

Characterization of Antibodies against Receptor Activity-Modifying Protein 1 (RAMP1): A Cautionary Tale
 , , , ,
, , , ,  , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
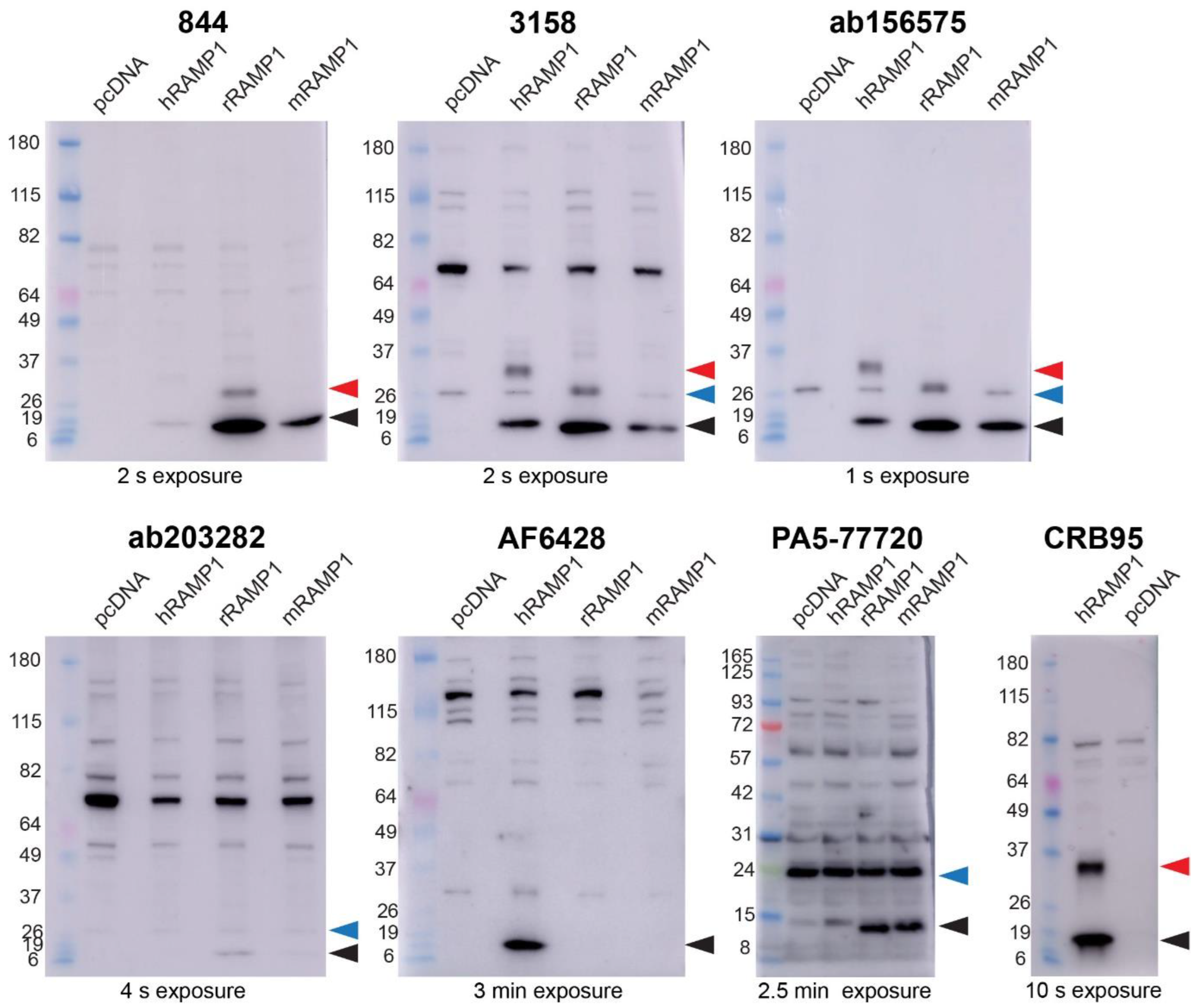
2.1. Anti-RAMP1 Antibodies Detected a Range of Bands in Transfected Cell Western Blots
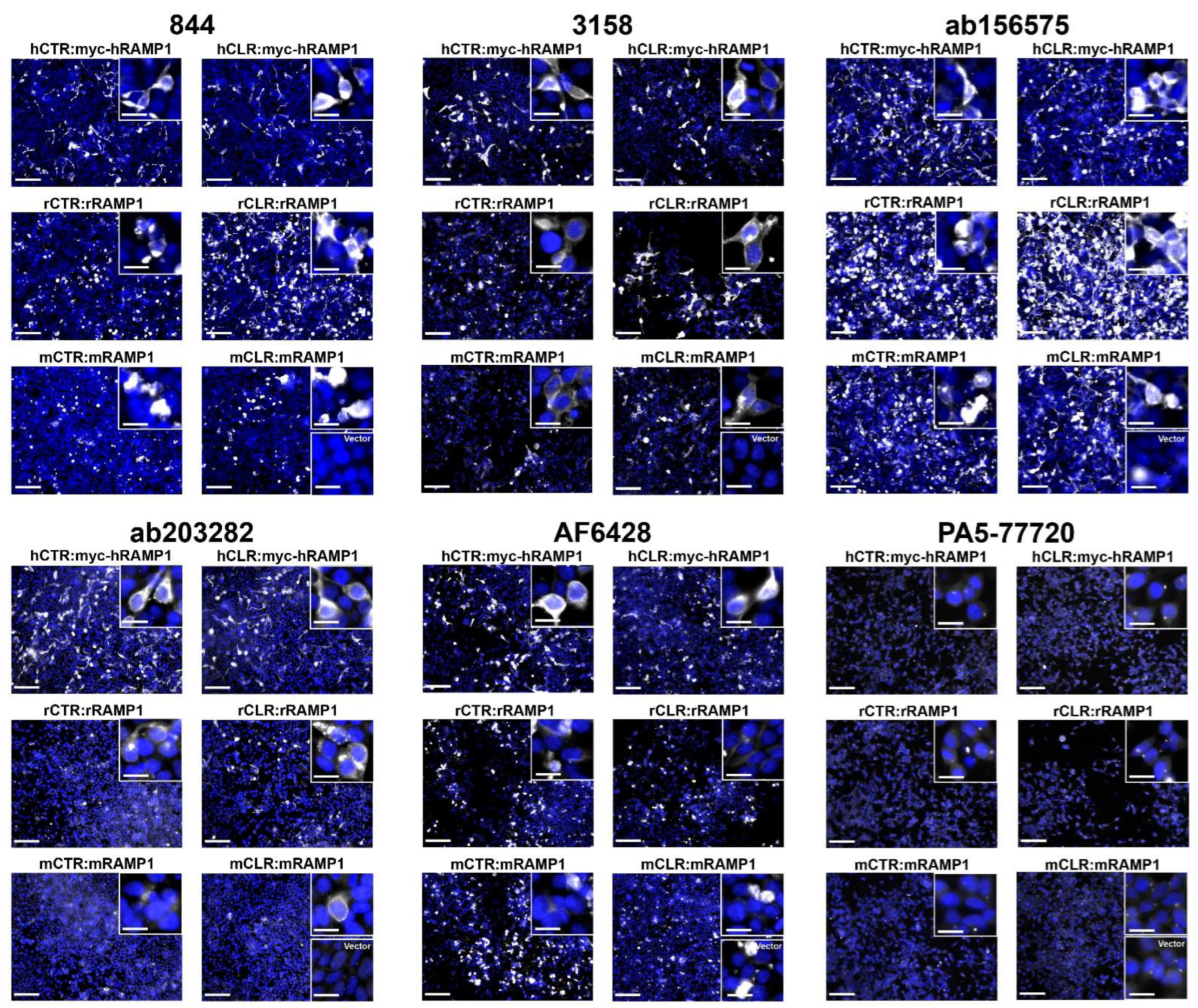
2.2. Anti-RAMP1 Antibodies Produced Immunoreactivity in RAMP1-Transfected Cells Using ICC
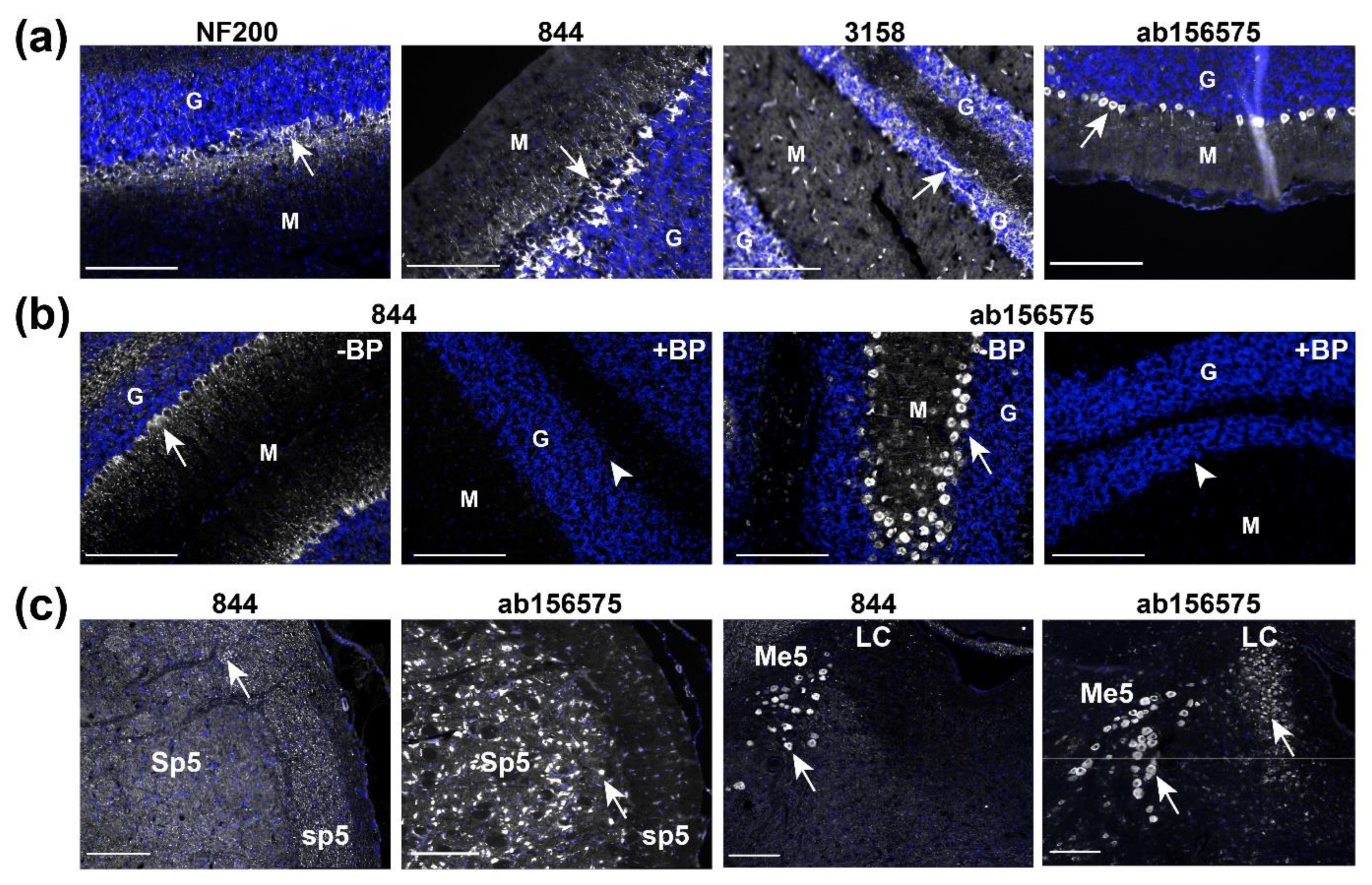
2.3. Anti-RAMP1 Antibodies Produced Immunoreactivity in Rat Brain Using IHC
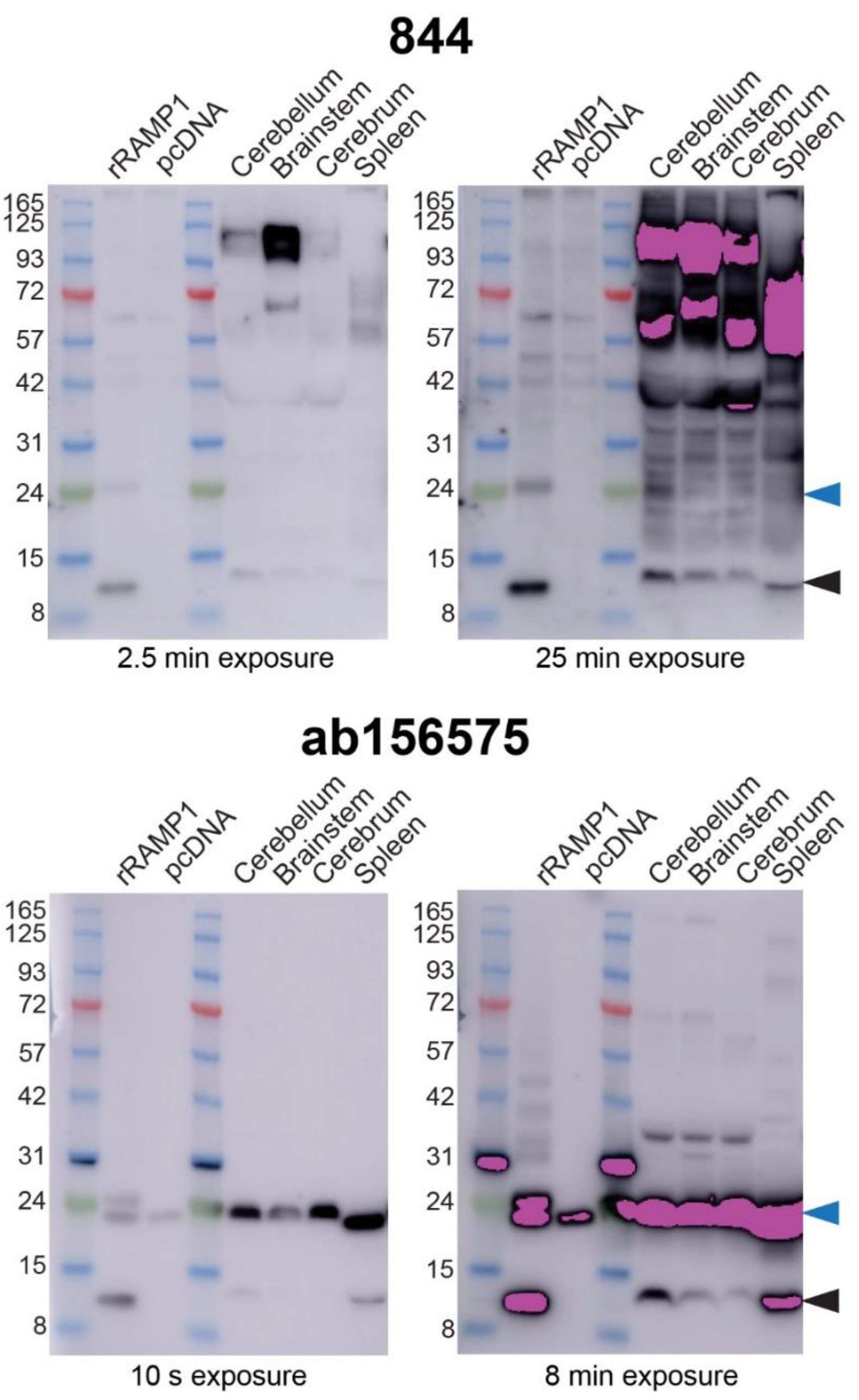
2.4. Anti-RAMP1 Antibodies Detected a Range of Bands in Rat Brain by Western Blotting
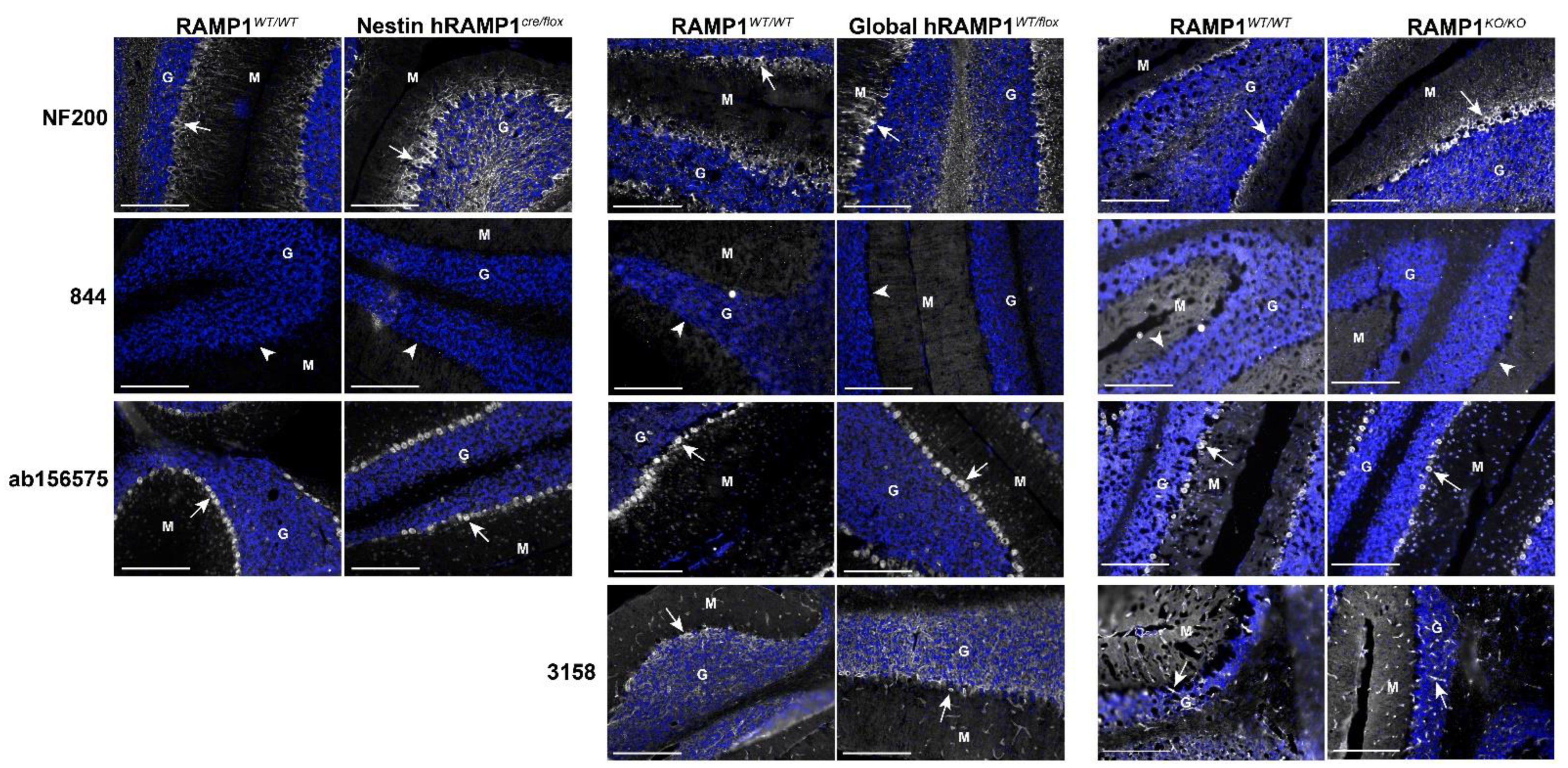
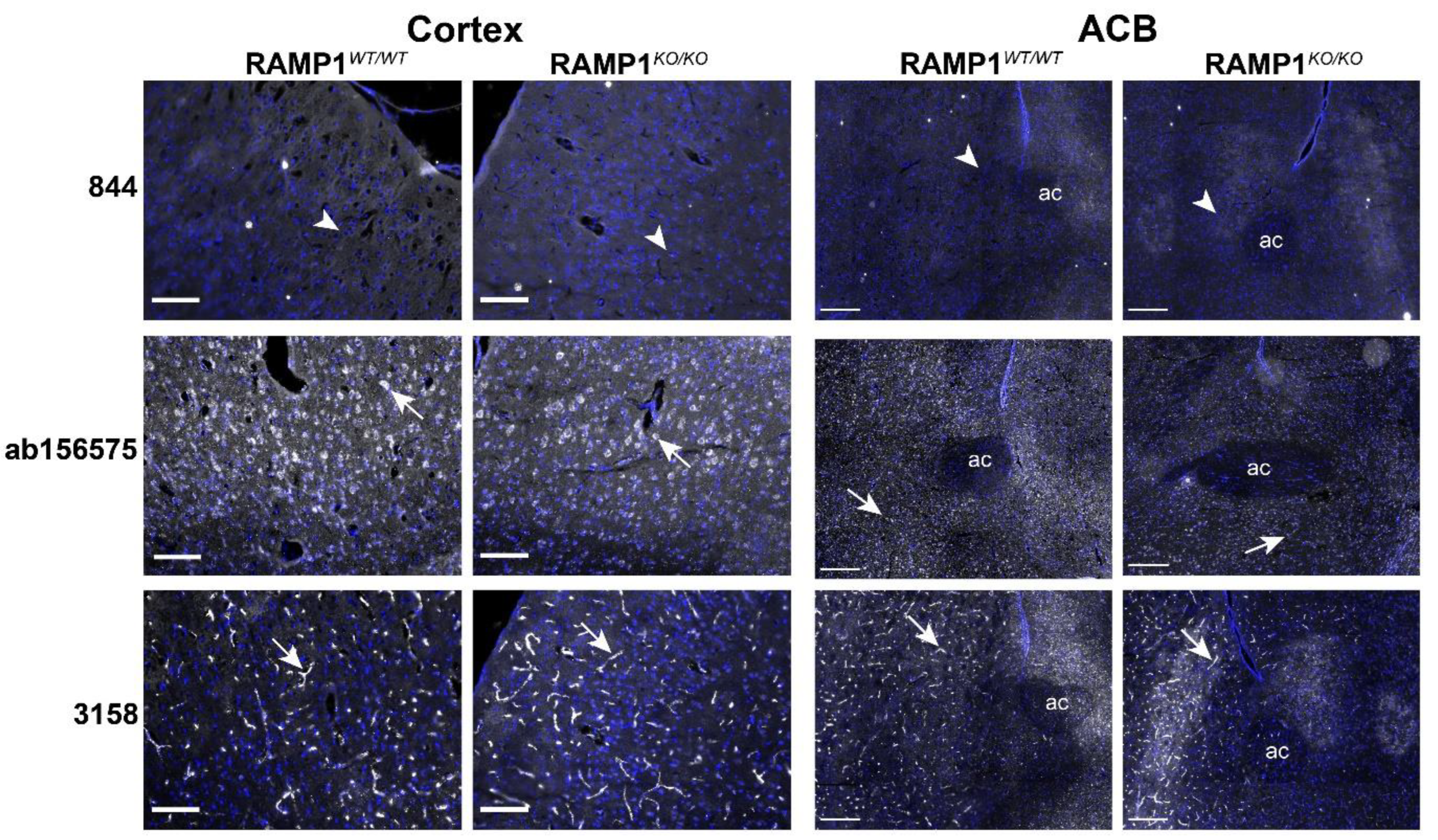
2.5. RAMP1 Antibodies Produced Different Immunofluorescence Patterns in Mouse Brain
2.6. Immunofluorescent Patterns in WT and Genetic RAMP1 Mouse Model Tissue
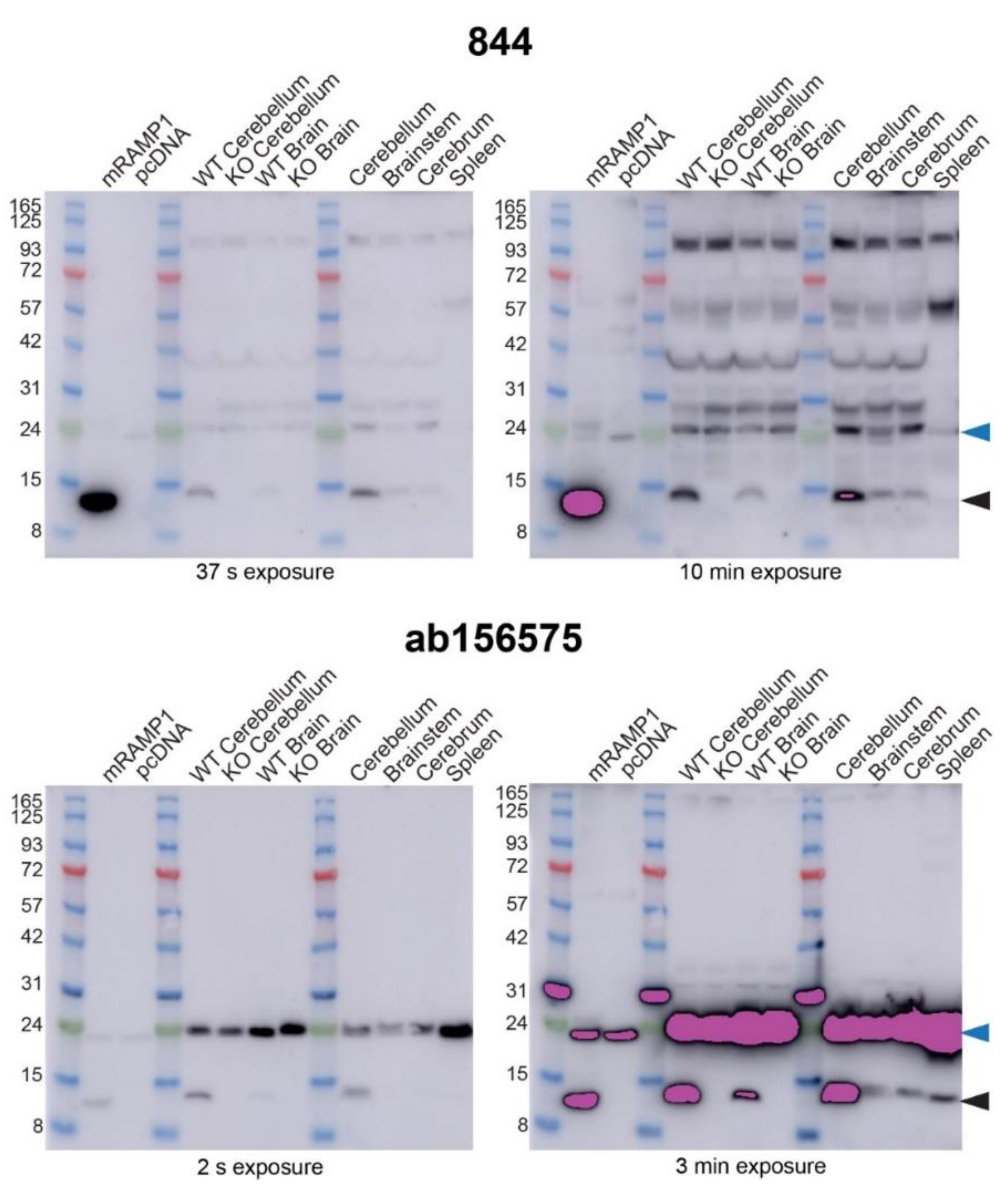
2.7. Anti-RAMP1 Antibodies Detected a Range of Bands in Mouse Brain by Western Blotting
3. Discussion
4. Materials and Methods
4.1. Antibodies
4.2. Animals and Tissue Collection
4.3. Rodent Tissue Sourced within New Zealand
4.4. Genetic Mouse Model Tissue
4.5. Cell Culture and Transfection
4.6. Western Blotting
4.7. ICC and IHC
4.8. Image Preparation and Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Deen, M.; Correnti, E.; Kamm, K.; Kelderman, T.; Papetti, L.; Rubio-Beltran, E.; Vigneri, S.; Edvinsson, L.; Maassen Van Den Brink, A.; European Headache Federation School of Advanced Studies (EHF-SAS). Blocking CGRP in migraine patients—A review of pros and cons. J. Headache Pain 2017, 18, 96. [Google Scholar] [CrossRef] [Green Version]
- Hay, D.L.; Garelja, M.L.; Poyner, D.R.; Walker, C.S. Update on the pharmacology of calcitonin/CGRP family of peptides: IUPHAR Review 25. Br. J. Pharmacol. 2018, 175, 3–17. [Google Scholar] [CrossRef]
- Walker, C.S.; Eftekhari, S.; Bower, R.L.; Wilderman, A.; Insel, P.A.; Edvinsson, L.; Waldvogel, H.J.; Jamaluddin, M.A.; Russo, A.F.; Hay, D.L. A second trigeminal CGRP receptor: Function and expression of the AMY1 receptor. Ann. Clin. Transl. Neurol. 2015, 2, 595–608. [Google Scholar] [CrossRef]
- Ghanizada, H.; Al-Karagholi, M.A.-M.; Walker, C.S.; Arngrim, N.; Rees, T.; Petersen, J.; Siow, A.; Mørch-Rasmussen, M.; Tan, S.; O’Carroll, S.J.; et al. Amylin Analog Pramlintide Induces Migraine-like Attacks in Patients. Ann. Neurol. 2021, 89, 1157–1171. [Google Scholar] [CrossRef]
- Tasma, Z.; Wills, P.; Hay, D.L.; Walker, C.S. Agonist bias and agonist-dependent antagonism at corticotrophin releasing factor receptors. Pharmacol. Res. Perspect. 2020, 8, e00595. [Google Scholar] [CrossRef]
- Lorenzen, E.; Dodig-Crnković, T.; Kotliar, I.B.; Pin, E.; Ceraudo, E.; Vaughan, R.D.; Uhlèn, M.; Huber, T.; Schwenk, J.M.; Sakmar, T.P. Multiplexed Analysis of the Secretin-like GPCR-RAMP Interactome. Sci. Adv. 2019, 5, eaaw2778. [Google Scholar] [CrossRef] [Green Version]
- Wootten, D.; Lindmark, H.; Kadmiel, M.; Willcockson, H.; Caron, K.M.; Barwell, J.; Drmota, T.; Poyner, D.R. Receptor activity modifying proteins (RAMPs) interact with the VPAC2 receptor and CRF1 receptors and modulate their function. Br. J. Pharmacol. 2013, 168, 822–834. [Google Scholar] [CrossRef] [Green Version]
- Shao, L.; Chen, Y.; Zhang, S.; Zhang, Z.; Cao, Y.; Yang, D.; Wang, M.-W. Modulating effects of RAMPs on signaling profiles of the glucagon receptor family. Acta Pharm. Sin. B 2022, 12, 637–650. [Google Scholar] [CrossRef] [PubMed]
- Sexton, P.; McKenzie, J.; Mason, R.; Martin, T.; Mendelsohn, F. Localization of binding sites for calcitonin gene-related peptide in rat brain by in vitro autoradiography. Neuroscience 1986, 19, 1235–1245. [Google Scholar] [CrossRef] [PubMed]
- Skofitsch, G.; Jacobowitz, D.M. Autoradiographic distribution of 125 I calcitonin gene-related peptide binding sites in the rat central nervous system. Peptides 1985, 6, 975–986. [Google Scholar] [CrossRef] [PubMed]
- Christopoulos, G.; Paxinos, G.; Huang, X.F.; Beaumont, K.; Toga, A.W.; Sexton, P.M. Comparative distribution of receptors for amylin and the related peptides calcitonin gene related peptide and calcitonin in rat and monkey brain. Can. J. Physiol. Pharmacol. 1995, 73, 1037–1041. [Google Scholar] [CrossRef]
- Hendrikse, E.R.; Bower, R.L.; Hay, D.L.; Walker, C.S. Molecular studies of CGRP and the CGRP family of peptides in the central nervous system. Cephalalgia 2019, 39, 403–419. [Google Scholar] [CrossRef]
- Oliver, K.R.; Kane, S.A.; Salvatore, C.A.; Mallee, J.J.; Kinsey, A.M.; Koblan, K.S.; Keyvan-Fouladi, N.; Heavens, R.P.; Wainwright, A.; Jacobson, M.; et al. Cloning, characterization and central nervous system distribution of receptor activity modifying proteins in the rat. Eur. J. Neurosci. 2001, 14, 618–628. [Google Scholar] [CrossRef] [PubMed]
- Eftekhari, S.; Salvatore, C.A.; Gaspar, R.C.; Roberts, R.; O’Malley, S.; Zeng, Z.; Edvinsson, L. Localization of CGRP receptor components, CGRP, and receptor binding sites in human and rhesus cerebellar cortex. Cerebellum 2013, 12, 937–949. [Google Scholar] [CrossRef] [PubMed]
- Ueda, T.; Ugawa, S.; Saishin, Y.; Shimada, S. Expression of receptor-activity modifying protein (RAMP) mRNAs in the mouse brain. Mol. Brain Res. 2001, 93, 36–45. [Google Scholar] [CrossRef]
- Liu, Y.; Beyer, A.; Aebersold, R. On the dependency of cellular protein levels on mRNA abundance. Cell 2016, 165, 535–550. [Google Scholar] [CrossRef] [Green Version]
- Tian, Q.; Stepaniants, S.B.; Mao, M.; Weng, L.; Feetham, M.C.; Doyle, M.J.; Yi, E.C.; Dai, H.; Thorsson, V.; Eng, J.; et al. Integrated genomic and proteomic analyses of gene expression in Mammalian cells. Mol. Cell. Proteom. 2004, 3, 960–969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warfvinge, K.; Edvinsson, L.; Pickering, D.S.; Sheykhzade, M. The Presence of Calcitonin Gene-Related Peptide and Its Receptors in Rat, Pig and Human Brain: Species Differences in Calcitonin Gene-Related Peptide Pharmacology. Pharmacology 2019, 104, 332–341. [Google Scholar] [CrossRef]
- Edvinsson, L.; Eftekhari, S.; Salvatore, C.A.; Warfvinge, K. Cerebellar distribution of calcitonin gene-related peptide (CGRP) and its receptor components calcitonin receptor-like receptor (CLR) and receptor activity modifying protein 1 (RAMP1) in rat. Mol. Cell. Neurosci. 2011, 46, 333–339. [Google Scholar] [CrossRef] [Green Version]
- Lennerz, J.K.; Ruhle, V.; Ceppa, E.P.; Neuhuber, W.L.; Bunnett, N.W.; Grady, E.F.; Messlinger, K. Calcitonin receptor-like receptor (CLR), receptor activity-modifying protein 1 (RAMP1), and calcitonin gene-related peptide (CGRP) immunoreactivity in the rat trigeminovascular system: Differences between peripheral and central CGRP receptor distribution. J. Comp. Neurol. 2008, 507, 1277–1299. [Google Scholar] [CrossRef]
- Uhlen, M.; Bandrowski, A.; Carr, S.; Edwards, A.; Ellenberg, J.; Lundberg, E.; Rimm, D.L.; Rodriguez, H.; Hiltke, T.; Snyder, M.; et al. A proposal for validation of antibodies. Nat. Methods 2016, 13, 823–827. [Google Scholar] [CrossRef]
- Hewitt, S.M.; Baskin, D.G.; Frevert, C.W.; Stahl, W.L.; Rosa-Molinar, E. Controls for immunohistochemistry: The Histochemical Society’s standards of practice for validation of immunohistochemical assays. J. Histochem. Cytochem. 2014, 62, 693–697. [Google Scholar] [CrossRef] [Green Version]
- Eftekhari, S.; Salvatore, C.A.; Calamari, A.; Kane, S.A.; Tajti, J.; Edvinsson, L. Differential distribution of calcitonin gene-related peptide and its receptor components in the human trigeminal ganglion. Neuroscience 2010, 169, 683–696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McLatchie, L.M.; Fraser, N.J.; Main, M.J.; Wise, A.; Brown, J.; Thompson, N.; Solari, R.; Lee, M.G.; Foord, S.M. RAMPs regulate the transport and ligand specificity of the calcitonin-receptor-like receptor. Nature 1998, 393, 333–339. [Google Scholar] [CrossRef]
- Rees, T.A.; Russo, A.F.; O’Carroll, S.J.; Hay, D.L.; Walker, C.S. CGRP and the Calcitonin Receptor are Co-Expressed in Mouse, Rat and Human Trigeminal Ganglia Neurons. Front. Physiol. 2022, 13, 860037. [Google Scholar] [CrossRef]
- Uhlen, M. Response to: Should we ignore western blots when selecting antibodies for other applications? Nat. Methods 2017, 14, 215–216. [Google Scholar] [CrossRef] [PubMed]
- Warfvinge, K.; Edvinsson, L. Distribution of CGRP and CGRP receptor components in the rat brain. Cephalalgia 2017, 39, 0333102417728873. [Google Scholar] [CrossRef]
- Inagaki, S.; Kito, S.; Kubota, Y.; Girgis, S.; Hillyard, C.J.; Macintyre, I. Autoradiographic localization of calcitonin gene-related peptide binding sites in human and rat brains. Brain Res. 1986, 374, 287–298. [Google Scholar] [CrossRef]
- Eftekhari, S.; Gaspar, R.C.; Roberts, R.; Chen, T.B.; Zeng, Z.; Villarreal, S.; Edvinsson, L.; Salvatore, C.A. Localization of CGRP receptor components and receptor binding sites in rhesus monkey brainstem: A detailed study using in situ hybridization, immunofluorescence, and autoradiography. J. Comp. Neurol. 2016, 524, 90–118. [Google Scholar] [CrossRef]
- Eftekhari, S.; Edvinsson, L. Calcitonin gene-related peptide (CGRP) and its receptor components in human and rat spinal trigeminal nucleus and spinal cord at C1-level. BMC Neurosci. 2011, 12, 112. [Google Scholar] [CrossRef] [PubMed]
- Holland, P.R.; Saengjaroentham, C.; Vila-Pueyo, M. The role of the brainstem in migraine: Potential brainstem effects of CGRP and CGRP receptor activation in animal models. Cephalalgia 2019, 39, 390–402. [Google Scholar] [CrossRef] [PubMed]
- Nagae, T.; Mukoyama, M.; Sugawara, A.; Mori, K.; Yahata, K.; Kasahara, M.; Suganami, T.; Makino, H.; Fujinaga, Y.; Yoshioka, T.; et al. Rat receptor-activity-modifying proteins (RAMPs) for adrenomedullin/CGRP receptor: Cloning and upregulation in obstructive nephropathy. Biochem. Biophys. Res. Commun. 2000, 270, 89–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Insel, P.A.; Sriram, K.; Gorr, M.W.; Wiley, S.Z.; Michkov, A.; Salmerón, C.; Chinn, A.M. GPCRomics: An approach to discover GPCR drug targets. Trends Pharmacol. Sci. 2019, 40, 378–387. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Winborn, C.S.; De Prado, B.M.; Russo, A.F. Sensitization of calcitonin gene-related peptide receptors by receptor activity-modifying protein-1 in the trigeminal ganglion. J. Neurosci. 2007, 27, 2693–2703. [Google Scholar] [CrossRef] [Green Version]
- Bohn, K.J.; Li, B.; Huang, X.; Mason, B.N.; Wattiez, A.S.; Kuburas, A.; Walker, C.S.; Yang, P.; Yu, J.; Heinz, B.A.; et al. CGRP receptor activity in mice with global expression of human receptor activity modifying protein 1. Br. J. Pharmacol. 2017, 174, 1826–1840. [Google Scholar] [CrossRef] [Green Version]
- Coester, B.; Pence, S.W.; Arrigoni, S.; Boyle, C.N.; Le Foll, C.; Lutz, T.A. RAMP1 and RAMP3 Differentially Control Amylin’s Effects on Food Intake, Glucose and Energy Balance in Male and Female Mice. Neuroscience 2020, 447, 74–93. [Google Scholar] [CrossRef]
- Li, M.; Wetzel-Strong, S.E.; Hua, X.; Tilley, S.L.; Oswald, E.; Krummel, M.F.; Caron, K.M. Deficiency of RAMP1 attenuates antigen-induced airway hyperresponsiveness in mice. PLoS ONE 2014, 9, e102356. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Liu, X.; Morgan, D.A.; Kuburas, A.; Thedens, D.R.; Russo, A.F.; Rahmouni, K. Neuronal Receptor Activity–Modifying Protein 1 Promotes Energy Expenditure in Mice. Diabetes 2011, 60, 1063–1071. [Google Scholar] [CrossRef] [Green Version]
- Barth, S.W.; Riediger, T.; Lutz, T.A.; Rechkemmer, G. Peripheral amylin activates circumventricular organs expressing calcitonin receptor a/b subtypes and receptor-activity modifying proteins in the rat. Brain Res. 2004, 997, 97–102. [Google Scholar] [CrossRef]
- Kalafateli, A.L.; Vallöf, D.; Colombo, G.; Lorrai, I.; Maccioni, P.; Jerlhag, E. An amylin analogue attenuates alcohol-related behaviours in various animal models of alcohol use disorder. Neuropsychopharmacology 2019, 44, 1093–1102. [Google Scholar] [CrossRef]
- Baisley, S.K.; Bremer, Q.Z.; Bakshi, V.P.; Baldo, B.A. Antipsychotic-like actions of the satiety peptide, amylin, in ventral striatal regions marked by overlapping calcitonin receptor and RAMP-1 gene expression. J. Neurosci. 2014, 34, 4318–4325. [Google Scholar] [CrossRef] [Green Version]
- Saper, C.B. A guide to the perplexed on the specificity of antibodies. J. Histochem. Cytochem. 2009, 57, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Hendrikse, E.R.; Rees, T.A.; Tasma, Z.; Le Foll, C.; Lutz, T.A.; Siow, A.; Wookey, P.J.; Walker, C.S.; Hay, D.L. Calcitonin receptor antibody validation and expression in the rodent brain. Cephalalgia 2022, 42, 815–826. [Google Scholar] [CrossRef] [PubMed]
- Moye, L.S.; Siegersma, K.; Dripps, I.; Witkowski, W.; Mangutov, E.; Wang, D.; Scherrer, G.; Pradhan, A.A. Delta opioid receptor regulation of calcitonin gene-related peptide dynamics in the trigeminal complex. Pain 2021, 162, 2297–2308. [Google Scholar] [CrossRef] [PubMed]
- Vilotti, S.; Vana, N.; Van den Maagdenberg, A.M.; Nistri, A. Expression and function of calcitonin gene-related peptide (CGRP) receptors in trigeminal ganglia of R192Q Cacna1a knock-in mice. Neurosci. Lett. 2016, 620, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Ernstsen, C.; Christensen, S.L.; Rasmussen, R.H.; Nielsen, B.S.; Jansen-Olesen, I.; Olesen, J.; Kristensen, D.M. The PACAP pathway is independent of CGRP in mouse models of migraine: Possible new drug target? Brain 2022, 145, 2450–2460. [Google Scholar] [CrossRef]
- Argunhan, F.; Thapa, D.; Aubdool, A.A.; Carlini, E.; Arkless, K.; Hendrikse, E.R.; Valente, J.d.S.; Kodji, X.; Barrett, B.; Ricciardi, C.A.; et al. Calcitonin Gene-Related Peptide Protects Against Cardiovascular Dysfunction Independently of Nitric Oxide In Vivo. Hypertension 2021, 77, 1178–1190. [Google Scholar] [CrossRef]
- Aranäs, C.; Vestlund, J.; Witley, S.; Edvardsson, C.E.; Kalafateli, A.L.; Jerlhag, E. Salmon Calcitonin Attenuates Some Behavioural Responses to Nicotine in Male Mice. Front. Pharmacol. 2021, 12, 685631. [Google Scholar] [CrossRef]
- De Logu, F.; Nassini, R.; Hegron, A.; Landini, L.; Jensen, D.D.; Latorre, R.; Ding, J.; Marini, M.; Souza Monteiro de Araujo, D.; Ramírez-Garcia, P.; et al. Schwann cell endosome CGRP signals elicit periorbital mechanical allodynia in mice. Nat. Commun. 2022, 13, 646. [Google Scholar] [CrossRef]
- Uhlén, M.; Fagerberg, L.; Hallström, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, Å.; Kampf, C.; Sjöstedt, E.; Asplund, A. Tissue-based map of the human proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef]
- Goemaere, J.; Knoops, B. Peroxiredoxin distribution in the mouse brain with emphasis on neuronal populations affected in neurodegenerative disorders. J. Comp. Neurol. 2012, 520, 258–280. [Google Scholar] [CrossRef] [PubMed]
- Kunz, T.H.; Mueller-Steiner, S.; Schwerdtfeger, K.; Kleinert, P.; Troxler, H.; Kelm, J.M.; Ittner, L.M.; Fischer, J.A.; Born, W. Interaction of receptor-activity-modifying protein1 with tubulin. Biochim. Biophys. Acta BBA Gen. Subj. 2007, 1770, 1145–1150. [Google Scholar] [CrossRef] [PubMed]
- Hostetler, E.D.; Joshi, A.D.; Sanabria-Bohorquez, S.; Fan, H.; Zeng, Z.; Purcell, M.; Gantert, L.; Riffel, K.; Williams, M.; O’Malley, S.; et al. In vivo quantification of calcitonin gene-related peptide receptor occupancy by telcagepant in rhesus monkey and human brain using the positron emission tomography tracer [11C]MK-4232. J. Pharmacol. Exp. Ther. 2013, 347, 478–486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bussiere, J.L.; Davies, R.; Dean, C.; Xu, C.; Kim, K.H.; Vargas, H.M.; Chellman, G.J.; Balasubramanian, G.; Rubio-Beltran, E.; Maassen Van Den Brink, A. Nonclinical safety evaluation of erenumab, a CGRP receptor inhibitor for the prevention of migraine. Regul. Toxicol. Pharmacol. 2019, 106, 224–238. [Google Scholar] [CrossRef] [PubMed]
- Miller, S.; Liu, H.; Warfvinge, K.; Shi, L.; Dovlatyan, M.; Xu, C.; Edvinsson, L. Immunohistochemical localization of the calcitonin gene-related peptide binding site in the primate trigeminovascular system using functional antagonist antibodies. Neuroscience 2016, 328, 165–183. [Google Scholar] [CrossRef] [PubMed]
- Bhakta, M.; Vuong, T.; Taura, T.; Wilson, D.S.; Stratton, J.R.; Mackenzie, K.D. Migraine therapeutics differentially modulate the CGRP pathway. Cephalalgia 2021, 41, 499–514. [Google Scholar] [CrossRef] [PubMed]
- Hage La Cour, S.; Juhler, K.; Kogelman, L.J.A.; Olesen, J.; Klaerke, D.A.; Kristensen, D.M.; Jansen-Olesen, I. Characterization of erenumab and rimegepant on calcitonin gene-related peptide induced responses in Xenopus Laevis oocytes expressing the calcitonin gene-related peptide receptor and the amylin-1 receptor. J. Headache Pain 2022, 23, 59. [Google Scholar] [CrossRef]
- Shi, L.; Lehto, S.G.; Zhu, D.X.; Sun, H.; Zhang, J.; Smith, B.P.; Immke, D.C.; Wild, K.D.; Xu, C. Pharmacologic Characterization of AMG 334, a Potent and Selective Human Monoclonal Antibody against the Calcitonin Gene-Related Peptide Receptor. J. Pharmacol. Exp. Ther. 2016, 356, 223–231. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Xu, M.; Bhuiyan, S.A.; Li, J.; Zhao, J.; Cohrs, R.J.; Susterich, J.T.; Signorelli, S.; Green, U.; Stone, J.R.; et al. Human and mouse trigeminal ganglia cell atlas implicates multiple cell types in migraine. Neuron 2022, 110, 1806–1821.E8. [Google Scholar] [CrossRef]
- Coester, B.; Koester-Hegmann, C.; Lutz, T.A.; Le Foll, C. Amylin/Calcitonin Receptor-Mediated Signaling in POMC Neurons Influences Energy Balance and Locomotor Activity in Chow-Fed Male Mice. Diabetes 2020, 69, 1110–1125. [Google Scholar] [CrossRef]
- Percie du Sert, N.; Hurst, V.; Ahluwalia, A.; Alam, S.; Avey, M.T.; Baker, M.; Browne, W.J.; Clark, A.; Cuthill, I.C.; Dirnagl, U.; et al. The ARRIVE guidelines 2.0: Updated guidelines for reporting animal research. J. Cereb. Blood Flow Metab. 2020, 40, 1769–1777. [Google Scholar] [CrossRef]
- Pawlak, J.; Wetzel-Strong, S.; Dunn, M.; Caron, K. Cardiovascular effects of exogenous adrenomedullin and CGRP in Ramp and Calcrl deficient mice. Peptides 2017, 88, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Gingell, J.J.; Rees, T.A.; Hendrikse, E.R.; Siow, A.; Rennison, D.; Scotter, J.; Harris, P.W.; Brimble, M.A.; Walker, C.S.; Hay, D.L. Distinct patterns of internalization of different calcitonin gene-related peptide receptors. ACS Pharmacol. Transl. Sci. 2020, 3, 296–304. [Google Scholar] [CrossRef]
- Bailey, R.J.; Hay, D.L. Pharmacology of the human CGRP1 receptor in Cos 7 cells. Peptides 2006, 27, 1367–1375. [Google Scholar] [CrossRef] [PubMed]
- Bower, R.L.; Yule, L.; Rees, T.A.; Deganutti, G.; Hendrikse, E.R.; Harris, P.W.; Kowalczyk, R.; Ridgway, Z.; Wong, A.G.; Swierkula, K. Molecular signature for receptor engagement in the metabolic peptide hormone amylin. ACS Pharmacol. Transl. Sci. 2018, 1, 32–49. [Google Scholar] [CrossRef]
- Qi, T.; Dong, M.; Watkins, H.A.; Wootten, D.; Miller, L.J.; Hay, D.L. Receptor activity-modifying protein-dependent impairment of calcitonin receptor splice variant Δ(1–47)hCT(a) function. Br. J. Pharmacol. 2013, 168, 644–657. [Google Scholar] [CrossRef] [Green Version]
- Garelja, M.L.; Bower, R.L.; Brimble, M.A.; Chand, S.; Harris, P.W.R.; Jamaluddin, M.A.; Petersen, J.; Siow, A.; Walker, C.S.; Hay, D.L. Pharmacological characterisation of mouse calcitonin and calcitonin receptor-like receptors reveals differences compared with human receptors. Br. J. Pharmacol. 2022, 179, 416–434. [Google Scholar] [CrossRef]
- Preibisch, S.; Saalfeld, S.; Tomancak, P. Globally optimal stitching of tiled 3D microscopic image acquisitions. Bioinformatics 2009, 25, 1463–1465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, J. Not seeing is not believing: Improving the visibility of your fluorescence images. Mol. Biol. Cell 2012, 23, 754–757. [Google Scholar] [CrossRef] [PubMed]
- Lein, E.S.; Hawrylycz, M.J.; Ao, N.; Ayres, M.; Bensinger, A.; Bernard, A.; Boe, A.F.; Boguski, M.S.; Brockway, K.S.; Byrnes, E.J.; et al. Genome-wide atlas of gene expression in the adult mouse brain. Nature 2007, 445, 168–176. [Google Scholar] [CrossRef] [PubMed]
- Edvinsson, J.C.A.; Warfvinge, K.; Krause, D.N.; Blixt, F.W.; Sheykhzade, M.; Edvinsson, L.; Haanes, K.A. C-fibers may modulate adjacent Adelta-fibers through axon-axon CGRP signaling at nodes of Ranvier in the trigeminal system. J. Headache Pain 2019, 20, 105. [Google Scholar] [CrossRef] [Green Version]
- Csati, A.; Tajti, J.; Tuka, B.; Edvinsson, L.; Warfvinge, K. Calcitonin gene-related peptide and its receptor components in the human sphenopalatine ganglion–interaction with the sensory system. Brain Res. 2012, 1435, 29–39. [Google Scholar] [CrossRef]
- Edvinsson, L.; Grell, A.S.; Warfvinge, K. Expression of the CGRP Family of Neuropeptides and their Receptors in the Trigeminal Ganglion. J. Mol. Neurosci. 2020, 70, 930–944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eftekhari, S.; Salvatore, C.A.; Johansson, S.; Chen, T.B.; Zeng, Z.; Edvinsson, L. Localization of CGRP, CGRP receptor, PACAP and glutamate in trigeminal ganglion. Relation to the blood-brain barrier. Brain Res. 2015, 1600, 93–109. [Google Scholar] [CrossRef]
- Eftekhari, S.; Warfvinge, K.; Blixt, F.W.; Edvinsson, L. Differentiation of nerve fibers storing CGRP and CGRP receptors in the peripheral trigeminovascular system. J. Pain 2013, 14, 1289–1303. [Google Scholar] [CrossRef] [Green Version]
- Filiz, A.; Tepe, N.; Eftekhari, S.; Boran, H.E.; Dilekoz, E.; Edvinsson, L.; Bolay, H. CGRP receptor antagonist MK-8825 attenuates cortical spreading depression induced pain behavior. Cephalalgia 2019, 39, 354–365. [Google Scholar] [CrossRef]
- Pozo-Rosich, P.; Storer, R.J.; Charbit, A.R.; Goadsby, P.J. Periaqueductal gray calcitonin gene-related peptide modulates trigeminovascular neurons. Cephalalgia 2015, 35, 1298–1307. [Google Scholar] [CrossRef] [PubMed]
- Summ, O.; Charbit, A.R.; Andreou, A.P.; Goadsby, P.J. Modulation of nocioceptive transmission with calcitonin gene-related peptide receptor antagonists in the thalamus. Brain 2010, 133, 2540–2548. [Google Scholar] [CrossRef]
- Warfvinge, K.; Krause, D.; Edvinsson, L. The distribution of oxytocin and the oxytocin receptor in rat brain: Relation to regions active in migraine. J. Headache Pain 2020, 21, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, Y.; Ding, L.; Dai, H.; Wang, F.; Zhou, H.; Gao, Q.; Xiong, X.; Zhang, F.; Song, T.; Yuan, Y.; et al. Intermedin in Paraventricular Nucleus Attenuates Ang II-Induced Sympathoexcitation through the Inhibition of NADPH Oxidase-Dependent ROS Generation in Obese Rats with Hypertension. Int. J. Mol. Sci. 2019, 20, 4217. [Google Scholar] [CrossRef]
- Blixt, F.W.; Radziwon-Balicka, A.; Edvinsson, L.; Warfvinge, K. Distribution of CGRP and its receptor components CLR and RAMP1 in the rat retina. Exp. Eye Res. 2017, 161, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Sheykhzade, M.; Amandi, N.; Pla, M.V.; Abdolalizadeh, B.; Sams, A.; Warfvinge, K.; Edvinsson, L.; Pickering, D.S. Binding and functional pharmacological characteristics of gepant-type antagonists in rat brain and mesenteric arteries. Vascul. Pharmacol. 2017, 90, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Seiler, K.; Nusser, J.I.; Lennerz, J.K.; Neuhuber, W.L.; Messlinger, K. Changes in calcitonin gene-related peptide (CGRP) receptor component and nitric oxide receptor (sGC) immunoreactivity in rat trigeminal ganglion following glyceroltrinitrate pretreatment. J. Headache Pain 2013, 14, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.; Cai, X.; Liu, X.; Guo, S.W. Possible involvement of neuropeptide and neurotransmitter receptors in Adenomyosis. Reprod. Biol. Endocrinol. 2021, 19, 25. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.Y.; Xu, X.M.; Wu, S.Y.; Wang, F.; Zhang, Z.C.; Yang, Y.L.; Li, M.; Wei, X.Z. Low-Intensity Pulsed Ultrasound-Induced Spinal Fusion is Coupled with Enhanced Calcitonin Gene-Related Peptide Expression in Rat Model. Ultrasound Med. Biol. 2017, 43, 1486–1493. [Google Scholar] [CrossRef] [PubMed]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Soding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef]
- UniProt, C. UniProt: The universal protein knowledgebase in 2021. Nucleic Acids Res. 2021, 49, D480–D489. [Google Scholar] [CrossRef]
- Bass, J.J.; Wilkinson, D.J.; Rankin, D.; Phillips, B.E.; Szewczyk, N.J.; Smith, K.; Atherton, P.J. An overview of technical considerations for Western blotting applications to physiological research. Scand. J. Med. Sci. Sports 2017, 27, 4–25. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | RAMP1 Band (~14 kDa) [Black] | Dimeric RAMP1 Band [Red] | Additional ~24 kDa Band (Non-Specific) [Blue] & |
|---|---|---|---|
| 844 | human, rat, mouse | human ^, rat * | No |
| 3158 | human, rat, mouse | human ^ | Yes |
| ab156575 | human, rat, mouse | human ^, rat * | Yes |
| ab203282 | rat, mouse | No | Yes |
| AF6428 | human | No | No |
| PA5-77720 | rat, mouse | No | Yes |
| CRB95† | human | human ^ | No |
| Antibody | ICC | Rat | Mouse | Mouse Overexpression | Mouse KO | |||
|---|---|---|---|---|---|---|---|---|
| IHC | WB # | IHC | WB # | IHC | IHC | WB # | ||
| 844 | Detected h/r/m RAMP1 | Between Gran/Mol layers (Cb) | Cb, BS, Cortex, Spleen | No IR | Cb, BS, Cortex, Spleen | No IR, no difference to WT | No IR, no difference to WT | RAMP1 monomer absent in KO |
| 3158 | Detected h/r/m RAMP1 | Granular layer (Cb) | NP | Cb, Cortex, ACB | NP | NP | No difference to WT | NP |
| ab156575 | Detected h/r/m RAMP1 | Between Gran/Mol layers (Cb) | Cb, BS, Cortex, Spleen | Nuclei/cell bodies in Cb, Cortex and ACB | Cb, BS, Cortex, Spleen | No difference to WT | No difference to WT | RAMP1 monomer absent in KO |
| ab203282 | Detected h/r RAMP1 | NP | NP | NP | NP | NP | No difference to WT | NP |
| AF6428 | Detected hRAMP1 ^ | NP | NP | NP | NP | No IR, no difference to WT | NP | NP |
| PA5-77720 | Did not detect RAMP1 | NP | NP | NP | NP | NP | NP | NP |
| CRB95 | Detected h/r RAMP1 * | Between Gran/Mol layers (Cb) | NP | NP | NP | NP | NP | NP |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hendrikse, E.R.; Rees, T.A.; Tasma, Z.; Garelja, M.L.; Siow, A.; Harris, P.W.R.; Pawlak, J.B.; Caron, K.M.; Blakeney, E.S.; Russo, A.F.; et al. Characterization of Antibodies against Receptor Activity-Modifying Protein 1 (RAMP1): A Cautionary Tale. Int. J. Mol. Sci. 2022, 23, 16035. https://doi.org/10.3390/ijms232416035
Hendrikse ER, Rees TA, Tasma Z, Garelja ML, Siow A, Harris PWR, Pawlak JB, Caron KM, Blakeney ES, Russo AF, et al. Characterization of Antibodies against Receptor Activity-Modifying Protein 1 (RAMP1): A Cautionary Tale. International Journal of Molecular Sciences. 2022; 23(24):16035. https://doi.org/10.3390/ijms232416035
Chicago/Turabian StyleHendrikse, Erica R., Tayla A. Rees, Zoe Tasma, Michael L. Garelja, Andrew Siow, Paul W. R. Harris, John B. Pawlak, Kathleen M. Caron, Elizabeth S. Blakeney, Andrew F. Russo, and et al. 2022. "Characterization of Antibodies against Receptor Activity-Modifying Protein 1 (RAMP1): A Cautionary Tale" International Journal of Molecular Sciences 23, no. 24: 16035. https://doi.org/10.3390/ijms232416035