
Fuziline Ameliorates Glucose and Lipid Metabolism by Activating Beta Adrenergic Receptors to Stimulate Thermogenesis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
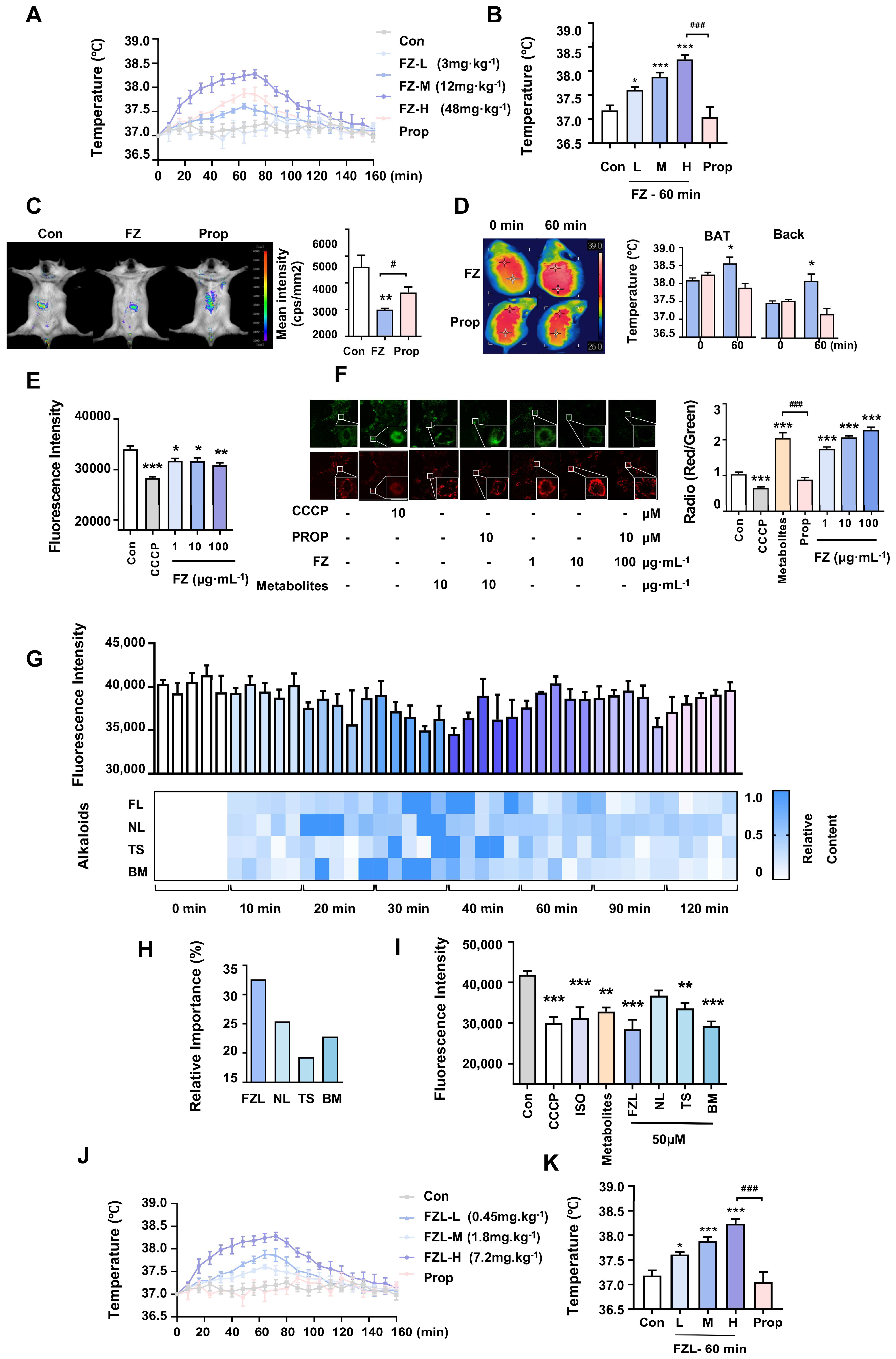
2.1. Radix Aconiti Carmichaeli Stimulated Thermogenesis via Activation of β-ARs
2.2. FZL Is the Key Bioactive Compound in FZ That Promotes Thermogenesis
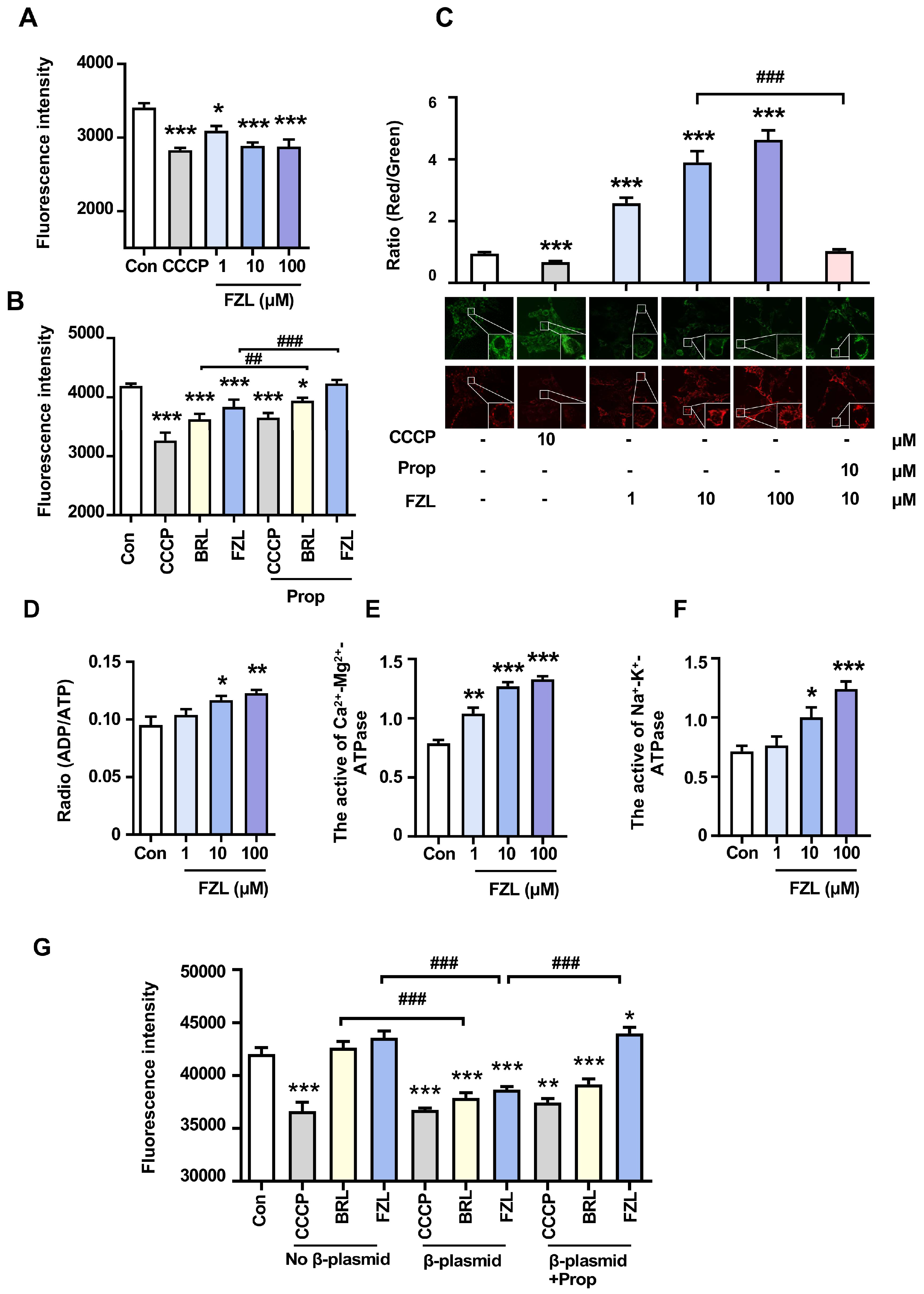
2.3. FZL Stimulated Thermogenesis through Activation of β3-ARs
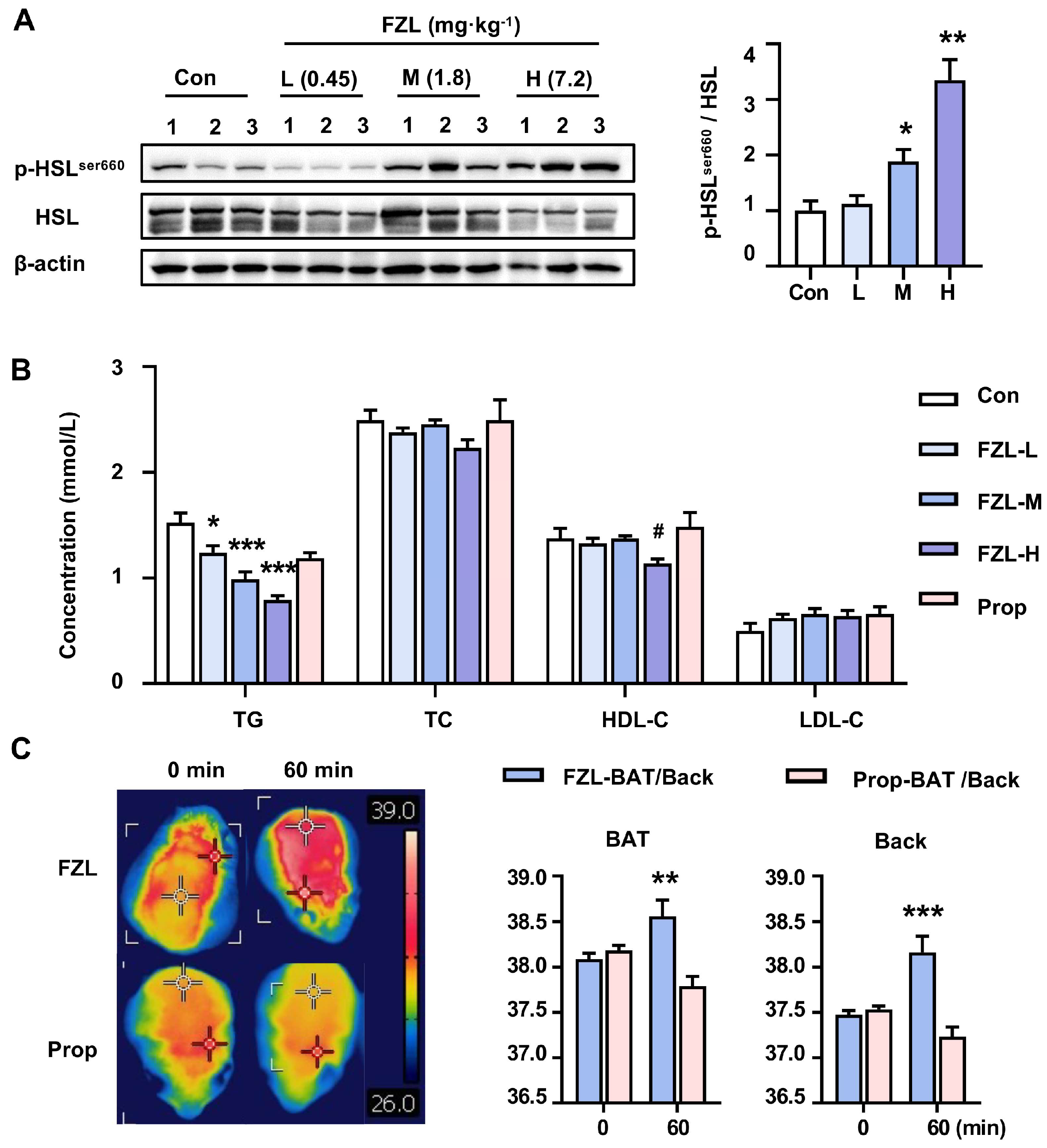
2.4. FZL Stimulated BAT Thermogenesis via β3-AR-Mediated Lipolysis
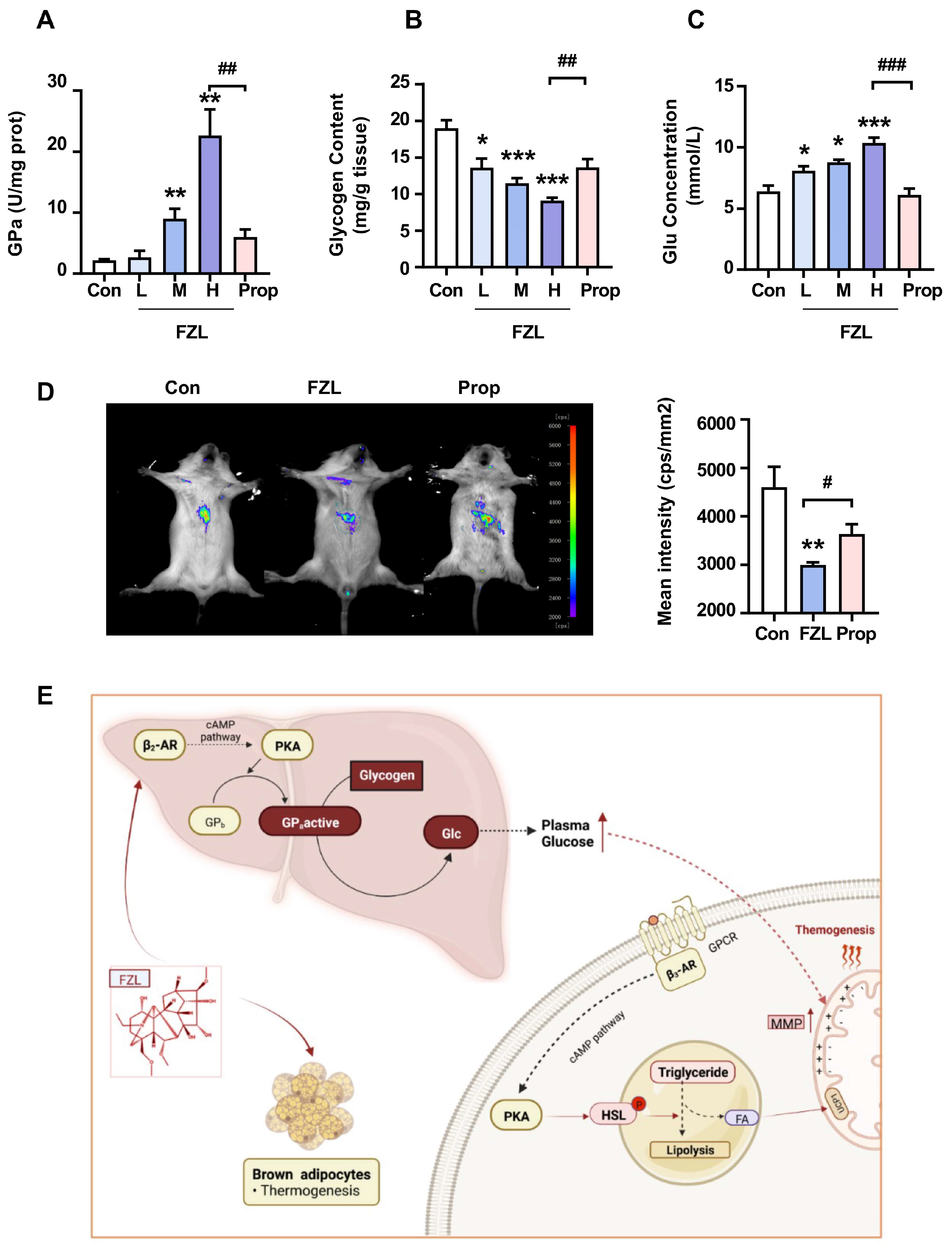
2.5. FZL Accelerated Liver Glycogenolysis through Activation of β2-Adrenergic Receptor (β2-AR)
3. Materials and Methods
3.1. Chemicals and Reagents
3.2. Extracts of Radix Aconiti Carmichaeli Preparation
3.3. UPLC-Q/TOF-MS Analysis of FZ
3.4. Animals
3.5. Anal Temperature Measurement in Rats
3.6. Liver Temperature Measurement
3.7. Thermographic Imaging
3.8. Cell Culture
3.9. Mitochondrial Membrane Electrochemical Potential (MMP) and Temperature Assay
3.10. Metabolites Identification
3.11. SPSS Analysis
3.12. ADP/ATP Ratio and ATPase Enzymatic Activity Assay in BFCs
3.13. Detection of Intracellular and Circulating Lipolytic Activity
3.14. Detection of Liver Glycogenolysis Activity
3.15. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yu, H.Y.; Wang, S.J.; Teng, J.L.; Ji, X.M.; Wu, Z.C.; Ma, Q.C.; Fu, X.J. Effects of Radix aconiti lateralis preparata and Rhizoma zingiberis on energy metabolism and expression of the genes related to metabolism in rats. Chin. J. Integr Med. 2012, 18, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Makino, T.; Kato, K.; Mizukami, H. Processed aconite root prevents cold-stress-induced hypothermia and immuno-suppression in mice. Biol. Pharm. Bull. 2009, 32, 741–1748. [Google Scholar] [CrossRef] [PubMed]
- Huh, E.; Lee, W.; Choi, Y.; Lee, T.H.; Oh, M.S. Geongangbuja-Tang Decoction and Its Active Ingredient, Aconiti Lateralis Radix Preparata, Exerts Inhibitory Effects on Heat Stress-Induced Inflammation in Mice. Appl. Sci. 2021, 11, 6902. [Google Scholar] [CrossRef]
- Liu, J.; Tan, Y.; Ao, H.; Feng, W.; Peng, C. Aqueous extracts of Aconite promote thermogenesis in rats with hypothermia via regulating gut microbiota and bile acid metabolism. Chin. Med. 2021, 16, 29. [Google Scholar] [CrossRef]
- Zhou, B.C.; Li, T.Y.N.; Yang, M.N.; Pang, J.X.; Min, L.Y.; Han, J.X. Characterization of the hot and cold medicinal properties of traditional Chinese herbs by spontaneous photon emission ratio of mice. J. Ethnopharmacol. 2019, 243, 112108. [Google Scholar] [CrossRef]
- Zhou, N.; Yang, Y.; Li, K.; Ke, Y.; Zheng, X.; Feng, W.; Bai, Z.; Liu, T.; Wang, Y.; Liu, Z.; et al. Integrating strategies of chemistry, biochemistry and metabolomics for characterization of the medication principle of treating cold/heat syndrome with hot/cold herbs. J. Ethnopharmacol. 2019, 239, 111899. [Google Scholar] [CrossRef]
- Liu, J.; Feng, W.; Peng, C. A Song of Ice and Fire: Cold and Hot Properties of Traditional Chinese Medicines. Front. Pharmacol. 2021, 11, 598744. [Google Scholar] [CrossRef]
- Wang, Q.H.; Yang, X.; Li, X.L.; Yang, B.Y.; Wang, Y.H.; Kuang, X.H. Based on proteomics to study the property and flavor of radix aconiti lateralis preparata, rhizoma zingiberis, pricklyash peel. World. J. Tradit. Chin. Med. 2015, 10, 1824–1836. [Google Scholar]
- Zhao, Y.; Wang, J.; Sun, X.; Jia, L.; Li, J.; Shan, L.; Li, R.; Liu, H.; Wang, R.; Song, X.; et al. Microcalorimetry coupled with chemometric techniques for toxicity evaluation of Radix Aconiti Lateralis Preparata (Fuzi) and its processed products on Escherichia coli. Appl. Microbiol. Biotechnol. 2014, 98, 437–444. [Google Scholar] [CrossRef]
- Zhou, G.; Tang, L.; Zhou, X.; Wang, T.; Kou, Z.; Wang, Z. A review on phytochemistry and pharmacological activities of the processed lateral root of Aconitum carmichaelii Debeaux. J. Ethnopharmacol. 2015, 160, 173–193. [Google Scholar] [CrossRef]
- Muller, M.J.; Bosy-Westphal, A. Adaptive thermogenesis with weight loss in humans. Obesity 2013, 21, 218–228. [Google Scholar] [CrossRef]
- Jansky, L. Non-shivering thermogenesis and its thermoregulatory significance. Biol. Rev. Camb. Philos. Soc. 1973, 48, 85–132. [Google Scholar] [CrossRef] [PubMed]
- Dewasmes, G.; Loos, N.; Delanaud, S.; Ramadan, W.; Dewasmes, D. Liver temperature during sleep. Sleep 2003, 26, 948–950. [Google Scholar] [CrossRef]
- Tan, C.Y.; Ishikawa, K.; Virtue, S.; Vidal-Puig, A. Brown adipose tissue in the treatment of obesity and diabetes: Are we hot enough? J. Diabetes Investig. 2011, 2, 341–350. [Google Scholar] [CrossRef] [PubMed]
- Lowell, B.B.; Spiegelman, B.M. Towards a molecular understanding of adaptive thermogenesis. Nature 2000, 404, 652–660. [Google Scholar] [CrossRef] [PubMed]
- Periasamy, M.; Maurya, S.K.; Sahoo, S.K.; Singh, S.; Sahoo, S.K.; Reis, F.C.G.; Bal, N.C. Role of SERCA Pump in Muscle Thermogenesis and Metabolism. Compr. Physiol. 2017, 7, 879–890. [Google Scholar] [PubMed]
- Nedergaard, J.; Cannon, B.; Lindberg, O. Microcalorimetry of isolated mammalian cells. Nature 1977, 267, 518–520. [Google Scholar] [CrossRef]
- Leiria, L.O.; Tseng, Y.H. Lipidomics of brown and white adipose tissue: Implications for energy metabolism. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2020, 1865, 158788. [Google Scholar] [CrossRef]
- Scheja, L.; Heeren, J. Metabolic interplay between white, beige, brown adipocytes and the liver. J. Hepatol. 2016, 64, 1176–1186. [Google Scholar] [CrossRef]
- Nonogaki, K. New insights into sympathetic regulation of glucose and fat metabolism. Diabetologia 2000, 43, 533–549. [Google Scholar] [CrossRef]
- Dijk, W.; Heine, M.; Vergnes, L.; Boon, M.R.; Schaart, G.; Hesselink, M.K.; Reue, K.; Van Marken Lichtenbelt, W.D.; Olivecrona, G.; Rensen, P.C.; et al. ANGPTL4 mediates shuttling of lipid fuel to brown adipose tissue during sustained cold exposure. Elife 2015, 4, e08428. [Google Scholar] [CrossRef] [PubMed]
- Mottillo, E.P.; Balasubramanian, P.; Lee, Y.H.; Weng, C.; Kershaw, E.E.; Granneman, J.G. Coupling of lipolysis and de novo lipogenesis in brown, beige, and white adipose tissues during chronic β3-adrenergic receptor activation. J. Lipid Res. 2014, 55, 2276–2286. [Google Scholar] [CrossRef] [PubMed]
- Guilherme, A.; Rowland, L.A.; Wang, H.; Czech, M.P. The adipocyte supersystem of insulin and cAMP signaling. Trends Cell Biol. 2023, 33, 340–354. [Google Scholar] [CrossRef] [PubMed]
- Morrison, S.F.; Madden, C.J.; Tupone, D. Central neural regulation of brown adipose tissue thermogenesis and energy expenditure. Cell Metab. 2014, 19, 741–756. [Google Scholar] [CrossRef] [PubMed]
- McNeill, B.T.; Morton, N.M.; Stimson, R.H. Substrate Utilization by Brown Adipose Tissue: What’s Hot and What’s Not? Front. Endocrinol. 2020, 11, 571659. [Google Scholar] [CrossRef]
- Ikeda, K.; Kang, Q.; Yoneshiro, T.; Camporez, J.P.; Maki, H.; Homma, M.; Shinoda, K.; Chen, Y.; Lu, X.; Maretich, P.; et al. UCP1-independent signaling involving SERCA2b-mediated calcium cycling regulates beige fat thermogenesis and systemic glucose homeostasis. Nat. Med. 2017, 23, 1454–1465. [Google Scholar] [CrossRef]
- Wu, J.J.; Guo, Z.Z.; Zhu, Y.F.; Huang, Z.J.; Gong, X.; Li, Y.H.; Son, W.J.; Li, X.Y.; Lou, Y.M.; Zhu, L.J.; et al. A systematic review of pharmacokinetic studies on herbal drug Fuzi: Implications for Fuzi as personalized medicine. Phytomedicine 2018, 44, 187–203. [Google Scholar] [CrossRef]
- Li, S.; Yu, L.; Shi, Q.; Liu, Y.; Zhang, Y.; Wang, S.; Lai, X. An insight into current advances on pharmacology, pharmacokinetics, toxicity and detoxification of aconitine. Biomed. Pharmacother. 2022, 151, 113115. [Google Scholar] [CrossRef]
- Chang, N.; Gao, J.; Niu, L.; Hou, Y.; Wang, X.; Jiang, M.; Bai, G. Integrated artificial neural network analysis and functional cell-based affinity mass spectrometry for screening a bifunctional activator of Ca2+ and β2-AR in aconite. J. Pharm. Biomed. Anal. 2020, 190, 113506. [Google Scholar] [CrossRef]
- Ameri, A. Effects of the Aconitum alkaloid mesaconitine in rat hippocampal slices and the involvement of alpha- and beta-adrenoceptors. Br. J. Pharmacol. 1998, 123, 243–250. [Google Scholar] [CrossRef]
- Huang, Y.; Li, L.; Li, X.; Fan, S.; Zhuang, P.; Zhang, Y. Ginseng Compatibility Environment Attenuates Toxicity and Keeps Efficacy in Cor Pulmonale Treated by Fuzi Beimu Incompatibility Through the Coordinated Crosstalk of PKA and Epac Signaling Pathways. Front. Pharmacol. 2018, 9, 634. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Lu, Z.Q.; Zhang, Y.J.; Li, Y.B.; Wang, Z.Y.; Zhang, Y.L.; Zhuang, P.W.; Bai, G. Looking for agonists of β2 adrenergic receptor from Fuzi and Chuanwu by virtual screening and dual-luciferase reporter assay. J. Asian Nat. Prod. Res. 2016, 18, 550–561. [Google Scholar] [CrossRef] [PubMed]
- Feng, G.P.; Zhang, S.D.; Yi, N.Y. Effects of Rehmannia glutinosa, Plastrum testudinis, Aconitum carmichaeli and Cinnamomum cassia on the β-adrenergic receptors of hyperthyroid rat kidneys. Zhong Xi Yi Jie He Za Zhi 1986, 6, 582–606. [Google Scholar] [PubMed]
- Chang, S.H.; Song, N.J.; Choi, J.H.; Yun, U.J.; Park, K.W. Mechanisms underlying UCP1 dependent and independent adipocyte thermogenesis. Obes. Rev. 2019, 20, 241–251. [Google Scholar] [CrossRef]
- Thiebaud, D.; Schutz, Y.; Acheson, K.; Jacot, E.; DeFronzo, R.A.; Felber, J.P.; Jequier, E. Energy cost of glucose storage in human subjects during glucose-insulin infusions. Am. J. Physiol. 1983, 244, E216–E221. [Google Scholar] [CrossRef]
- Bai, G.; Yang, Y.; Shi, Q.; Liu, Z.; Zhang, Q.; Zhu, Y.Y. Identification of higenamine in Radix Aconiti Lateralis Preparata as a β2-adrenergic receptor agonist. Acta Pharmacol. Sin. 2008, 29, 1187–1194. [Google Scholar] [CrossRef]
- Wu, M.P.; Zhang, Y.S.; Zhou, Q.M.; Xiong, J.; Dong, Y.R.; Yan, C. Higenamine protects ischemia/reperfusion induced cardiac injury and myocyte apoptosis through activation of β2-AR/PI3K/AKT signaling pathway. Pharmacol. Res. 2016, 104, 115–123. [Google Scholar] [CrossRef]
- DeFronzo, R.A.; Thorin, D.; Felber, J.P.; Simonson, D.C.; Thiebaud, D.; Jequier, E.; Golay, A. Effect of β and α adrenergic blockade on glucose-induced thermogenesis in man. J. Clin. Investig. 1984, 73, 633–639. [Google Scholar] [CrossRef]
- Abumrad, N.A. The Liver as a Hub in Thermogenesis. Cell Metab. 2017, 26, 454–455. [Google Scholar] [CrossRef]
- Simcox, J.; Geoghegan, G.; Maschek, J.A.; Bensard, C.L.; Pasquali, M.; Miao, R.; Lee, S.; Jiang, L.; Huck, I.; Kershaw, E.E.; et al. Global Analysis of Plasma Lipids Identifies Liver-Derived Acylcarnitines as a Fuel Source for Brown Fat Thermogenesis. Cell Metab. 2017, 26, 509–522. [Google Scholar] [CrossRef]
- Blachere, J.C.; Perusse, F.; Bukowiecki, L.J. Lowering plasma free fatty acids with Acipimox mimics the antidiabetic effects of the β3-adrenergic agonist CL-316243 in obese Zucker diabetic fatty rats. Metabolism 2001, 50, 945–951. [Google Scholar] [PubMed]
- Deng, G.; Schoch, T.D.; Cavicchi, K.A. Systematic Modification of the Glass Transition Temperature of Ion-Pair Comonomer Based Polyelectrolytes and Ionomers by Copolymerization with a Chemically Similar Cationic Monomer. Gels 2021, 7, 45. [Google Scholar] [CrossRef] [PubMed]
- Marsili, A.; Zavacki, A.M.; Harney, J.W.; Larsen, P.R. Physiological role and regulation of iodothyronine deiodinases: A 2011 update. J. Endocrinol. Investig. 2011, 34, 395–407. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Cai, H.; Shui, S.; Lin, Y.; Wang, F.; Wang, L.; Chen, J.; Liu, J. Chrysin Stimulates Subcutaneous Fat Thermogenesis in Mice by Regulating PDGFRα and MicroRNA Expressions. J. Agric. Food Chem. 2021, 69, 5897–5906. [Google Scholar] [CrossRef] [PubMed]
- Seale, L.A.; Ogawa-Wong, A.N.; Watanabe, L.M.; Khadka, V.S.; Menor, M.; Torres, D.J.; Carlson, B.A.; Hatfield, D.L.; Berry, M.J. Adaptive Thermogenesis in a Mouse Model Lacking Selenoprotein Biosynthesis in Brown Adipocytes. Int. J. Mol. Sci. 2021, 22, 611. [Google Scholar] [CrossRef]
- Chan, T.Y.; Tomlinson, B.; Tse, L.K.; Chan, J.C.; Chan, W.W.; Critchley, J.A. Aconitine poisoning due to Chinese herbal medicines: A review. Vet. Hum. Toxicol. 1994, 36, 452–455. [Google Scholar]
- Wang, S.; Jiang, X.; Xie, X.; Yin, J.; Zhang, J.; Liu, T.; Chen, S.; Wang, Y.; Zhou, X.; Wang, Y.; et al. piR-823 inhibits cell apoptosis via modulating mitophagy by binding to PINK1 in colorectal cancer. Cell Death Dis. 2022, 13, 465. [Google Scholar] [CrossRef]
- Shen, F.; Yang, W.; Cui, J.; Hou, Y.; Bai, G. Small-Molecule Fluorogenic Probe for the Detection of Mitochondrial Temperature In Vivo. Anal. Chem. 2021, 93, 13417–13420. [Google Scholar] [CrossRef]
- Zhang, W.; Gao, J.; Shen, F.; Ma, X.; Wang, Z.; Hou, X.; Hao, E.; Hou, Y.; Bai, G. Cinnamaldehyde changes the dynamic balance of glucose metabolism by targeting ENO. Life Sci. 2020, 258, 118151. [Google Scholar] [CrossRef]
- Kosaisawe, N.; Sparta, B.; Pargett, M.; Teragawa, C.K.; Albeck, J.G. Transient phases of OXPHOS inhibitor resistance reveal underlying metabolic heterogeneity in single cells. Cell Metab. 2021, 33, 649–665. [Google Scholar] [CrossRef]
- Ma, X.; Bai, Y.; Liu, K.; Han, Y.; Zhang, J.; Liu, Y.; Hou, X.; Hao, E.; Hou, Y.; Bai, G. Ursolic acid inhibits the cholesterol biosynthesis and alleviates high fat diet-induced hypercholesterolemia via irreversible inhibition of HMGCS1 in vivo. Phytomedicine 2022, 103, 154233. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wang, K.; Zhou, J.; Sullivan, M.A.; Liu, Y.; Gilbert, R.G.; Deng, B. Metformin and Berberine suppress glycogenolysis by inhibiting glycogen phosphorylase and stabilizing the molecular structure of glycogen in db/db mice. Carbohydr. Polym. 2020, 243, 116435. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, H.; Li, Z.; Cheng, C.; Cui, J.; Peng, J.; Wang, X.; Zhang, M.; Hou, Y.; Bai, G. Fuziline Ameliorates Glucose and Lipid Metabolism by Activating Beta Adrenergic Receptors to Stimulate Thermogenesis. Int. J. Mol. Sci. 2023, 24, 8362. https://doi.org/10.3390/ijms24098362
Gao H, Li Z, Cheng C, Cui J, Peng J, Wang X, Zhang M, Hou Y, Bai G. Fuziline Ameliorates Glucose and Lipid Metabolism by Activating Beta Adrenergic Receptors to Stimulate Thermogenesis. International Journal of Molecular Sciences. 2023; 24(9):8362. https://doi.org/10.3390/ijms24098362
Chicago/Turabian StyleGao, He, Zhenqiang Li, Chuanjing Cheng, Jing Cui, Jiamin Peng, Xiaoying Wang, Man Zhang, Yuanyuan Hou, and Gang Bai. 2023. "Fuziline Ameliorates Glucose and Lipid Metabolism by Activating Beta Adrenergic Receptors to Stimulate Thermogenesis" International Journal of Molecular Sciences 24, no. 9: 8362. https://doi.org/10.3390/ijms24098362