Lung Inflammation. Jak-Stat 2013, 2, e25301. [Google Scholar] [CrossRef]
- Ernandez, T.; Mayadas, T.N. The Changing Landscape of Renal Inflammation. Trends Mol. Med. 2016, 22, 151–163. [Google Scholar] [CrossRef]
- Strober, W.; Fuss, I.; Mannon, P.; Strober, W.; Fuss, I.; Mannon, P. The Fundamental Basis of Inflammatory Bowel Disease Find the Latest Version : Science in Medicine The Fundamental Basis of Inflammatory Bowel Disease. J. Clin. Investig. 2007, 117, 514–521. [Google Scholar] [CrossRef] [PubMed]
- Fukata, M.; Arditi, M. The Role of Pattern Recognition Receptors in Intestinal Inflammation. Mucosal Immunol. 2013, 6, 451–463. [Google Scholar] [CrossRef] [PubMed]
- Levi, M.; Van Der Poll, T. Two-Way Interactions between Inflammation and Coagulation. Trends Cardiovasc. Med. 2005, 15, 254–259. [Google Scholar] [CrossRef] [PubMed]
- Ekdahl, C.T.; Claasen, J.H.; Bonde, S.; Kokaia, Z.; Lindvall, O. Inflammation Is Detrimental for Neurogenesis in Adult Brain. Proc. Natl. Acad. Sci. USA 2003, 100, 13632–13637. [Google Scholar] [CrossRef] [PubMed]
- Allan, S.M.; Rothwell, N.J. Cytokines and acute neurodegeneration. Neuroscience 2001, 2, 734–744. [Google Scholar] [CrossRef] [PubMed]
- Ramachandran, G. Gram-Positive and Gram-Negative Bacterial Toxins in Sepsis. Landes Biosci. 2014, 5, 213–218. [Google Scholar] [CrossRef] [PubMed]
- Holmes, K.K.; Bertozzi, S.; Bloom, B.R.; Jha, P. Major Infectious Diseases, Disease Control Priorities, 3rd ed.; The International Bank for Reconstruction and Development/The World Bank: Washington, DC, USA, 2017; Volume 6, ISBN 978-1-4648-0524-0. [Google Scholar]
- Steponavičienė, A.; Burokienė, S.; Ivaškevičienė, I.; Stacevičienė, I.; Vaičiūnienė, D.; Jankauskienė, A. Influenza and Respiratory Syncytial Virus Infections in Pediatric Patients during the COVID-19 Pandemic: A Single-Center Experience. Children 2023, 10, 126. [Google Scholar] [CrossRef]
- Mpox. Centers for Disease Control and Preventaion. Available online: https://www.Cdc.Gov/Poxvirus/Mpox/Response/2022/Index.Html (accessed on 6 December 2023).
- Global Measles Threat Continues to Grow as Another Year Passes with Millions of Children Unvaccinated. (404) 639-3286. Available online: https://www.Cdc.Gov/Media/Releases/2023/P1116-Global-Measles.Html (accessed on 6 December 2023).
- De Gier, B.; Marchal, N.; De Beer-Schuurman, I.; Te Wierik, M.; Hooiveld, M.; De Melker, H.E.; Van Sorge, N.M.; Stuart, J.W.T.C.; Melles, D.C.; Van Dijk, K.; et al. Increase in Invasive Group A Streptococcal (Streptococcus Pyogenes) Infections (IGAS) in Young Children in the Netherlands, 2022. Eurosurveillance 2023, 28. [Google Scholar] [CrossRef]
- 2022 Provisional Pertussis Surveillance Report. Available online: https://www.Cdc.Gov/Pertussis/Downloads/Pertuss-Surv-Report-2022_PROVISIONAL.Pdf (accessed on 6 December 2023).
- Beale, M.A.; Marks, M.; Cole, M.J.; Lee, M.K.; Pitt, R.; Ruis, C.; Balla, E.; Crucitti, T.; Ewens, M.; Fernández-Naval, C.; et al. Global Phylogeny of Treponema Pallidum Lineages Reveals Recent Expansion and Spread of Contemporary Syphilis. Nat. Microbiol. 2021, 6, 1549–1560. [Google Scholar] [CrossRef] [PubMed]
- Sexually Transmitted Disease Surveillance 2021. Available online: https://www.Cdc.Gov/Std/Statistics/2021/Default.Htm (accessed on 6 December 2023).
- Hamson, E.; Forbes, C.; Wittkopf, P.; Pandey, A.; Mendes, D.; Kowalik, J.; Czudek, C.; Mugwagwa, T. Impact of Pandemics and Disruptions to Vaccination on Infectious Diseases Epidemiology Past and Present. Hum. Vaccines Immunother. 2023, 19, 2219577. [Google Scholar] [CrossRef]
- Fisman, D.N.; Amoako, A.; Tuite, A.R. Impact of Population Mixing between Vaccinated and Unvaccinated Subpopulations on Infectious Disease Dynamics: Implications for SARS-CoV-2 Transmission. Cmaj 2022, 194, E573–E580. [Google Scholar] [CrossRef]
- Vignier, N.; Bouchaud, O. Meeting the Needs of Mediterranean Nations: Transmissible Diseases in the Mediterranean Area Travel, Migration and Emerging Infectious Diseases. EJIFCC 2018, 29, 175–179. [Google Scholar]
- Castelli, F.; Sulis, G. Migration and Infectious Diseases. Clin. Microbiol. Infect. 2017, 23, 283–289. [Google Scholar] [CrossRef]
- Huang, E.; Yang, X.; Leighton, E.; Li, X. Carbapenem Resistance in the Food Supply Chain. J. Food Prot. 2023, 86, 100108. [Google Scholar] [CrossRef] [PubMed]
- Das, S. The Crisis of Carbapenemase-Mediated Carbapenem Resistance across the Human–Animal–Environmental Interface in India. Infect. Dis. Now 2023, 53, 104628. [Google Scholar] [CrossRef] [PubMed]
- Caliskan-Aydogan, O.; Alocilja, E.C. A Review of Carbapenem Resistance in Enterobacterales and Its Detection Techniques. Microorganisms 2023, 11, 1491. [Google Scholar] [CrossRef]
- El-Mahallawy, H.A.; El Swify, M.; Abdul Hak, A.; Zafer, M.M. Increasing Trends of Colistin Resistance in Patients at High-Risk of Carbapenem-Resistant Enterobacteriaceae. Ann. Med. 2022, 54. [Google Scholar] [CrossRef] [PubMed]
- Kresken, M.; Klare, I.; Wichelhaus, T.A.; Wohlfarth, E.; Layer-Nicolaou, F.; Neumann, B.; Werner, G. Glycopeptide Resistance in Enterococcus spp. and Coagulase-Negative Staphylococci from Hospitalised Patients in Germany: Occurrence, Characteristics and Dalbavancin Susceptibility. J. Glob. Antimicrob. Resist. 2022, 28, 102–107. [Google Scholar] [CrossRef] [PubMed]
- Urban-Chmiel, R.; Marek, A.; Stępień-Pyśniak, D.; Wieczorek, K.; Dec, M.; Nowaczek, A.; Osek, J. Antibiotic Resistance in Bacteria—A Review. Antibiotics 2022, 11, 1079. [Google Scholar] [CrossRef] [PubMed]
- Jian, Z.; Zeng, L.; Xu, T.; Sun, S.; Yan, S.; Yang, L.; Huang, Y.; Jia, J.; Dou, T. Antibiotic Resistance Genes in Bacteria: Occurrence, Spread, and Control. J. Basic Microbiol. 2021, 61, 1049–1070. [Google Scholar] [CrossRef]
- Khameneh, B.; Iranshahy, M.; Soheili, V.; Sedigheh, B.; Bazzaz, F. Khameneh2019.Pdf. Antimicrob. Resist. Infect. Control 2019, 8, 118. [Google Scholar] [CrossRef]
- Drug-Resistant Neisseria Gonorrhoeae. Available online: https://www.Cdc.Gov/Drugresistance/Pdf/Threats-Report/Gonorrhea-508.Pdf (accessed on 6 December 2023).
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, Research, and Development of New Antibiotics: The WHO Priority List of Antibiotic-Resistant Bacteria and Tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef]
- Och, A.; Podgórski, R.; Nowak, R. Biological Activity of Berberine—A Summary Update. Toxins 2020, 12, 713. [Google Scholar] [CrossRef]
- Warowicka, A.; Nawrot, R.; Goździcka-Józefiak, A. Antiviral Activity of Berberine. Arch. Virol. 2020, 165, 1935–1945. [Google Scholar] [CrossRef]
- Neag, M.A.; Mocan, A.; Echeverría, J.; Pop, R.M.; Bocsan, C.I.; Crisan, G.; Buzoianu, A.D. Berberine: Botanical Occurrence, Traditional Uses, Extraction Methods, and Relevance in Cardiovascular, Metabolic, Hepatic, and Renal Disorders. Front. Pharmacol. 2018, 9, 557. [Google Scholar] [CrossRef]
- Xu, J.; Long, Y.; Ni, L.; Yuan, X.; Yu, N.; Wu, R.; Tao, J.; Zhang, Y. Anticancer Effect of Berberine Based on Experimental Animal Models of Various Cancers: A Systematic Review and Meta-Analysis. BMC Cancer 2019, 19, 589. [Google Scholar] [CrossRef] [PubMed]
- Tan, W.; Li, Y.; Chen, M.; Wang, Y. Berberine Hydrochloride: Anticancer Activity and Nanoparticulate Delivery System. Int. J. Nanomed. 2011, 6, 1773–1777. [Google Scholar] [CrossRef]
- Kong, W.; Wei, J.; Abidi, P.; Lin, M.; Inaba, S.; Li, C.; Wang, Y.; Wang, Z.; Si, S.; Pan, H.; et al. Berberine Is a Novel Cholesterol-Lowering Drug Working through a Unique Mechanism Distinct from Statins. Nat. Med. 2004, 10, 1344–1351. [Google Scholar] [CrossRef]
- Cicero, A.F.G.; Tartagni, E. Antidiabetic Properties of Berberine: From Cellular Pharmacology to Clinical Effects. Hosp. Pract. 2012, 40, 56–63. [Google Scholar] [CrossRef]
- Ilyas, Z.; Perna, S.; Al-thawadi, S.; Alalwan, T.A.; Riva, A.; Petrangolini, G.; Gasparri, C.; Infantino, V.; Peroni, G.; Rondanelli, M. The Effect of Berberine on Weight Loss in Order to Prevent Obesity: A Systematic Review. Biomed. Pharmacother. 2020, 127, 110137. [Google Scholar] [CrossRef]
- Yi, M.; Jun-ping, C.; Li, X.; Xia, L.; Dong-mei, L.; Hui-ping, S. Hypolipidemic Effect of Combination of Berberine and Metformin on Experimental Hyperlipidemia Models. Lab. Anim. Comp. Med. 2014, 2006, 228–231. [Google Scholar] [CrossRef]
- Zhou, M.; Deng, Y.; Liu, M.; Liao, L.; Dai, X.; Guo, C.; Zhao, X.; He, L.; Peng, C.; Li, Y. The Pharmacological Activity of Berberine, a Review for Liver Protection. Eur. J. Pharmacol. 2021, 890, 173655. [Google Scholar] [CrossRef] [PubMed]
- Suadoni, M.T.; Atherton, I. Berberine for the Treatment of Hypertension: A Systematic Review. Complement. Ther. Clin. Pract. 2021, 42, 101287. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, S.K.; Dhir, A. On the Mechanism of Antidepressant-like Action of Berberine Chloride. Eur. J. Pharmacol. 2008, 589, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Kuo, C.L.; Chi, C.W.; Liu, T.Y. The Anti-Inflammatory Potential of Berberine In Vitro and In Vivo. Cancer Lett. 2004, 203, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Jin, X.; Liang, C.; Bu, F.; Pan, D.; He, Q.; Ming, Y.; Little, P.; Du, H.; Liang, S.; et al. Berberine for Diarrhea in Children and Adults: A Systematic Review and Meta-Analysis. Ther. Adv. Gastroenterol. 2020, 13, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Liang, R.; Zhang, T.; Yuan, Y.; Shen, S.; Ye, H. Microarray Analysis of the Transcriptome of the Escherichia coli (E. coli) Regulated by Cinnamaldehyde (CMA). Food Agric. Immunol. 2017, 28, 500–515. [Google Scholar] [CrossRef]
- Shou, J.W.; Shaw, P.C. Therapeutic Efficacies of Berberine against Neurological Disorders: An Update of Pharmacological Effects and Mechanisms. Cells 2022, 11, 796. [Google Scholar] [CrossRef]
- Jian-Ling, J.; Guo-Qiang, H.; Zhen, M.; Gao, P.-J. Antibacterial Mechanisms of Berberine and Reasons for Little Resistance of Bacteria. Chin. Herb. Med. 2010, 3, 27–35. [Google Scholar] [CrossRef]
- Hayashi, K.; Minoda, K.; Nagaoka, Y.; Hayashi, T.; Uesato, S. Antiviral Activity of Berberine and Related Compounds against Human Cytomegalovirus. Bioorg. Med. Chem. Lett. 2007, 17, 1562–1564. [Google Scholar] [CrossRef]
- Zhang, S.-L.; Chang, J.-J.; Damu, G.L.V.; Fang, B.; Zhou, X.-D.; Geng, R.-X.; Zhou, C.-H. Novel Berberine Triazoles: Synthesis, Antimicrobial Evaluation and Competitive Interactions with Metal Ions to Human Serum Albumin. Bioorg. Med. Chem. Lett. 2013, 23, 1008–1012. [Google Scholar] [CrossRef]
- Sahibzada, M.U.K.; Sadiq, A.; Faidah, H.S.; Khurram, M.; Amin, M.U.; Haseeb, A.; Kakar, M. Berberine Nanoparticles with Enhanced In Vitro Bioavailability: Characterization and Antimicrobial Activity. Drug Des. Dev. Ther. 2018, 12, 303–312. [Google Scholar] [CrossRef]
- Zhang, C.; Li, Z.; Pan, Q.; Fan, L.; Pan, T.; Zhu, F.; Pan, Q.; Shan, L.; Zhao, L. Berberine at Sub-Inhibitory Concentration Inhibits Biofilm Dispersal in Staphylococcus aureus. Microbiology 2022, 168. [Google Scholar] [CrossRef]
- Zheng, T.; Cui, M.; Chen, H.; Wang, J.; Ye, H.; Zhang, Q.; Sun, S.; Feng, Y.; Zhang, Y.; Liu, W.; et al. Co-Assembled Nanocomplexes Comprising Epigallocatechin Gallate and Berberine for Enhanced Antibacterial Activity against Multidrug Resistant Staphylococcus aureus. Biomed. Pharmacother. 2023, 163, 114856. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; Wang, J.; Yang, C.; Zhu, C.; Guo, G.; Tang, J.; Shen, H. Antimicrobial Characteristics of Berberine against Prosthetic Joint Infection-Related Staphylococcus aureus of Different Multi-Locus Sequence Types. BMC Complement. Altern. Med. 2019, 19, 218. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, P.; Hu, X.; Zhang, Y.; Lu, X.; Li, C.; Nie, T.; Li, G.; Wang, X.; Pang, J.; et al. The Combined Antibacterial Effects of Sodium New Houttuyfonate and Berberine Chloride against Growing and Persistent Methicillin-Resistant and Vancomycin-Intermediate Staphylococcus aureus. BMC Microbiol. 2020, 20, 317. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Liu, S.; Zhao, C.; Zhang, X.; Ning, Y.; Lu, F. Inhibition of Berberine on Methicillin-Resistant Staphylococcus aureus Biofilm Formation. Rev. Bras. Farmacogn. 2022, 32, 455–459. [Google Scholar] [CrossRef]
- Sun, H.; Ansari, M.F.; Fang, B.; Zhou, C.-H. Natural Berberine-Hybridized Benzimidazoles as Novel Unique Bactericides against Staphylococcus aureus. J. Agric. Food Chem. 2021, 69, 7831–7840. [Google Scholar] [CrossRef] [PubMed]
- Xia, S.; Ma, L.; Wang, G.; Yang, J.; Zhang, M.; Wang, X.; Su, J.; Xie, M. In Vitro Antimicrobial Activity and the Mechanism of Berberine against Methicillin-Resistant Staphylococcus aureus Isolated from Bloodstream Infection Patients. Infect. Drug Resist. 2022, 15, 1933–1944. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Sun, X.; Wu, J.; Wu, Y.; Wang, Y.; Hu, X.; Wang, X. Berberine Damages the Cell Surface of Methicillin-Resistant Staphylococcus aureus. Front. Microbiol. 2020, 11, 621. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Wang, P.; Li, T.; Tian, X.; Guo, W.; Xu, B.; Huang, G.; Cai, D.; Zhou, F.; Zhang, H.; et al. Self-Assemblies Based on Traditional Medicine Berberine and Cinnamic Acid for Adhesion-Induced Inhibition Multidrug-Resistant Staphylococcus aureus. ACS Appl. Mater. Interfaces 2020, 12, 227–237. [Google Scholar] [CrossRef] [PubMed]
- Liang, R.; Yong, X.; Duan, Y.; Tan, Y.; Zeng, P.; Zhou, Z.; Jiang, Y.; Wang, S.; Jiang, Y.; Huang, X.; et al. Potent In Vitro Synergism of Fusidic Acid (FA) and Berberine Chloride (BBR) against Clinical Isolates of Methicillin-Resistant Staphylococcus aureus (MRSA). World J. Microbiol. Biotechnol. 2014, 30, 2861–2869. [Google Scholar] [CrossRef] [PubMed]
- Chu, M.; Zhang, M.-B.; Liu, Y.-C.; Kang, J.-R.; Chu, Z.-Y.; Yin, K.-L.; Ding, L.-Y.; Ding, R.; Xiao, R.-X.; Yin, Y.-N.; et al. Role of Berberine in the Treatment of Methicillin-Resistant Staphylococcus aureus Infections. Sci. Rep. 2016, 6, 24748. [Google Scholar] [CrossRef]
- Fan, T.-Y.; Wang, Y.-X.; Tang, S.; Hu, X.-X.; Zen, Q.-X.; Pang, J.; Yang, Y.-S.; You, X.-F.; Song, D.-Q. Synthesis and Antibacterial Evaluation of 13-Substituted Cycloberberine Derivatives as a Novel Class of Anti-MRSA Agents. Eur. J. Med. Chem. 2018, 157, 877–886. [Google Scholar] [CrossRef]
- Bhatia, E.; Sharma, S.; Jadhav, K.; Banerjee, R. Combinatorial Liposomes of Berberine and Curcumin Inhibit Biofilm Formation and Intracellular Methicillin Resistant Staphylococcus aureus Infections and Associated Inflammation. J. Mater. Chem. B 2021, 9, 864–875. [Google Scholar] [CrossRef]
- Aksoy, C.S.; Avci, F.G.; Ugurel, O.M.; Atas, B.; Sayar, N.A.; Sariyar Akbulut, B. Potentiating the Activity of Berberine for Staphylococcus aureus in a Combinatorial Treatment with Thymol. Microb. Pathog. 2020, 149, 104542. [Google Scholar] [CrossRef]
- Wojtyczka, R.D.; Dziedzic, A.; Kępa, M.; Kubina, R.; Kabała-Dzik, A.; Mularz, T.; Idzik, D. Berberine Enhances the Antibacterial Activity of Selected Antibiotics against Coagulase-Negative Staphylococcus Strains In Vitro. Molecules 2014, 19, 6583–6596. [Google Scholar] [CrossRef]
- Zhou, X.; Yang, C.; Li, Y.; Liu, X.; Wang, Y. Potential of Berberine to Enhance Antimicrobial Activity of Commonly Used Antibiotics for Dairy Cow Mastitis Caused by Multiple Drug-Resistant Staphylococcus Epidermidis Infection. Genet. Mol. Res. 2015, 14, 9683–9692. [Google Scholar] [CrossRef]
- Chen, L.; Bu, Q.; Xu, H.; Liu, Y.; She, P.; Tan, R.; Wu, Y. The Effect of Berberine Hydrochloride on Enterococcus Faecalis Biofilm Formation and Dispersion In Vitro. Microbiol. Res. 2016, 186–187, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Guefack, M.-G.F.; Messina, N.D.M.; Mbaveng, A.T.; Nayim, P.; Kuete, J.R.N.; Matieta, V.Y.; Chi, G.F.; Ngadjui, B.T.; Kuete, V. Antibacterial and Antibiotic-Potentiation Activities of the Hydro-Ethanolic Extract and Protoberberine Alkaloids from the Stem Bark of Enantia Chlorantha against Multidrug-Resistant Bacteria Expressing Active Efflux Pumps. J. Ethnopharmacol. 2022, 296, 115518. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.-Y.; Ye, X.-G.; He, L.-T.; Zhang, S.-R.; Wang, R.-L.; Zhou, J.; He, Z.-S. In Vitro Characterization and Inhibition of the Interaction between Ciprofloxacin and Berberine against Multidrug-Resistant Klebsiella Pneumoniae. J. Antibiot. 2016, 69, 741–746. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Song, Y.; Wang, L.; Kang, G.; Wang, P.; Yin, H.; Huang, H. A Potential Combination Therapy of Berberine Hydrochloride with Antibiotics against Multidrug-Resistant Acinetobacter Baumannii. Front. Cell. Infect. Microbiol. 2021, 11, 660431. [Google Scholar] [CrossRef]
- Gao, W.-W.; Gopala, L.; Bheemanaboina, R.R.Y.; Zhang, G.-B.; Li, S.; Zhou, C.-H. Discovery of 2-Aminothiazolyl Berberine Derivatives as Effectively Antibacterial Agents toward Clinically Drug-Resistant Gram-Negative Acinetobacter Baumanii. Eur. J. Med. Chem. 2018, 146, 15–37. [Google Scholar] [CrossRef]
- Ahmadi, F.; Khalvati, B.; Eslami, S.; Mirzaii, M.; Roustaei, N.; Mazloomirad, F.; Khoramrooz, S.S. The Inhibitory Effect of Thioridazine on AdeB Efflux Pump Gene Expression in Multidrug-Resistant Acinetobacter Baumannii Isolates Using Real Time PCR. Avicenna J. Med. Biotechnol. 2022, 14, 132–136. [Google Scholar] [CrossRef] [PubMed]
- Thakur, P.; Chawla, R.; Goel, R.; Narula, A.; Arora, R.; Sharma, R.K. Augmenting the Potency of Third-Line Antibiotics with Berberis Aristata : In Vitro Synergistic Activity against Carbapenem-Resistant Escherichia coli. J. Glob. Antimicrob. Resist. 2016, 6, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Bandyopadhyay, S.; Patra, P.H.; Mahanti, A.; Mondal, D.K.; Dandapat, P.; Bandyopadhyay, S.; Samanta, I.; Lodh, C.; Bera, A.K.; Bhattacharyya, D.; et al. Potential Antibacterial Activity of Berberine against Multi Drug Resistant Enterovirulent Escherichia coli Isolated from Yaks (Poephagus Grunniens) with Haemorrhagic Diarrhoea. Asian Pac. J. Trop. Med. 2013, 6, 315–319. [Google Scholar] [CrossRef]
- Li, Y.; Ge, X. Role of Berberine as a Potential Efflux Pump Inhibitor against MdfA from Escherichia coli: In Vitro and In Silico Studies. Microbiol. Spectr. 2023, 11. [Google Scholar] [CrossRef]
- Zhang, G.-B.; Maddili, S.K.; Tangadanchu, V.K.R.; Gopala, L.; Gao, W.-W.; Cai, G.-X.; Zhou, C.-H. Discovery of Natural Berberine-Derived Nitroimidazoles as Potentially Multi-Targeting Agents against Drug-Resistant Escherichia coli. Sci. China Chem. 2018, 61, 557–568. [Google Scholar] [CrossRef]
- Sun, H.; Huang, S.-Y.; Jeyakkumar, P.; Cai, G.-X.; Fang, B.; Zhou, C.-H. Natural Berberine-Derived Azolyl Ethanols as New Structural Antibacterial Agents against Drug-Resistant Escherichia coli. J. Med. Chem. 2022, 65, 436–459. [Google Scholar] [CrossRef] [PubMed]
- Morita, Y.; Nakashima, K.-I.; Nishino, K.; Kotani, K.; Tomida, J.; Inoue, M.; Kawamura, Y. Berberine Is a Novel Type Efflux Inhibitor Which Attenuates the MexXY-Mediated Aminoglycoside Resistance in Pseudomonas Aeruginosa. Front. Microbiol. 2016, 7, 1223. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Huang, J.; Li, L.; Liu, L. Synergistic Activity of Berberine with Azithromycin against Pseudomonas Aeruginosa Isolated from Patients with Cystic Fibrosis of Lung In Vitro and In Vivo. Cell. Physiol. Biochem. 2017, 42, 1657–1669. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Guo, M.; Xu, X.; Hu, Y.; Liu, D.; Wang, C.; Liu, X.; Li, Y. In Vitro Synergistic Inhibitory Activity of Natural Alkaloid Berberine Combined with Azithromycin against Alginate Production by Pseudomonas Aeruginosa PAO1. Oxid. Med. Cell. Longev. 2022, 2022, 3858500. [Google Scholar] [CrossRef]
- Laudadio, E.; Cedraro, N.; Mangiaterra, G.; Citterio, B.; Mobbili, G.; Minnelli, C.; Bizzaro, D.; Biavasco, F.; Galeazzi, R. Natural Alkaloid Berberine Activity against Pseudomonas Aeruginosa MexXY-Mediated Aminoglycoside Resistance: In Silico and In Vitro Studies. J. Nat. Prod. 2019, 82, 1935–1944. [Google Scholar] [CrossRef] [PubMed]
- Giorgini, G.; Mangiaterra, G.; Cedraro, N.; Laudadio, E.; Sabbatini, G.; Cantarini, M.; Minnelli, C.; Mobbili, G.; Frangipani, E.; Biavasco, F.; et al. Berberine Derivatives as Pseudomonas Aeruginosa MexXY-OprM Inhibitors: Activity and In Silico Insights. Molecules 2021, 26, 6644. [Google Scholar] [CrossRef]
- Mangiaterra, G.; Cedraro, N.; Laudadio, E.; Minnelli, C.; Citterio, B.; Andreoni, F.; Mobbili, G.; Galeazzi, R.; Biavasco, F. The Natural Alkaloid Berberine Can Reduce the Number of Pseudomonas Aeruginosa Tolerant Cells. J. Nat. Prod. 2021, 84, 993–1001. [Google Scholar] [CrossRef]
- Kavanaugh, L.G.; Mahoney, A.R.; Dey, D.; Wuest, W.M.; Conn, G.L. Di-Berberine Conjugates as Chemical Probes of Pseudomonas Aeruginosa MexXY-OprM Efflux Function and Inhibition. bioRxiv 2023. [Google Scholar] [CrossRef]
- Aghayan, S.S.; Kalalian Mogadam, H.; Fazli, M.; Darban-Sarokhalil, D.; Khoramrooz, S.S.; Jabalameli, F.; Yaslianifard, S.; Mirzaii, M. The Effects of Berberine and Palmatine on Efflux Pumps Inhibition with Different Gene Patterns in Pseudomonas Aeruginosa Isolated from Burn Infections. Avicenna J. Med. Biotechnol. 2017, 9, 2–7. [Google Scholar]
- Wu, S.; Yang, K.; Hong, Y.; Gong, Y.; Ni, J.; Yang, N.; Ding, W. A New Perspective on the Antimicrobial Mechanism of Berberine Hydrochloride against Staphylococcus aureus Revealed by Untargeted Metabolomic Studies. Front. Microbiol. 2022, 13, 917414. [Google Scholar] [CrossRef]
- Guo, N.; Zhao, X.; Li, W.; Shi, C.; Meng, R.; Liu, Z.; Yu, L. The Synergy of Berberine Chloride and Totarol against Staphylococcus aureus Grown in Planktonic and Biofilm Cultures. J. Med. Microbiol. 2015, 64, 891–900. [Google Scholar] [CrossRef]
- Karaosmanoglu, K.; Sayar, N.A.; Kurnaz, I.A.; Akbulut, B.S. Assessment of Berberine as a Multi-Target Antimicrobial: A Multi-Omics Study for Drug Discovery and Repositioning. OMICS 2014, 18, 42–53. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Li, X.-D.; Hong, J.; Liu, C.; Zhang, X.-L.; Zheng, J.-P.; Xu, Y.-J.; Ou, Z.-Y.; Zheng, J.-L.; Yu, D.-J. Inhibitory Effect of Two Traditional Chinese Medicine Monomers, Berberine and Matrine, on the Quorum Sensing System of Antimicrobial-Resistant Escherichia coli. Front. Microbiol. 2019, 10, 2584. [Google Scholar] [CrossRef] [PubMed]
- Su, F.; Wang, J. Berberine Inhibits the MexXY-OprM Efflux Pump to Reverse Imipenem Resistance in a Clinical Carbapenem-resistant Pseudomonas Aeruginosa Isolate in a Planktonic State. Exp. Ther. Med. 2017, 15, 467–472. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Ge, C.; Ye, J.; Li, R.; Zhang, Y. Demethyleneberberine Alleviates Pseudomonas Aeruginosa-Induced Acute Pneumonia by Inhibiting the AIM2 Inflammasome and Oxidative Stress. Pulm. Pharmacol. Ther. 2023, 83, 102259. [Google Scholar] [CrossRef] [PubMed]
- Aswathanarayan, J.B.; Vittal, R.R. Inhibition of Biofilm Formation and Quorum Sensing Mediated Phenotypes by Berberine in Pseudomonas Aeruginosa and Salmonella Typhimurium. RSC Adv. 2018, 8, 36133–36141. [Google Scholar] [CrossRef] [PubMed]
- Bernatoniene, J.; Kopustinskiene, D.M. The Role of Catechins in Cellular Responses to Oxidative Stress. Molecules 2018, 23, 965. [Google Scholar] [CrossRef] [PubMed]
- Latos-Brozio, M.; Masek, A. Natural Polymeric Compound Based on High Thermal Stability Catechin from Green Tea. Biomolecules 2020, 10, 1191. [Google Scholar] [CrossRef]
- Gadkari, P.V.; Balaraman, M. Catechins: Sources, Extraction and Encapsulation: A Review. Food Bioprod. Process. 2015, 93, 122–138. [Google Scholar] [CrossRef]
- Pedro, A.C.; Maciel, G.M.; Rampazzo Ribeiro, V.; Haminiuk, C.W.I. Fundamental and Applied Aspects of Catechins from Different Sources: A Review. Int. J. Food Sci. Technol. 2020, 55, 429–442. [Google Scholar] [CrossRef]
- Yilmaz, Y. Novel Uses of Catechins in Foods. Trends Food Sci. Technol. 2006, 17, 64–71. [Google Scholar] [CrossRef]
- Gomes, F.M.S.; da Cunha Xavier, J.; dos Santos, J.F.S.; de Matos, Y.M.L.S.; Tintino, S.R.; de Freitas, T.S.; Coutinho, H.D.M. Evaluation of Antibacterial and Modifying Action of Catechin Antibiotics in Resistant Strains. Microb. Pathog. 2018, 115, 175–178. [Google Scholar] [CrossRef]
- Molina-Hernández, J.B.; Scroccarello, A.; Della Pelle, F.; De Flaviis, R.; Compagnone, D.; Del Carlo, M.; Paparella, A.; Chaves Lόpez, C. Synergistic Antifungal Activity of Catechin and Silver Nanoparticles on Aspergillus Niger Isolated from Coffee Seeds. LWT 2022, 169. [Google Scholar] [CrossRef]
- Xu, J.; Xu, Z.; Zheng, W. A Review of the Antiviral Role of Green Tea Catechins. Molecules 2017, 22, 1337. [Google Scholar] [CrossRef]
- Yu, Y.; Deng, Y.; Lu, B.M.; Liu, Y.X.; Li, J.; Bao, J.K. Green Tea Catechins: A Fresh Flavor to Anticancer Therapy. Apoptosis 2014, 19, 1–18. [Google Scholar] [CrossRef]
- Geetha, T.; Garg, A.; Chopra, K.; Pal Kaur, I. Delineation of Antimutagenic Activity of Catechin, Epicatechin and Green Tea Extract. Mutat. Res. Fundam. Mol. Mech. Mutagen. 2004, 556, 65–74. [Google Scholar] [CrossRef]
- Delgado, L.; Fernandes, I.; González-Manzano, S.; De Freitas, V.; Mateus, N.; Santos-Buelga, C. Anti-Proliferative Effects of Quercetin and Catechin Metabolites. Food Funct. 2014, 5, 797–803. [Google Scholar] [CrossRef] [PubMed]
- Zanwar, A.A.; Badole, S.L.; Shende, P.S.; Hegde, M.V.; Bodhankar, S.L. Antioxidant Role of Catechin in Health and Disease; Elsevier Inc.: Amsterdam, The Netherlands, 2013; Volume 1, ISBN 9780123984562. [Google Scholar]
- Nakanishi, T.; Mukai, K.; Yumoto, H.; Hirao, K.; Hosokawa, Y.; Matsuo, T. Anti-Inflammatory Effect of Catechin on Cultured Human Dental Pulp Cells Affected by Bacteria-Derived Factors. Eur. J. Oral Sci. 2010, 118, 145–150. [Google Scholar] [CrossRef]
- Yoshino, S.; Mitoma, T.; Tsuruta, K.; Todo, H.; Sugibayashi, K. Effect of Emulsification on the Skin Permeation and UV Protection of Catechin. Pharm. Dev. Technol. 2014, 19, 395–400. [Google Scholar] [CrossRef]
- Pervin, M.; Unno, K.; Ohishi, T.; Tanabe, H.; Miyoshi, N.; Nakamura, Y. Beneficial Effects of Green Tea Catechins on Neurodegenerative Diseases. Molecules 2018, 23, 1297. [Google Scholar] [CrossRef] [PubMed]
- Veiko, A.G.; Olchowik-Grabarek, E.; Sekowski, S.; Roszkowska, A.; Lapshina, E.A.; Dobrzynska, I.; Zamaraeva, M.; Zavodnik, I.B. Antimicrobial Activity of Quercetin, Naringenin and Catechin: Flavonoids Inhibit Staphylococcus aureus-Induced Hemolysis and Modify Membranes of Bacteria and Erythrocytes. Molecules 2023, 28, 1252. [Google Scholar] [CrossRef] [PubMed]
- Miklasińska, M.; Kępa, M.; Wojtyczka, R.D.; Idzik, D.; Dziedzic, A.; Wąsik, T.J. Catechin Hydrate Augments the Antibacterial Action of Selected Antibiotics against Staphylococcus aureus Clinical Strains. Molecules 2016, 21, 244. [Google Scholar] [CrossRef] [PubMed]
- Qin, R.; Xiao, K.; Li, B.; Jiang, W.; Peng, W.; Zheng, J.; Zhou, H. The Combination of Catechin and Epicatechin Gallate from Fructus Crataegi Potentiates β-Lactam Antibiotics against Methicillin-Resistant Staphylococcus aureus (MRSA) In Vitro and In Vivo. Int. J. Mol. Sci. 2013, 14, 1802–1821. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Venegas, G.; Gómez-Mora, J.A.; Meraz-Rodríguez, M.A.; Flores-Sánchez, M.A.; Ortiz-Miranda, L.F. Effect of Flavonoids on Antimicrobial Activity of Microorganisms Present in Dental Plaque. Heliyon 2019, 5, e03013. [Google Scholar] [CrossRef] [PubMed]
- Stanislaus, N.N.; Ibrahim, S.M.; Usman, P.U.; Sa’adiya, H.H.; Garba, M.M.; Sa’ad Toyin, A.; Taibat Moji, B.-O.; Ndifor, A.R.; Osas, E.G.; Oyetunji, S.A.; et al. Antimicrobial and Antioxidant Activity of Catechin-3-o-Rhamnoside Isolated from the Stem Bark of Lannea Kerstingii Engl. and K. Krause (Anacardiaceae). Pak. J. Pharm. Sci. 2021, 34, 629–634. [Google Scholar] [PubMed]
- Gonçalves, I.; Matamá, T.; Cavaco-Paulo, A.; Silva, C. Laccase Coating of Catheters with Poly(Catechin) for Biofilm Reduction. Biocatal. Biotransformation 2014, 32, 2–12. [Google Scholar] [CrossRef]
- Kurnia, D.; Ramadhanty, Z.F.; Ardani, A.M.; Zainuddin, A.; Dharsono, H.D.A.; Satari, M.H. Bio-Mechanism of Catechin as Pheromone Signal Inhibitor: Prediction of Antibacterial Agent Action Mode by In Vitro and In Silico Study. Molecules 2021, 26, 6381. [Google Scholar] [CrossRef]
- Qabaha, K.; Abbadi, J.; Yaghmour, R.; Hijawi, T.; Naser, S.A.; Al-Rimawi, F. Unveiling the Antibacterial and Antioxidant Potential of Hedera Helix Leaf Extracts: Recent Findings. Can. J. Physiol. Pharmacol. 2023, 102. [Google Scholar] [CrossRef]
- Albayrak, S.; Aksoy, A. Comparative Analysis of Phenolic Compositions and Biological Activities of Three Endemic Teucrium L. (Lamiaceae) Species from Turkey. An. Acad. Bras. Cienc. 2023, 95, e20220184. [Google Scholar] [CrossRef]
- Alqahtani, A.; Marrez, D.A.; Aleraky, M.; Fagir, N.A.; Alqahtani, O.; Othman, S.; El Raey, M.A.; Attia, H.G. Characterization and Isolation of the Major Biologically Active Metabolites Isolated from Ficus Retusa and Their Synergistic Effect with Tetracycline against Certain Pathogenic-Resistant Bacteria. Pharmaceuticals 2022, 15, 1473. [Google Scholar] [CrossRef]
- Kuma, D.N.; Boye, A.; Kwakye-Nuako, G.; Boakye, Y.D.; Addo, J.K.; Asiamah, E.A.; Aboagye, E.A.; Martey, O.; Essuman, M.A.; Atsu Barku, V.Y. Wound Healing Properties and Antimicrobial Effects of Parkia Clappertoniana Keay Fruit Husk Extract in a Rat Excisional Wound Model. BioMed Res. Int. 2022, 2022, 9709365. [Google Scholar] [CrossRef]
- Magangana, T.P.; Makunga, N.P.; la Grange, C.; Stander, M.A.; Fawole, O.A.; Opara, U.L. Blanching Pre-Treatment Promotes High Yields, Bioactive Compounds, Antioxidants, Enzyme Inactivation and Antibacterial Activity of “Wonderful” Pomegranate Peel Extracts at Three Different Harvest Maturities. Antioxidants 2021, 10, 1119. [Google Scholar] [CrossRef]
- Ibitoye, O.B.; Ajiboye, T.O. (+)-Catechin Potentiates the Oxidative Response of Acinetobacter Baumannii to Quinolone-Based Antibiotics. Microb. Pathog. 2019, 127, 239–245. [Google Scholar] [CrossRef]
- Yang, M.-J.; Hung, Y.-A.; Wong, T.-W.; Lee, N.-Y.; Yuann, J.-M.P.; Huang, S.-T.; Wu, C.-Y.; Chen, I.-Z.; Liang, J.-Y. Effects of Blue-Light-Induced Free Radical Formation from Catechin Hydrate on the Inactivation of Acinetobacter Baumannii, Including a Carbapenem-Resistant Strain. Molecules 2018, 23, 1631. [Google Scholar] [CrossRef]
- Mumtaz, L.; Farid, A.; Yousef Alomar, S.; Ahmad, N.; Nawaz, A.; Andleeb, S.; Amin, A. Assesment of Polyphenolic Compounds against Biofilms Produced by Clinical Acinetobacter Baumannii Strains Using In Silico and In Vitro Models. Saudi J. Biol. Sci. 2023, 30, 103743. [Google Scholar] [CrossRef] [PubMed]
- Garcia, J.; Rodrigues, F.; Castro, F.; Aires, A.; Marques, G.; Saavedra, M.J. Antimicrobial, Antibiofilm, and Antioxidant Properties of Boletus Edulis and Neoboletus Luridiformis against Multidrug-Resistant ESKAPE Pathogens. Front. Nutr. 2022, 8. [Google Scholar] [CrossRef] [PubMed]
- Abdel Bar, F.M.; Alossaimi, M.A.; Elekhnawy, E.; Alzeer, M.A.A.; Abo Kamer, A.; Moglad, E.; ElNaggar, M.H. Anti-Quorum Sensing and Anti-Biofilm Activity of Pelargonium × Hortorum Root Extract against Pseudomonas Aeruginosa: Combinatorial Effect of Catechin and Gallic Acid. Molecules 2022, 27, 7841. [Google Scholar] [CrossRef] [PubMed]
- Kalirajan, C.; Palanisamy, T. Bioengineered Hybrid Collagen Scaffold Tethered with Silver-Catechin Nanocomposite Modulates Angiogenesis and TGF-β Toward Scarless Healing in Chronic Deep Second Degree Infected Burns. Adv. Healthc. Mater. 2020, 9, e2000247. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Xiao, X.; Li, K.; Li, X.; Yu, K.; Liao, X.; Shi, B. Prevention of Bacterial Colonization Based on Self-Assembled Metal–Phenolic Nanocoating from Rare-Earth Ions and Catechin. ACS Appl. Mater. Interfaces 2020, 12, 22237–22245. [Google Scholar] [CrossRef]
- Ajiboye, T.O.; Aliyu, M.; Isiaka, I.; Haliru, F.Z.; Ibitoye, O.B.; Uwazie, J.N.; Muritala, H.F.; Bello, S.A.; Yusuf, I.I.; Mohammed, A.O. Contribution of Reactive Oxygen Species to (+)-Catechin-Mediated Bacterial Lethality. Chem. Biol. Interact. 2016, 258, 276–287. [Google Scholar] [CrossRef]
- Bernal-Mercado, A.; Vazquez-Armenta, F.; Tapia-Rodriguez, M.; Islas-Osuna, M.; Mata-Haro, V.; Gonzalez-Aguilar, G.; Lopez-Zavala, A.; Ayala-Zavala, J. Comparison of Single and Combined Use of Catechin, Protocatechuic, and Vanillic Acids as Antioxidant and Antibacterial Agents against Uropathogenic Escherichia coli at Planktonic and Biofilm Levels. Molecules 2018, 23, 2813. [Google Scholar] [CrossRef]
- Bernal-Mercado, A.T.; Gutierrez-Pacheco, M.M.; Encinas-Basurto, D.; Mata-Haro, V.; Lopez-Zavala, A.A.; Islas-Osuna, M.A.; Gonzalez-Aguilar, G.A.; Ayala-Zavala, J.F. Synergistic Mode of Action of Catechin, Vanillic and Protocatechuic Acids to Inhibit the Adhesion of Uropathogenic Escherichia coli on Silicone Surfaces. J. Appl. Microbiol. 2020, 128, 387–400. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Gómez, R.; Toledo-Araya, H.; López-Solís, R.; Obreque-Slier, E. Combined Effect of Gallic Acid and Catechin against Escherichia coli. LWT-Food Sci. Technol. 2014, 59, 896–900. [Google Scholar] [CrossRef]
- Kim, J.-W.; Kim, C.-Y.; Kim, J.-H.; Jeong, J.-S.; Lim, J.-O.; Ko, J.-W.; Kim, T.-W. Prophylactic Catechin-Rich Green Tea Extract Treatment Ameliorates Pathogenic Enterotoxic Escherichia coli-Induced Colitis. Pathogens 2021, 10, 1573. [Google Scholar] [CrossRef]
- Etsè, K.D.; Aïdam, A.V.; Melin, C.; Blanc, N.; Oudin, A.; Courdavault, V.; Creche, J.; Lanoue, A. Optimized Genetic Transformation of Zanthoxylum Zanthoxyloides by Agrobacterium Rhizogenes and the Production of Chelerythrine and Skimmiamine in Hairy Root Cultures. Eng. Life Sci. 2014, 14, 95–99. [Google Scholar] [CrossRef]
- Heng, W.S.; Cheah, S.-C. Chelerythrine Chloride Downregulates β-Catenin and Inhibits Stem Cell Properties of Non-Small Cell Lung Carcinoma. Molecules 2020, 25, 224. [Google Scholar] [CrossRef]
- Chen, N.; Qi, Y.; Ma, X.; Xiao, X.; Liu, Q.; Xia, T.; Xiang, J.; Zeng, J.; Tang, J. Rediscovery of Traditional Plant Medicine: An Underestimated Anticancer Drug of Chelerythrine. Front. Pharmacol. 2022, 13, 906301. [Google Scholar] [CrossRef] [PubMed]
- He, N.; Wang, P.; Wang, P.; Ma, C.; Kang, W. Antibacterial Mechanism of Chelerythrine Isolated from Root of Toddalia Asiatica (Linn) Lam. BMC Complement. Altern. Med. 2018, 18, 261. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.J.; Miao, F.; Yao, Y.; Cao, F.J.; Yang, R.; Ma, Y.N.; Qin, B.F.; Zhou, L. In Vitro Antifungal Activity of Sanguinarine and Chelerythrine Derivatives against Phytopathogenic Fungi. Molecules 2012, 17, 13026–13035. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Pan, Y.; Zhang, G.; Wu, Y.; Zhong, W.; Chu, C.; Qian, Y.; Zhu, G. Chelerythrine Inhibits Human Hepatocellular Carcinoma Metastasis In Vitro. Biol. Pharm. Bull. 2018, 41, 36–46. [Google Scholar] [CrossRef]
- Qian, W.; Sun, Z.; Fu, Y.; Yang, M.; Wang, T.; Li, Y. Efficacy of Chelerythrine against Dual-Species Biofilms of Staphylococcus aureus and Staphylococcus Lugdunensis. 3 Biotech 2020, 10, 427. [Google Scholar] [CrossRef] [PubMed]
- Móricz, Á.M.; Fornal, E.; Jesionek, W.; Majer-Dziedzic, B.; Choma, I.M. Effect-Directed Isolation and Identification of Antibacterial Chelidonium Majus L. Alkaloids. Chromatographia 2015, 78, 707–716. [Google Scholar] [CrossRef]
- Tavares, L.d.C.; Zanon, G.; Weber, A.D.; Neto, A.T.; Mostardeiro, C.P.; Da Cruz, I.B.M.; Oliveira, R.M.; Ilha, V.; Dalcol, I.I.; Morel, A.F. Structure-Activity Relationship of Benzophenanthridine Alkaloids from Zanthoxylum Rhoifolium Having Antimicrobial Activity. PLoS ONE 2014, 9, e97000. [Google Scholar] [CrossRef] [PubMed]
- Zielińska, S.; Wójciak-Kosior, M.; Dziągwa-Becker, M.; Gleńsk, M.; Sowa, I.; Fijałkowski, K.; Rurańska-Smutnicka, D.; Matkowski, A.; Junka, A. The Activity of Isoquinoline Alkaloids and Extracts from Chelidonium Majus against Pathogenic Bacteria and Candida sp. Toxins 2019, 11, 406. [Google Scholar] [CrossRef] [PubMed]
- Qian, W.-D.; Huang, J.; Zhang, J.-N.; Li, X.-C.; Kong, Y.; Wang, T.; Li, Y.-D. Antimicrobial and Antibiofilm Activities and Mechanism of Action of Chelerythrine against Carbapenem-Resistant Serratia Marcescens In Vitro. Microb. Drug Resist. 2021, 27, 1105–1116. [Google Scholar] [CrossRef]
- Friedman, M. Chemistry, Antimicrobial Mechanisms, and Antibiotic Activities of Cinnamaldehyde against Pathogenic Bacteria in Animal Feeds and Human Foods. J. Agric. Food Chem. 2017, 65, 10406–10423. [Google Scholar] [CrossRef]
- Doyle, A.A.; Stephens, J.C. A Review of Cinnamaldehyde and Its Derivatives as Antibacterial Agents. Fitoterapia 2019, 139, 104405. [Google Scholar] [CrossRef]
- Thirapanmethee, K.; Kanathum, P.; Khuntayaporn, P.; Huayhongthong, S.; Surassmo, S.; Chomnawang, M.T. Cinnamaldehyde: A Plant-Derived Antimicrobial for Overcoming Multidrug-Resistant Acinetobacter Baumannii Infection. Eur. J. Integr. Med. 2021, 48, 101376. [Google Scholar] [CrossRef]
- Ibi, A.A.; Kyuka, C.K. Sources, Extraction and Biological Activities of Cinnamaldehyde. Trends Pharm. Sci. 2022, 2022, 263–282. [Google Scholar] [CrossRef]
- Arianie, L.; Eko Hermanto, F.; Dhiaul Iftitah, E.; Warsito, W.; Widodo, N. Novel Antimalarial Drug Screening Based on Methyl Eugenol, Cinnamaldehyde, and Thiosemicarbazone with Cysteine Protease Inhibition: In Silico Molecular Docking, Molecular Dynamics, and ADMET Studies. J. Pure Appl. Chem. Res. 2022, 11, 102–112. [Google Scholar] [CrossRef]
- Chang, S.; Qin, D.; Wang, L.; Zhang, M.; Yan, R.; Zhao, C. Preparation of Novel Cinnamaldehyde Derivative–BSA Nanoparticles with High Stability, Good Cell Penetrating Ability, and Promising Anticancer Activity. Colloids Surf. A Physicochem. Eng. Asp. 2021, 624, 126765. [Google Scholar] [CrossRef]
- Sun, Q.; Li, J.; Sun, Y.; Chen, Q.; Zhang, L.; Le, T. The Antifungal Effects of Cinnamaldehyde against Aspergillus Niger and Its Application in Bread Preservation. Food Chem. 2020, 317, 126405. [Google Scholar] [CrossRef]
- Goswami, A.; Rahman, A. Antiviral Activity of (Ε)-Cinnamaldehyde Revisited with Nanoscience Tools. Nat. Preced. 2010, 1–4. [Google Scholar] [CrossRef]
- Firmino, D.F.; Cavalcante, T.T.A.; Gomes, G.A.; Firmino, N.C.S.; Rosa, L.D.; De Carvalho, M.G.; Catunda, F.E.A. Antibacterial and Antibiofilm Activities of Cinnamomum sp. Essential Oil and Cinnamaldehyde: Antimicrobial Activities. Sci. World J. 2018, 2018, 7405736. [Google Scholar] [CrossRef] [PubMed]
- Subash Babu, P.; Prabuseenivasan, S.; Ignacimuthu, S. Cinnamaldehyde-A Potential Antidiabetic Agent. Phytomedicine 2007, 14, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Subash-Babu, P.; Alshatwi, A.A.; Ignacimuthu, S. Beneficial Antioxidative and Antiperoxidative Effect of Cinnamaldehyde Protect Streptozotocin-Induced Pancreatic β-Cells Damage in Wistar Rats. Biomol. Ther. 2014, 22, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.X.; Zhong, S.; Meng, X.B.; Zheng, N.Y.; Zhang, P.; Wang, Y.; Qin, L.; Wang, X.L. Cinnamaldehyde Inhibits Inflammation of Human Synoviocyte Cells through Regulation of JAK/STAT Pathway and Ameliorates Collagen-Induced Arthritis in Rats. J. Pharmacol. Exp. Ther. 2020, 373, 302–310. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhou, W.; Zhang, W.; Yang, A.; Liu, Y.; Jiang, Y.; Huang, S.; Su, J. Inhibitory Effects of Citral, Cinnamaldehyde, and Tea Polyphenols on Mixed Biofilm Formation by Foodborne Staphylococcus aureus and Salmonella Enteritidis. J. Food Prot. 2014, 77, 927–933. [Google Scholar] [CrossRef] [PubMed]
- Shen, S.; Zhang, T.; Yuan, Y.; Lin, S.; Xu, J.; Ye, H. Effects of Cinnamaldehyde on Escherichia coli and Staphylococcus aureus Membrane. Food Control 2015, 47, 196–202. [Google Scholar] [CrossRef]
- Ferro, T.A.F.; Araújo, J.M.M.; dos Santos Pinto, B.L.; dos Santos, J.S.; Souza, E.B.; da Silva, B.L.R.; Colares, V.L.P.; Novais, T.M.G.; Filho, C.M.B.; Struve, C.; et al. Cinnamaldehyde Inhibits Staphylococcus aureus Virulence Factors and Protects against Infection in a Galleria Mellonella Model. Front. Microbiol. 2016, 7, 2052. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, I.F.S.; Araújo, L.G.; Assunção, R.G.; Dutra, I.L.; Nascimento, J.R.; Rego, F.S.; Rolim, C.S.; Alves, L.S.R.; Frazão, M.A.; Cadete, S.F.; et al. Cinnamaldehyde Increases the Survival of Mice Submitted to Sepsis Induced by Extraintestinal Pathogenic Escherichia coli. Antibiotics 2022, 11, 364. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Kang, O.-H.; Kwon, D.-Y. Trans-Cinnamaldehyde Exhibits Synergy with Conventional Antibiotic against Methicillin-Resistant Staphylococcus aureus. Int. J. Mol. Sci. 2021, 22, 2752. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Zhang, J.; Sun, Y.; Xu, J.; Yu, Z.; Huang, L.; Yao, W.; Xie, Y. Inactivation Action of Ultrasound-Assisted Cinnamaldehyde on Planktonic and Biofilm Methicillin-Resistant Staphylococcus aureus and Its Application in Beef System. Food Biosci. 2023, 55, 103031. [Google Scholar] [CrossRef]
- Shi, C.; Zhang, X.; Zhao, X.; Meng, R.; Liu, Z.; Chen, X.; Guo, N. Synergistic Interactions of Nisin in Combination with Cinnamaldehyde against Staphylococcus aureus in Pasteurized Milk. Food Control 2017, 71, 10–16. [Google Scholar] [CrossRef]
- Kim, Y.; Kim, S.; Cho, K.-H.; Lee, J.-H.; Lee, J. Antibiofilm Activities of Cinnamaldehyde Analogs against Uropathogenic Escherichia coli and Staphylococcus aureus. Int. J. Mol. Sci. 2022, 23, 7225. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Lin, Q.; Sheng, M.; Ding, T.; Li, B.; Gao, Y.; Tan, Y. Antibiofilm Effect of Cinnamaldehyde-Chitosan Nanoparticles against the Biofilm of Staphylococcus aureus. Antibiotics 2022, 11, 1403. [Google Scholar] [CrossRef] [PubMed]
- Albano, M.; Crulhas, B.P.; Alves, F.C.B.; Pereira, A.F.M.; Andrade, B.F.M.T.; Barbosa, L.N.; Furlanetto, A.; Lyra, L.P.d.S.; Rall, V.L.M.; Júnior, A.F. Antibacterial and Anti-Biofilm Activities of Cinnamaldehyde against S. epidermidis. Microb. Pathog. 2019, 126, 231–238. [Google Scholar] [CrossRef]
- Sharma, G.; Raturi, K.; Dang, S.; Gupta, S.; Gabrani, R. Inhibitory Effect of Cinnamaldehyde Alone and in Combination with Thymol, Eugenol and Thymoquinone against Staphylococcus Epidermidis. J. Herb. Med. 2017, 9, 68–73. [Google Scholar] [CrossRef]
- Ali, I.A.A.; Matinlinna, J.P.; Lévesque, C.M.; Neelakantan, P. Trans-Cinnamaldehyde Attenuates Enterococcus Faecalis Virulence and Inhibits Biofilm Formation. Antibiotics 2021, 10, 702. [Google Scholar] [CrossRef] [PubMed]
- Akshaya, B.S.; Premraj, K.; Iswarya, C.; Muthusamy, S.; Ibrahim, H.-I.M.; Khalil, H.E.; Ashokkumar, V.; Vickram, S.; Senthil Kumar, V.; Palanisamy, S.; et al. Cinnamaldehyde Inhibits Enterococcus Faecalis Biofilm Formation and Promotes Clearance of Its Colonization by Modulation of Phagocytes In Vitro. Microb. Pathog. 2023, 181, 106157. [Google Scholar] [CrossRef] [PubMed]
- Ali, I.A.A.; Cheung, B.P.K.; Matinlinna, J.; Lévesque, C.M.; Neelakantan, P. Trans-Cinnamaldehyde Potently Kills Enterococcus Faecalis Biofilm Cells and Prevents Biofilm Recovery. Microb. Pathog. 2020, 149, 104482. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.; Kalimuthu, S.; Zhang, C.; Ali, I.A.A.; Neelakantan, P. Trans-Cinnamaldehyde-Biosurfactant Complex as a Potent Agent against Enterococcus Faecalis Biofilms. Pharmaceutics 2022, 14, 2355. [Google Scholar] [CrossRef] [PubMed]
- Vaz, M.S.M.; de Almeida de Souza, G.H.; dos Santos Radai, J.A.; Fraga, T.L.; de Oliveira, G.G.; Wender, H.; da Silva, K.E.; Simionatto, S. Antimicrobial Activity of Cinnamaldehyde against Multidrug-Resistant Klebsiella Pneumoniae: An In Vitro and In Vivo Study. Braz. J. Microbiol. 2023, 54, 1655–1664. [Google Scholar] [CrossRef] [PubMed]
- Dhara, L.; Tripathi, A. Cinnamaldehyde: A Compound with Antimicrobial and Synergistic Activity against ESBL-Producing Quinolone-Resistant Pathogenic Enterobacteriaceae. Eur. J. Clin. Microbiol. Infect. Dis. 2020, 39, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, S.H.; Salem, D.; Azmy, M.; Fam, N.S. Antibacterial and Antibiofilm Activity of Cinnamaldehyde against Carbapenem-Resistant Acinetobacter Baumannii in Egypt: In Vitro Study. J. Appl. Pharm. Sci. 2018, 8, 151–156. [Google Scholar] [CrossRef]
- Chai, W.C.; Whittall, J.J.; Polyak, S.W.; Foo, K.; Li, X.; Dutschke, C.J.; Ogunniyi, A.D.; Ma, S.; Sykes, M.J.; Semple, S.J.; et al. Cinnamaldehyde Derivatives Act as Antimicrobial Agents against Acinetobacter Baumannii through the Inhibition of Cell Division. Front. Microbiol. 2022, 13, 967949. [Google Scholar] [CrossRef]
- Karumathil, D.P.; Nair, M.S.; Gaffney, J.; Kollanoor-Johny, A.; Venkitanarayanan, K. Trans-Cinnamaldehyde and Eugenol Increase Acinetobacter Baumannii Sensitivity to Beta-Lactam Antibiotics. Front. Microbiol. 2018, 9, 1011. [Google Scholar] [CrossRef]
- Karumathil, D.P.; Surendran-Nair, M.; Venkitanarayanan, K. Efficacy of Trans-Cinnamaldehyde and Eugenol in Reducing Acinetobacter Baumannii Adhesion to and Invasion of Human Keratinocytes and Controlling Wound Infection In Vitro. Phytother. Res. 2016, 30, 2053–2059. [Google Scholar] [CrossRef] [PubMed]
- Topa, S.H.; Palombo, E.A.; Kingshott, P.; Blackall, L.L. Activity of Cinnamaldehyde on Quorum Sensing and Biofilm Susceptibility to Antibiotics in Pseudomonas Aeruginosa. Microorganisms 2020, 8, 455. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Yang, H.; Meng, X.; Su, R.; Cheng, S.; Wang, H.; Bai, X.; Guo, D.; Lü, X.; Xia, X.; et al. Inhibitory Effects of Trans-Cinnamaldehyde against Pseudomonas Aeruginosa Biofilm Formation. Foodborne Pathog. Dis. 2023, 20, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Chadha, J.; Ravi; Singh, J.; Chhibber, S.; Harjai, K. Gentamicin Augments the Quorum Quenching Potential of Cinnamaldehyde In Vitro and Protects Caenorhabditis Elegans from Pseudomonas Aeruginosa Infection. Front. Cell. Infect. Microbiol. 2022, 12. [Google Scholar] [CrossRef] [PubMed]
- Subhaswaraj, P.; Barik, S.; Macha, C.; Chiranjeevi, P.V.; Siddhardha, B. Anti Quorum Sensing and Anti Biofilm Efficacy of Cinnamaldehyde Encapsulated Chitosan Nanoparticles against Pseudomonas Aeruginosa PAO1. LWT 2018, 97, 752–759. [Google Scholar] [CrossRef]
- Mohammadzamani, Z.; Khorshidi, A.; Khaledi, A.; Shakerimoghaddam, A.; Moosavi, G.A.; Piroozmand, A. Inhibitory Effects of Cinnamaldehyde, Carvacrol, and Honey on the Expression of ExoS and AmpC Genes in Multidrug-Resistant Pseudomonas Aeruginosa Isolated from Burn Wound Infections. Microb. Pathog. 2020, 140, 103946. [Google Scholar] [CrossRef] [PubMed]
- Ferro, T.A.F.; Souza, E.B.; Suarez, M.A.M.; Rodrigues, J.F.S.; Pereira, D.M.S.; Mendes, S.J.F.; Gonzaga, L.F.; Machado, M.C.A.M.; Bomfim, M.R.Q.; Calixto, J.B.; et al. Topical Application of Cinnamaldehyde Promotes Faster Healing of Skin Wounds Infected with Pseudomonas Aeruginosa. Molecules 2019, 24, 1627. [Google Scholar] [CrossRef] [PubMed]
- Tetard, A.; Zedet, A.; Girard, C.; Plésiat, P.; Llanes, C. Cinnamaldehyde Induces Expression of Efflux Pumps and Multidrug Resistance in Pseudomonas Aeruginosa. Antimicrob. Agents Chemother. 2019, 63. [Google Scholar] [CrossRef] [PubMed]
- Tetard, A.; Gaillot, S.; Dubois, E.; Aarras, S.; Valot, B.; Phan, G.; Plésiat, P.; Llanes, C. Exposure of Pseudomonas Aeruginosa to Cinnamaldehyde Selects Multidrug Resistant Mutants. Antibiotics 2022, 11, 1790. [Google Scholar] [CrossRef]
- He, T.-F.; Zhang, Z.-H.; Zeng, X.-A.; Wang, L.-H.; Brennan, C.S. Determination of Membrane Disruption and Genomic DNA Binding of Cinnamaldehyde to Escherichia coli by Use of Microbiological and Spectroscopic Techniques. J. Photochem. Photobiol. B Biol. 2018, 178, 623–630. [Google Scholar] [CrossRef]
- Visvalingam, J.; Palaniappan, K.; Holley, R.A. In Vitro Enhancement of Antibiotic Susceptibility of Drug Resistant Escherichia coli by Cinnamaldehyde. Food Control 2017, 79, 288–291. [Google Scholar] [CrossRef]
- Pereira, W.A.; Pereira, C.D.S.; Assunção, R.G.; da Silva, I.S.C.; Rego, F.S.; Alves, L.S.R.; Santos, J.S.; Nogueira, F.J.R.; Zagmignan, A.; Thomsen, T.T.; et al. New Insights into the Antimicrobial Action of Cinnamaldehyde towards Escherichia coli and Its Effects on Intestinal Colonization of Mice. Biomolecules 2021, 11, 302. [Google Scholar] [CrossRef]
- Narayanan, A.; Muyyarikkandy, M.S.; Mooyottu, S.; Venkitanarayanan, K.; Amalaradjou, M.A.R. Oral Supplementation of Trans -Cinnamaldehyde Reduces Uropathogenic Escherichia coli Colonization in a Mouse Model. Lett. Appl. Microbiol. 2017, 64, 192–197. [Google Scholar] [CrossRef]
- Kot, B.; Wicha, J.; Piechota, M.; Wolska, K.; Grużewska, A. Antibiofilm Activity of Trans-Cinnamaldehyde, p-Coumaric, and Ferulic Acids on Uropathogenic Escherichia coli. Turk. J. Med. Sci. 2015, 45, 919–924. [Google Scholar] [CrossRef] [PubMed]
- Morgaan, H.A.; Omar, H.M.G.; Zakaria, A.S.; Mohamed, N.M. Repurposing Carvacrol, Cinnamaldehyde, and Eugenol as Potential Anti-Quorum Sensing Agents against Uropathogenic Escherichia coli Isolates in Alexandria, Egypt. BMC Microbiol. 2023, 23, 300. [Google Scholar] [CrossRef]
- Prasastha Ram, V.; Yasur, J.; Abishad, P.; Unni, V.; Purushottam Gourkhede, D.; Nishanth, M.A.D.; Niveditha, P.; Vergis, J.; Singh Malik, S.V.; Kullaiah, B.; et al. Antimicrobial Efficacy of Green Synthesized Nanosilver with Entrapped Cinnamaldehyde against Multi-Drug-Resistant Enteroaggregative Escherichia coli in Galleria Mellonella. Pharmaceutics 2022, 14, 1924. [Google Scholar] [CrossRef]
- Chuang, S.; Sheen, S.; Sommers, C.H.; Sheen, L. Modeling the Effect of Simultaneous Use of Allyl Isothiocyanate and Cinnamaldehyde on High Hydrostatic Pressure Inactivation of Uropathogenic and Shiga Toxin-producing Escherichia coli in Ground Chicken. J. Sci. Food Agric. 2021, 101, 1193–1201. [Google Scholar] [CrossRef] [PubMed]
- Sheen, S.; Huang, C.; Ramos, R.; Chien, S.; Scullen, O.J.; Sommers, C. Lethality Prediction for Escherichia coli O157:H7 and Uropathogenic E. coli in Ground Chicken Treated with High Pressure Processing and Trans-Cinnamaldehyde. J. Food Sci. 2018, 83, 740–749. [Google Scholar] [CrossRef]
- Zhu, L.; Olsen, C.; McHugh, T.; Friedman, M.; Levin, C.E.; Jaroni, D.; Ravishankar, S. Edible Films Containing Carvacrol and Cinnamaldehyde Inactivate Escherichia coli O157:H7 on Organic Leafy Greens in Sealed Plastic Bags. J. Food Saf. 2020, 40. [Google Scholar] [CrossRef]
- Olszewska, M.A.; Gędas, A.; Simões, M. The Effects of Eugenol, Trans-Cinnamaldehyde, Citronellol, and Terpineol on Escherichia coli Biofilm Control as Assessed by Culture-Dependent and -Independent Methods. Molecules 2020, 25, 2641. [Google Scholar] [CrossRef]
- Yossa, N.; Patel, J.; Macarisin, D.; Millner, P.; Murphy, C.; Bauchan, G.; Lo, Y.M. Antibacterial Activity of Cinnamaldehyde and Sporan against Escherichia coli O157:H7 and Salmonella. J. Food Process. Preserv. 2014, 38, 749–757. [Google Scholar] [CrossRef]
- Pang, D.; Huang, Z.; Li, Q.; Wang, E.; Liao, S.; Li, E.; Zou, Y.; Wang, W. Antibacterial Mechanism of Cinnamaldehyde: Modulation of Biosynthesis of Phosphatidylethanolamine and Phosphatidylglycerol in Staphylococcus aureus and Escherichia coli. J. Agric. Food Chem. 2021, 69, 13628–13636. [Google Scholar] [CrossRef]
- Kot, B.; Sytykiewicz, H.; Sprawka, I.; Witeska, M. Effect of Trans-Cinnamaldehyde on Methicillin-Resistant Staphylococcus aureus Biofilm Formation: Metabolic Activity Assessment and Analysis of the Biofilm-Associated Genes Expression. Int. J. Mol. Sci. 2019, 21, 102. [Google Scholar] [CrossRef]
- Andrade de Oliveira, E.L.; Da Silva Rodrigues, G.; Alves de Oliveira Filho, A.; Medeiros Medeiros, C.I. Atividade Anti-Staphylococcus aureus e Anti- Klebsiella Pneumoniae Do Cinamaldeído e Suas Interações Com ATP Sintase e FtsZ Através Do Docking Molecular. Rev. Colomb. Cienc. Químico-Farm. 2023, 52, 499–513. [Google Scholar] [CrossRef]
- Montagu, A.; Joly-Guillou, M.-L.; Rossines, E.; Cayon, J.; Kempf, M.; Saulnier, P. Stress Conditions Induced by Carvacrol and Cinnamaldehyde on Acinetobacter Baumannii. Front. Microbiol. 2016, 7, 1133. [Google Scholar] [CrossRef]
- Topa, S.H.; Subramoni, S.; Palombo, E.A.; Kingshott, P.; Rice, S.A.; Blackall, L.L. Cinnamaldehyde Disrupts Biofilm Formation and Swarming Motility of Pseudomonas Aeruginosa. Microbiology 2018, 164, 1087–1097. [Google Scholar] [CrossRef] [PubMed]
- He, T.-F.; Wang, L.-H.; Niu, D.; Wen, Q.; Zeng, X.-A. Cinnamaldehyde Inhibit Escherichia coli Associated with Membrane Disruption and Oxidative Damage. Arch. Microbiol. 2019, 201, 451–458. [Google Scholar] [CrossRef] [PubMed]
- Du, G.-F.; Yin, X.-F.; Yang, D.-H.; He, Q.-Y.; Sun, X. Proteomic Investigation of the Antibacterial Mechanism of Trans-Cinnamaldehyde against Escherichia coli. J. Proteome Res. 2021, 20, 2319–2328. [Google Scholar] [CrossRef] [PubMed]
- Aguilera-Carbo, A.F.; Augur, C.; Prado-Barragan, L.A.; Aguilar, C.N.; Favela-Torres, E. Extraction and Analysis of Ellagic Acid from Novel Complex Sources. Chem. Pap. 2008, 62, 440–444. [Google Scholar] [CrossRef]
- Sepúlveda, L.; Ascacio, A.; Rodríguez-Herrera, R.; Aguilera-Carbó, A.; Aguilar, C.N. Ellagic Acid: Biological Properties and Biotechnological Development for Production Processes. Afr. J. Biotechnol. 2011, 10, 4518–4523. [Google Scholar]
- Sharifi-Rad, J.; Quispe, C.; Castillo, C.M.S.; Caroca, R.; Lazo-Vélez, M.A.; Antonyak, H.; Polishchuk, A.; Lysiuk, R.; Oliinyk, P.; De Masi, L.; et al. Ellagic Acid: A Review on Its Natural Sources, Chemical Stability, and Therapeutic Potential. Oxid. Med. Cell. Longev. 2022, 2022, 3848084. [Google Scholar] [CrossRef] [PubMed]
- Evtyugin, D.D.; Magina, S.; Evtuguin, D.V. Recent Advances in the Production and Applications of Ellagic Acid and Its Derivatives. A Review. Molecules 2020, 25, 2745. [Google Scholar] [CrossRef] [PubMed]
- Priyadarsini, K.I.; Khopde, S.M.; Kumar, S.S.; Mohan, H. Free Radical Studies of Ellagic Acid, a Natural Phenolic Antioxidant. J. Agric. Food Chem. 2002, 50, 2200–2206. [Google Scholar] [CrossRef] [PubMed]
- Corbett, S.; Daniel, J.; Drayton, R.; Field, M.; Steinhardt, R.; Garrett, N. Evaluation of the Anti-Inflammatory Effects of Ellagic Acid. J. Perianesthesia Nurs. 2010, 25, 214–220. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Gao, X.; Li, H.; Li, Z.; Wang, X.; Zhang, L.; Wang, B.; Chen, X.; Meng, X.; Yu, J. Ellagic Acid Exerts Anti-Fibrotic Effects on Hypertrophic Scar Fibroblasts via Inhibition of TGF-Β1/Smad2/3 Pathway. Appl. Biol. Chem. 2021, 64, 67. [Google Scholar] [CrossRef]
- Zahin, M.; Ahmad, I.; Gupta, R.C.; Aqil, F. Punicalagin and Ellagic Acid Demonstrate Antimutagenic Activity and Inhibition of Benzo[a]Pyrene Induced DNA Adducts. BioMed Res. Int. 2014, 2014, 467465. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Choi, Y.J.; Kim, H.J. Determining the Effect of Ellagic Acid on the Proliferation and Migration of Pancreatic Cancer Cell Lines. Transl. Cancer Res. 2021, 10, 424–433. [Google Scholar] [CrossRef]
- Goudarzi, M.; Mombeini, M.A.; Fatemi, I.; Aminzadeh, A.; Kalantari, H.; Nesari, A.; Najafzadehvarzi, H.; Mehrzadi, S. Neuroprotective Effects of Ellagic Acid against Acrylamide-Induced Neurotoxicity in Rats. Neurol. Res. 2019, 41, 419–428. [Google Scholar] [CrossRef]
- Zhao, L.; Mehmood, A.; Soliman, M.M.; Iftikhar, A.; Iftikhar, M.; Aboelenin, S.M.; Wang, C. Protective Effects of Ellagic Acid against Alcoholic Liver Disease in Mice. Front. Nutr. 2021, 8, 4–6. [Google Scholar] [CrossRef]
- Altamimi, J.Z.; Alfaris, N.A.; Alshammari, G.M.; Alagal, R.I.; Aljabryn, D.H.; Aldera, H.; Alkhateeb, M.A.; Yahya, M.A. Ellagic Acid Protects against Diabetic Cardiomyopathy in Rats by Stimulating Cardiac Silent Information Regulator 1 Signaling. J. Physiol. Pharmacol. 2020, 71, 891–904. [Google Scholar] [CrossRef]
- Lukiswanto, B.S.; Miranti, A.; Sudjarwo, S.A.; Primarizky, H.; Yuniarti, W.M. Evaluation of Wound Healing Potential of Pomegranate (Punica Granatum) Whole Fruit Extract on Skin Burn Wound in Rats (Rattus Norvegicus). J. Adv. Vet. Anim. Res. 2019, 6, 202–207. [Google Scholar] [CrossRef]
- Wardhana, A.S.; Nirwana, I.; Budi, H.S.; Surboyo, M.D.C. Role of Hydroxyapatite and Ellagic Acid in the Osteogenesis. Eur. J. Dent. 2021, 15, 8–12. [Google Scholar] [CrossRef] [PubMed]
- Savic, I.M.; Jocic, E.; Nikolic, V.D.; Popsavin, M.M.; Rakic, S.J.; Savic-Gajic, I.M. The Effect of Complexation with Cyclodextrins on the Antioxidant and Antimicrobial Activity of Ellagic Acid. Pharm. Dev. Technol. 2019, 24, 410–418. [Google Scholar] [CrossRef] [PubMed]
- Acquadro, S.; Civra, A.; Cagliero, C.; Marengo, A.; Rittà, M.; Francese, R.; Sanna, C.; Bertea, C.; Sgorbini, B.; Lembo, D.; et al. Punica Granatum Leaf Ethanolic Extract and Ellagic Acid as Inhibitors of Zika Virus Infection. Planta Med. 2020, 86, 1363–1374. [Google Scholar] [CrossRef] [PubMed]
- Beshbishy, A.M.; Batiha, G.E.S.; Yokoyama, N.; Igarashi, I. Ellagic Acid Microspheres Restrict the Growth of Babesia and Theileria In Vitro and Babesia Microti In Vivo. Parasites Vectors 2019, 12, 269. [Google Scholar] [CrossRef]
- Tavares, W.d.S.; Martin-Pastor, M.; Tavares, A.G.; Sousa, F.F.O. Biopharmaceutical Activities Related to Ellagic Acid, Chitosan, and Zein and Their Improvement by Association. J. Food Sci. 2018, 83, 2970–2975. [Google Scholar] [CrossRef]
- Tavares, W.d.S.; Barreto, G.A.V.; Pinto, E.P.; Silva, P.G.d.B.; Sousa, F.F.O.d. Influence of Gelatin on the Functional Characteristics and Wound Healing Potential of Chitosan/Zein Films Loaded with Ellagic Acid Nanoparticles. J. Drug Deliv. Sci. Technol. 2023, 88, 104942. [Google Scholar] [CrossRef]
- Tavares, W.S.; Tavares-Júnior, A.G.; Otero-Espinar, F.J.; Martín-Pastor, M.; Sousa, F.F.O. Design of Ellagic Acid-Loaded Chitosan/Zein Films for Wound Bandaging. J. Drug Deliv. Sci. Technol. 2020, 59, 101903. [Google Scholar] [CrossRef]
- Tavares, W.d.S.; Pena, G.R.; Martin-Pastor, M.; Sousa, F.F.O. de Design and Characterization of Ellagic Acid-Loaded Zein Nanoparticles and Their Effect on the Antioxidant and Antibacterial Activities. J. Mol. Liq. 2021, 341, 116915. [Google Scholar] [CrossRef]
- Yu, C.; Naeem, A.; Liu, Y.; Guan, Y. Ellagic Acid Inclusion Complex-Loaded Hydrogels as an Efficient Controlled Release System: Design, Fabrication and In Vitro Evaluation. J. Funct. Biomater. 2023, 14, 278. [Google Scholar] [CrossRef]
- Macêdo, N.S.; Barbosa, C.R.d.S.; Bezerra, A.H.; Silveira, Z.d.S.; da Silva, L.; Coutinho, H.D.M.; Dashti, S.; Kim, B.; da Cunha, F.A.B.; da Silva, M.V. Evaluation of Ellagic Acid and Gallic Acid as Efflux Pump Inhibitors in Strains of Staphylococcus aureus. Biol. Open 2022, 11. [Google Scholar] [CrossRef]
- Vigbedor, B.Y.; Akoto, C.O.; Neglo, D. Isolation and Characterization of 3,3′-Di-O-Methyl Ellagic Acid from the Root Bark of Afzelia Africana and Its Antimicrobial and Antioxidant Activities. Sci. Afr. 2022, 17, e01332. [Google Scholar] [CrossRef]
- Dahash, S.L.; Al-Windy, S.B.; Al-Kuraishi, A.H.; Hussien, N.R.; Al-Niemi, M.S.; Al-Kuraishy, H.M. Ellagic Acid-Rich Pomegranate Extracts Synergizes Moxifloxacin against Methicillin Resistance Staphylococcus aureus (MRSA). J. Pak. Med. Assoc. 2021, 71 (Suppl. S8), S88–S92. [Google Scholar]
- Al-Mugdadi, S.F.H.; Al-Sudani, B.; Mohsin, R.A.; Mjali, A.J. Antimicrobial Activity Effects of the Ellagic Acid Extract. Int. J. Res. Pharm. Sci. 2019, 10, 1172–1180. [Google Scholar] [CrossRef]
- Fontaine, B.M.; Nelson, K.; Lyles, J.T.; Jariwala, P.B.; García-Rodriguez, J.M.; Quave, C.L.; Weinert, E.E. Identification of Ellagic Acid Rhamnoside as a Bioactive Component of a Complex Botanical Extract with Anti-Biofilm Activity. Front. Microbiol. 2017, 8, 496. [Google Scholar] [CrossRef] [PubMed]
- Ekrikaya, S.; Yilmaz, E.; Celik, C.; Demirbuga, S.; Ildiz, N.; Demirbas, A.; Ocsoy, I. Investigation of Ellagic Acid Rich-Berry Extracts Directed Silver Nanoparticles Synthesis and Their Antimicrobial Properties with Potential Mechanisms towards Enterococcus Faecalis and Candida Albicans. J. Biotechnol. 2021, 341, 155–162. [Google Scholar] [CrossRef]
- Allemailem, K.S. Enhanced Activity of Ellagic Acid in Lipid Nanoparticles (EA-Liposomes) against Acinetobacter Baumannii in Immunosuppressed Mice. Saudi J. Biol. Sci. 2023, 30, 103707. [Google Scholar] [CrossRef]
- Bai, S.; Yu, Y.; An, L.; Wang, W.; Fu, X.; Chen, J.; Ma, J. Ellagic Acid Increases Stress Resistance via Insulin/IGF-1 Signaling Pathway in Caenorhabditis Elegans. Molecules 2022, 27, 6168. [Google Scholar] [CrossRef]
- Siddiqui, M.F.; Oh, H.-S.; Rzechowicz, M.; Winters, H.; Chong, T.H.; Fane, A.G. Biofouling Control Potential of Tannic Acid, Ellagic Acid, and Epigallocatechin against Pseudomonas Aeruginosa and Reverse Osmosis Membrane Multispecies Community. J. Ind. Eng. Chem. 2015, 30, 204–211. [Google Scholar] [CrossRef]
- Sarabhai, S.; Sharma, P.; Capalash, N. Ellagic Acid Derivatives from Terminalia Chebula Retz. Downregulate the Expression of Quorum Sensing Genes to Attenuate Pseudomonas Aeruginosa PAO1 Virulence. PLoS ONE 2013, 8, e53441. [Google Scholar] [CrossRef]
- Sarabhai, S.; Harjai, K.; Sharma, P.; Capalash, N. Ellagic Acid Derivatives from Terminalia chebula Retz. Increase the Susceptibility of Pseudomonas aeruginosa to Stress by Inhibiting Polyphosphate Kinase. J. Appl. Microbiol. 2015, 118, 817–825. [Google Scholar] [CrossRef]
- El-sonbaty, S.M.; Moawed, F.S.; Kandil, E.I.; Tamamm, A.M. Antitumor and Antibacterial Efficacy of Gallium Nanoparticles Coated by Ellagic Acid. Dose-Response 2022, 20, 155932582110689. [Google Scholar] [CrossRef] [PubMed]
- Jenic, D.; Waller, H.; Collins, H.; Erridge, C. Reversal of Tetracycline Resistance by Cepharanthine, Cinchonidine, Ellagic Acid and Propyl Gallate in a Multi-Drug Resistant Escherichia coli. Nat. Prod. Bioprospect. 2021, 11, 345–355. [Google Scholar] [CrossRef] [PubMed]
- Ratti, A.; Fassi, E.M.A.; Forlani, F.; Mori, M.; Villa, F.; Cappitelli, F.; Sgrignani, J.; Roda, G.; Cavalli, A.; Villa, S.; et al. Mechanistic Insights into the Antibiofilm Mode of Action of Ellagic Acid. Pharmaceutics 2023, 15, 1757. [Google Scholar] [CrossRef] [PubMed]
- Rauf, A.; Imran, M.; Abu-Izneid, T.; Iahtisham-Ul-Haq; Patel, S.; Pan, X.; Naz, S.; Sanches Silva, A.; Saeed, F.; Rasul Suleria, H.A. Proanthocyanidins: A Comprehensive Review. Biomed. Pharmacother. 2019, 116, 108999. [Google Scholar] [CrossRef] [PubMed]
- Dixon, R.A.; Sarnala, S. Proanthocyanidin Biosynthesis—A Matter of Protection. Plant Physiol. 2020, 184, 579–591. [Google Scholar] [CrossRef]
- Qi, Q.; Chu, M.; Yu, X.; Xie, Y.; Li, Y.; Du, Y.; Liu, X.; Zhang, Z.; Shi, J.; Yan, N. Anthocyanins and Proanthocyanidins: Chemical Structures, Food Sources, Bioactivities, and Product Development. Food Rev. Int. 2023, 39, 4581–4609. [Google Scholar] [CrossRef]
- Yu, K.; Song, Y.; Lin, J.; Dixon, R.A. The Complexities of Proanthocyanidin Biosynthesis and Its Regulation in Plants. Plant Commun. 2023, 4, 100498. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Liang, Y.; Ren, L.; Zhang, T. In Vitro Anti-Inflammatory Potency of Sanguinarine and Chelerythrine via Interaction with Glucocorticoid Receptor. eFood 2020, 1, 392–398. [Google Scholar] [CrossRef]
- Zhang, Y.; Dong, Y.; Li, X.; Wang, F. Proanthocyanidin Encapsulated in Ferritin Enhances Its Cellular Absorption and Antioxidant Activity. J. Agric. Food Chem. 2019, 67, 11498–11507. [Google Scholar] [CrossRef]
- Li, X.; Liu, J.; Chang, Q.; Zhou, Z.; Han, R.; Liang, Z. Antioxidant and Antidiabetic Activity of Proanthocyanidins from Fagopyrum Dibotrys. Molecules 2021, 26, 2417. [Google Scholar] [CrossRef]
- Chen, J.; Chen, Y.; Zheng, Y.; Zhao, J.; Yu, H.; Zhu, J.; Li, D. Neuroprotective Effects and Mechanisms of Procyanidins In Vitro and In Vivo. Molecules 2021, 26, 2963. [Google Scholar] [CrossRef] [PubMed]
- Ruan, Y.; Jin, Q.; Zeng, J.; Ren, F.; Xie, Z.; Ji, K.; Wu, L.; Wu, J.; Li, L. Grape Seed Proanthocyanidin Extract Ameliorates Cardiac Remodelling after Myocardial Infarction through PI3K/AKT Pathway in Mice. Front. Pharmacol. 2020, 11, 585984. [Google Scholar] [CrossRef] [PubMed]
- Alfaro-Viquez, E.; Esquivel-Alvarado, D.; Madrigal-Carballo, S.; Krueger, C.G.; Reed, J.D. Antimicrobial Proanthocyanidin-Chitosan Composite Nanoparticles Loaded with Gentamicin. Int. J. Biol. Macromol. 2020, 162, 1500–1508. [Google Scholar] [CrossRef] [PubMed]
- Andersen-Civil, A.I.S.; Arora, P.; Williams, A.R. Regulation of Enteric Infection and Immunity by Dietary Proanthocyanidins. Front. Immunol. 2021, 12, 637603. [Google Scholar] [CrossRef]
- Nawrot-Hadzik, I.; Matkowski, A.; Kubasiewicz-Ross, P.; Hadzik, J. Proanthocyanidins and Flavan-3-Ols in the Prevention and Treatment of Periodontitis—Immunomodulatory Effects, Animal and Clinical Studies. Nutrients 2021, 13, 239. [Google Scholar] [CrossRef]
- Liu, M.; Yun, P.; Hu, Y.; Yang, J.; Khadka, R.B.; Peng, X. Effects of Grape Seed Proanthocyanidin Extract on Obesity. Obes. Facts 2020, 13, 279–291. [Google Scholar] [CrossRef]
- Lu, Z.; Jia, Q.; Wang, R.; Wu, X.; Wu, Y.; Huang, C.; Li, Y. Hypoglycemic Activities of A- and B-Type Procyanidin Oligomer-Rich Extracts from Different Cinnamon Barks. Phytomedicine 2011, 18, 298–302. [Google Scholar] [CrossRef]
- Bladé, C.; Arola, L.; Salvadó, M.J. Hypolipidemic Effects of Proanthocyanidins and Their Underlying Biochemical and Molecular Mechanisms. Mol. Nutr. Food Res. 2010, 54, 37–59. [Google Scholar] [CrossRef]
- Kawahara, S.I.; Ishihara, C.; Matsumoto, K.; Senga, S.; Kawaguchi, K.; Yamamoto, A.; Suwannachot, J.; Hamauzu, Y.; Makabe, H.; Fujii, H. Identification and Characterization of Oligomeric Proanthocyanidins with Significant Anti-Cancer Activity in Adzuki Beans (Vigna Angularis). Heliyon 2019, 5, e02610. [Google Scholar] [CrossRef]
- Kumar, N.S.; Bandara, B.R.; Hettihewa, S.K.; Panagoda, G.J. Oligomeric Proanthocyanidin Fractions from Fresh Tea Leaves and Their Antibacterial Activity against Staphylococcus aureus. J. Natl. Sci. Found. Sri Lanka 2014, 42, 241. [Google Scholar] [CrossRef]
- Nakamura, K.; Shirato, M.; Ikai, H.; Kanno, T.; Sasaki, K.; Kohno, M.; Niwano, Y. Photo-Irradiation of Proanthocyanidin as a New Disinfection Technique via Reactive Oxygen Species Formation. PLoS ONE 2013, 8, e60053. [Google Scholar] [CrossRef] [PubMed]
- Lai, H.Y.; Lim, Y.Y.; Kim, K.H. Isolation and Characterisation of a Proanthocyanidin with Antioxidative, Antibacterial and Anti-Cancer Properties from Fern Blechnum Orientale. Pharmacogn. Mag. 2017, 13, 31–37. [Google Scholar] [PubMed]
- Shejawal, K.P.; Randive, D.S.; Bhinge, S.D.; Bhutkar, M.A.; Wadkar, G.H.; Jadhav, N.R. Green Synthesis of Silver and Iron Nanoparticles of Isolated Proanthocyanidin: Its Characterization, Antioxidant, Antimicrobial, and Cytotoxic Activities against COLO320DM and HT29. J. Genet. Eng. Biotechnol. 2020, 18, 43. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.Y.; Liu, Y.; Mao, J.; Wu, Y.B.; Deng, Y.L.; Qi, S.C.; Zhou, Y.C.; Gong, S.Q. The Antibiofilm and Collagen-stabilizing Effects of Proanthocyanidin as an Auxiliary Endodontic Irrigant. Int. Endod. J. 2020, 53, 824–833. [Google Scholar] [CrossRef] [PubMed]
- Alfaro-Viquez, E.; Esquivel-Alvarado, D.; Madrigal-Carballo, S.; Krueger, C.G.; Reed, J.D. Proanthocyanidin-Chitosan Composite Nanoparticles Prevent Bacterial Invasion and Colonization of Gut Epithelial Cells by Extra-Intestinal Pathogenic Escherichia coli. Int. J. Biol. Macromol. 2019, 135, 630–636. [Google Scholar] [CrossRef] [PubMed]
- Alfaro-Viquez, E.; Esquivel-Alvarado, D.; Madrigal-Carballo, S.; Krueger, C.G.; Reed, J.D. Cranberry Proanthocyanidin-Chitosan Hybrid Nanoparticles as a Potential Inhibitor of Extra-Intestinal Pathogenic Escherichia coli Invasion of Gut Epithelial Cells. Int. J. Biol. Macromol. 2018, 111, 415–420. [Google Scholar] [CrossRef] [PubMed]
- Feliciano, R.P.; Meudt, J.J.; Shanmuganayagam, D.; Krueger, C.G.; Reed, J.D. Ratio of “a-Type” to “b-Type” Proanthocyanidin Interflavan Bonds Affects Extra-Intestinal Pathogenic Escherichia coli Invasion of Gut Epithelial Cells. J. Agric. Food Chem. 2014, 62, 3919–3925. [Google Scholar] [CrossRef]
- Vadekeetil, A.; Alexandar, V.; Chhibber, S.; Harjai, K. Adjuvant Effect of Cranberry Proanthocyanidin Active Fraction on Antivirulent Property of Ciprofloxacin against Pseudomonas Aeruginosa. Microb. Pathog. 2016, 90, 98–103. [Google Scholar] [CrossRef]
- Maisuria, V.B.; Okshevsky, M.; Déziel, E.; Tufenkji, N. Proanthocyanidin Interferes with Intrinsic Antibiotic Resistance Mechanisms of Gram-Negative Bacteria. Adv. Sci. 2019, 6. [Google Scholar] [CrossRef]
- Esquivel-Alvarado, D.; Alfaro-Viquez, E.; Polewski, M.A.; Krueger, C.G.; Vestling, M.M.; Reed, J.D. Synthesis of Fluorescent Proanthocyanidin-Cinnamaldehydes Pyrylium Products for Microscopic Detection of Interactions with Extra-Intestinal PathogenicEscherichia coli. J. Agric. Food Chem. 2021, 69, 10700–10708. [Google Scholar] [CrossRef]
- Mackraj, I.; Govender, T.; Gathiram, P. Sanguinarine. Cardiovasc. Ther. 2008, 26, 75–83. [Google Scholar] [CrossRef]
- Singh, N.; Sharma, B. Toxicological Effects of Berberine and Sanguinarine. Front. Mol. Biosci. 2018, 5, 21. [Google Scholar] [CrossRef]
- Miao, F.; Yang, X.J.; Zhou, L.; Hu, H.J.; Zheng, F.; Ding, X.D.; Sun, D.M.; Zhou, C.D.; Sun, W. Structural Modification of Sanguinarine and Chelerythrine and Their Antibacterial Activity. Nat. Prod. Res. 2011, 25, 863–875. [Google Scholar] [CrossRef] [PubMed]
- Bavarsadi, M.; Mahdavi, A.H.; Ansari-Mahyari, S.; Jahanian, E. Effects of Different Levels of Sanguinarine on Antioxidant Indices, Immunological Responses, Ileal Microbial Counts and Jejunal Morphology of Laying Hens Fed Diets with Different Levels of Crude Protein. J. Anim. Physiol. Anim. Nutr. (Berl). 2017, 101, 936–948. [Google Scholar] [CrossRef] [PubMed]
- Niu, X.; Fan, T.; Li, W.; Xing, W.; Huang, H. The Anti-Inflammatory Effects of Sanguinarine and Its Modulation of Inflammatory Mediators from Peritoneal Macrophages. Eur. J. Pharmacol. 2012, 689, 262–269. [Google Scholar] [CrossRef]
- Serafim, T.L.; Matos, J.A.C.; Sardão, V.A.; Pereira, G.C.; Branco, A.F.; Pereira, S.L.; Parke, D.; Perkins, E.L.; Moreno, A.J.M.; Holy, J.; et al. Sanguinarine Cytotoxicity on Mouse Melanoma K1735-M2 Cells-Nuclear vs. Mitochondrial Effects. Biochem. Pharmacol. 2008, 76, 1459–1475. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Deng, L.; Tang, Y.; Li, J.; Zhong, T.; Hao, X.; Fan, Y.; Mu, S. Cytostatic Activity of Sanguinarine and a Cyanide Derivative in Human Erythroleukemia Cells Is Mediated by Suppression of C-MET/MAPK Signaling. Int. J. Mol. Sci. 2023, 24, 8113. [Google Scholar] [CrossRef] [PubMed]
- Chien, M.H.; Yu, W.C.; Jiunn, W.L.; Huei, W.C.; Jaw, J.K. Induction of Contracture and Extracellular Ca2+ Influx in Cardiac Muscle by Sanguinarine: A Study on Cardiotoxicity of Sanguinarine. J. Biomed. Sci. 2005, 12, 399–407. [Google Scholar] [CrossRef]
- Jeng, J.H.; Wu, H.L.; Lin, B.R.; Lan, W.H.; Chang, H.H.; Ho, Y.S.; Lee, P.H.; Wang, Y.J.; Wang, J.S.; Chen, Y.J.; et al. Antiplatelet Effect of Sanguinarine Is Correlated to Calcium Mobilization, Thromboxane and CAMP Production. Atherosclerosis 2007, 191, 250–258. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Jiang, M.; Liu, Q.; Yan, S.; Feng, L.; Lan, Y.; Shan, G.; Xue, W.; Guo, R. Enhanced Healing Activity of Burn Wound Infection by a Dextran-HA Hydrogel Enriched with Sanguinarine. Biomater. Sci. 2018, 6, 2472–2486. [Google Scholar] [CrossRef]
- Gu, Y.; Dong, J.; Li, J.; Luo, Q.; Dong, X.; Tang, G.; Zhang, J.; Du, X.; Pu, Q.; He, L.; et al. Antibacterial Activity and Mechanism of Sanguinarine against Staphylococcus aureus by Interfering with the Permeability of the Cell Wall and Membrane and Inducing Bacterial ROS Production. Front. Vet. Sci. 2023, 10, 1121082. [Google Scholar] [CrossRef] [PubMed]
- Hamoud, R.; Reichling, J.; Wink, M. Synergistic Antibacterial Activity of the Combination of the Alkaloid Sanguinarine with EDTA and the Antibiotic Streptomycin against Multidrug Resistant Bacteria. J. Pharm. Pharmacol. 2015, 67, 264–273. [Google Scholar] [CrossRef] [PubMed]
- Osei-Owusu, H.; Kudera, T.; Strakova, M.; Rondevaldova, J.; Skrivanova, E.; Novy, P.; Kokoska, L. In Vitro Selective Combinatory Effect of Ciprofloxacin with Nitroxoline, Sanguinarine, and Zinc Pyrithione against Diarrhea-Causing and Gut Beneficial Bacteria. Microbiol. Spectr. 2022, 10. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wu, X.; Wang, Q.; Xu, W.; Zhao, Q.; Xu, N.; Hu, X.; Ye, Z.; Yu, S.; Liu, J.; et al. Sanguinarine Ameliorates DSS Induced Ulcerative Colitis by Inhibiting NLRP3 Inflammasome Activation and Modulating Intestinal Microbiota in C57BL/6 Mice. Phytomedicine 2022, 104, 154321. [Google Scholar] [CrossRef]
- Liang, J.-Y.; Li, Q.; Feng, L.-B.; Hu, S.-X.; Zhang, S.-Q.; Li, C.-X.; Zhang, X.-B. Injectable Antimicrobial Hydrogels with Antimicrobial Peptide and Sanguinarine Controlled Release Ability for Preventing Bacterial Infections. Am. J. Transl. Res. 2021, 13, 12614–12625. [Google Scholar]
- Lu, C.; Zhang, N.; Kou, S.; Gao, L.; Peng, B.; Dai, Y.; Zheng, J. Sanguinarine Synergistically Potentiates Aminoglycoside-Mediated Bacterial Killing. Microb. Biotechnol. 2022, 15, 2055–2070. [Google Scholar] [CrossRef]
- Falchi, F.A.; Borlotti, G.; Ferretti, F.; Pellegrino, G.; Raneri, M.; Schiavoni, M.; Caselli, A.; Briani, F. Sanguinarine Inhibits the 2-Ketogluconate Pathway of Glucose Utilization in Pseudomonas Aeruginosa. Front. Microbiol. 2021, 12, 744458. [Google Scholar] [CrossRef]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
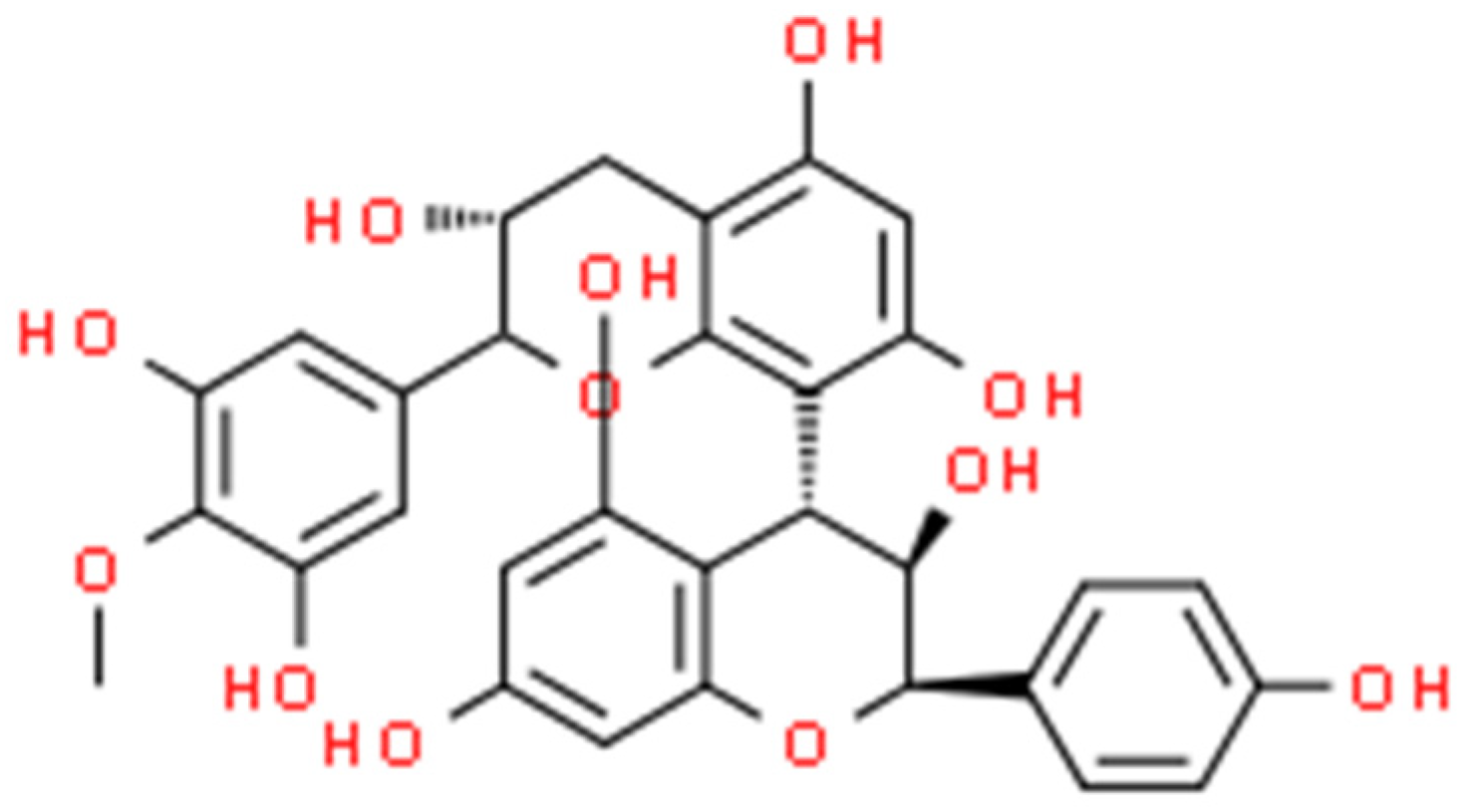{kind=link}
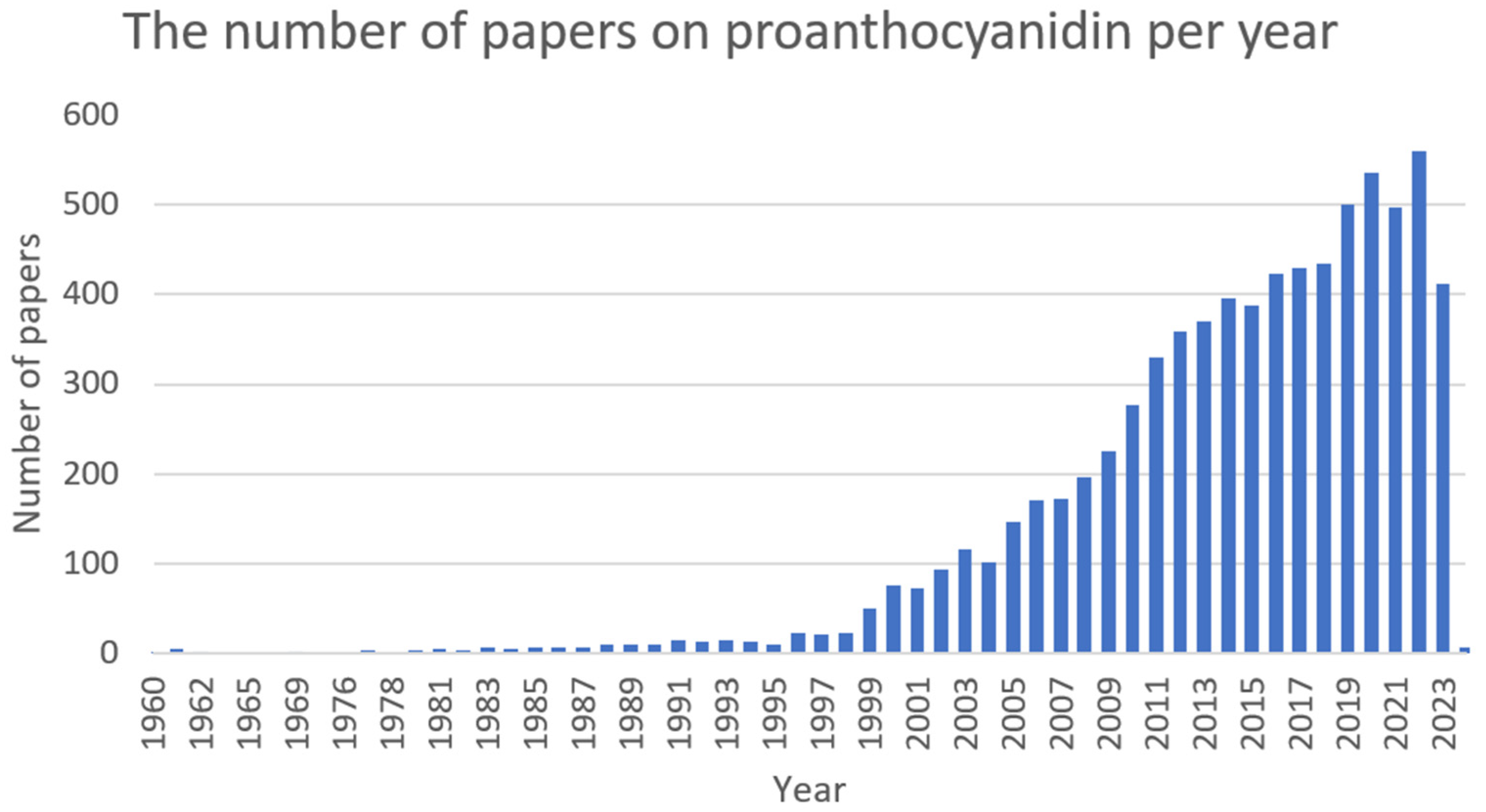{kind=link}
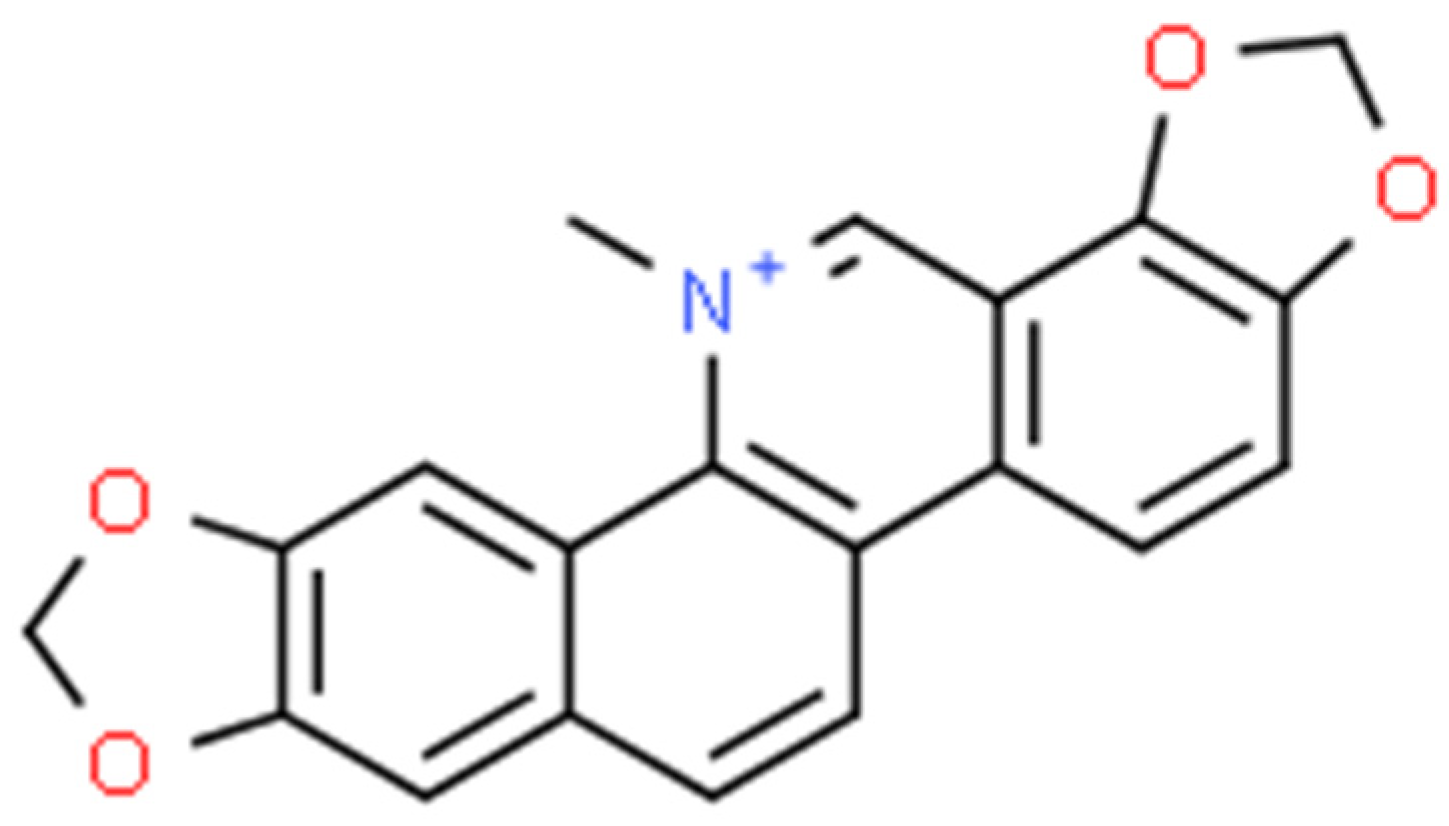{kind=link}
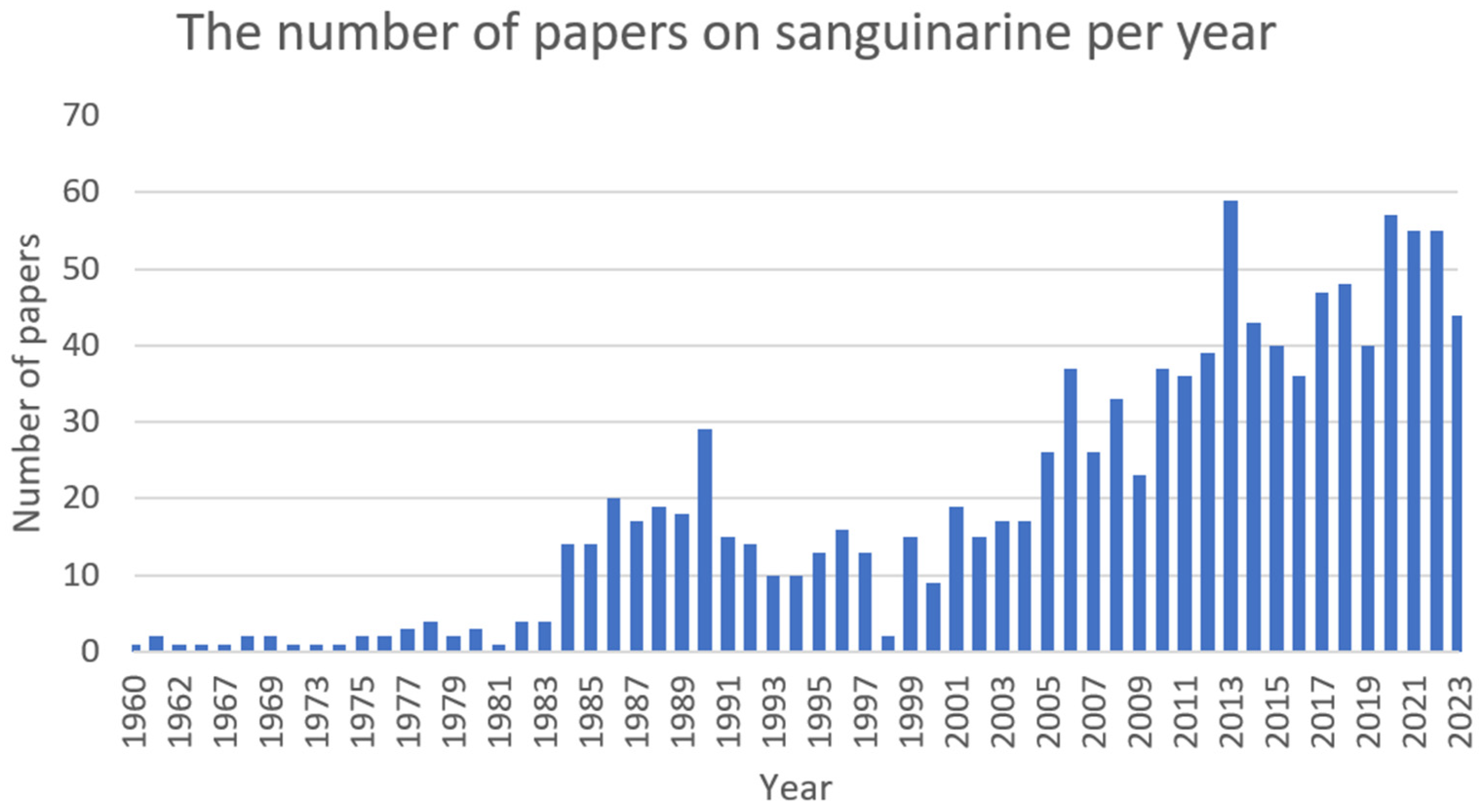{kind=link}
| Substance | Antibacterial Effect | References |
|---|---|---|
| Berberine | Inhibition of D-Ala-D-Ala synthesis Inhibition of metabolism of nucleic acids and aminosaccharides Generation of oxidative stress with inhibition of antioxidant synthesis | [99] |
| FtsZ protein inhibition | [75] | |
| acrAB-TolC efflux pump overexpression | [82] | |
| Nucleic acid metabolism dysfunction Cellular wall lysis Inhibition of transmembrane transport | [101] | |
| Inhibition of bacterial DNA synthesis | [89] | |
| Inhibition of the MexXY-OprM efflux pump | [91,94,95,96,103] | |
| Catechin | Increase in oxidative stress due to the generation of ROS in bacterial cells Degradation of DNA polymers | [140] |
| Inhibition of norA, norC, and abcA MRSA efflux pump gene expression | [123] | |
| Direct inhibition of MurA, GBAP, and gelatinase | [127] | |
| Chelerythrine | Increase in membrane permeability | [148] |
| Cinnamaldehyde | Inhibition of phosphatidylglycerol and phosphatidylethanolamine metabolism Increase in cell membrane permeability | [209] |
| Inhibition of mecA and hld gene expression | [172] | |
| Inhibition of (eno), (ebps), and (fib) gene expression | [210] | |
| Inhibition of acrB gene expression Inhibition of blaTEM, blaSHV, and blaCTX-M genes expression | [184] | |
| Inhibition of the FtsZ protein | [186,211] | |
| Inhibition of: blaP, adeA, adeB, adeC, mdrp gene expression | [187] | |
| Inhibition of Las and Rhl virulence factors | [189] | |
| Inhibition of exoS and ampC gene expression | [193] | |
| Inhibition of flgD, flhB, fliP, and fliR gene expression | [58] | |
| Ellagic acid | Direct inhibition of polyphosphate kinase 1 Inhibition of rpoS gene expression Decrease in catalase and peroxide dismutase activity Cell damage due to ROS accumulation | [248] |
| Cell damage due to ROS accumulation | [251] | |
| Proanthocyanidin | Bacterial agglutination due to affinity for the fimbriae | [278] |
| Sanguinarine | Cell damage due to ROS generation Increase of membrane permeability | [289] |
| Direct inhibition of KguD or KguK enzymes Inhibition of cellular glucose–6-phosphate synthesis | [295] |
| Substance | Antibiofilm Effect | References |
|---|---|---|
| Berberine | Inhibition of agrA-D gene expression | [68] |
| Inhibition of PSM aggregation | [74] | |
| Inhibition of expression of sortase genes (srtA and esp) | [80] | |
| Decrease in the production of alginate | [93] | |
| Inhibition of sarA, cidA, and icaA gene expression | [100] | |
| Inhibition of luxS, pfS, hflX, ftsQ, uxS, and ftsE gene expression | [102] | |
| Inhibition of the exopolysaccharide synthesis | [90] | |
| Inhibition of QS signalling by docking at LasR and RhlR | [92,105] | |
| Catechin | Inhibition of the GBAP protein | [127] |
| Direct inhibition of the BfmR protein | [135] | |
| Inhibition of lasR and lasI gene expression | [137] | |
| Direct inhibition of the LasR factor | [137] | |
| Cinnamaldehyde | Increased expression of the AI-2 factor | [168] |
| Reduction of bacterial exopolysaccharide formation | [179] | |
| Direct inhibition of ESP protein | [180] | |
| Decreased level of QS involved genes encoding: hemolysins, proteases, elastases, alginate, pyrocyanin, and ramnolipid expression | [190,191,192] | |
| Inhibition of exopolysaccharide synthesis | [190] | |
| Inhibition of icaA and icaD gene expression | [173] | |
| Inhibition of gene-encoding PIA glucosamine polymer expression | [173] | |
| Inhibition of fsrB, fsrC, luxS, and gelE gene expression | [179] | |
| Inhibition of: csuA, csuE, bfmS, bfmR, ompa, bap, abal genes expression | [188] | |
| Reduction of the intracellular concentration of bis-(3′-5′)-cyclic dimeric GMP | [213] | |
| Reduction in the formation of acyl homoserine lactones | [191] | |
| Inhibition of luxS, fimA, and fliC gene expression | [202] | |
| Ellagic acid | Inhibition of lasIR and rhlIR gene expression | [247] |
| Inhibition of the formation of exopolysaccharides | [246] | |
| Direct inhibition of WrbA protein | [251] | |
| Inhibition of the synthesis of alginate | [248] | |
| Proanthocyanidine | Reduction of expression of genes that encode: elastase, cell-associated hemolysin, cell-free hemolysin, rhamnolipid, and alginate | [276] |
| Inhibition of flagellin synthesis due to inhibition of fliC gene expression | [275] | |
| Inhibition of the production of acyl-hoserine lactone | [276] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pacyga, K.; Pacyga, P.; Topola, E.; Viscardi, S.; Duda-Madej, A. Bioactive Compounds from Plant Origin as Natural Antimicrobial Agents for the Treatment of Wound Infections. Int. J. Mol. Sci. 2024, 25, 2100. https://doi.org/10.3390/ijms25042100
Pacyga K, Pacyga P, Topola E, Viscardi S, Duda-Madej A. Bioactive Compounds from Plant Origin as Natural Antimicrobial Agents for the Treatment of Wound Infections. International Journal of Molecular Sciences. 2024; 25(4):2100. https://doi.org/10.3390/ijms25042100
Chicago/Turabian StylePacyga, Katarzyna, Paweł Pacyga, Ewa Topola, Szymon Viscardi, and Anna Duda-Madej. 2024. "Bioactive Compounds from Plant Origin as Natural Antimicrobial Agents for the Treatment of Wound Infections" International Journal of Molecular Sciences 25, no. 4: 2100. https://doi.org/10.3390/ijms25042100