
Mitochondrial Respiratory Chain Supercomplexes: From Structure to Function

Beijing Institute of Radiation Medicine, Beijing 100850, China
*
Authors to whom correspondence should be addressed.
Int. J. Mol. Sci. 2022, 23(22), 13880; https://doi.org/10.3390/ijms232213880
Submission received: 8 October 2022
/
Revised: 1 November 2022
/
Accepted: 7 November 2022
/
Published: 10 November 2022
(This article belongs to the Section Molecular Biophysics)
Abstract
:Mitochondrial oxidative phospho rylation, the center of cellular metabolism, is pivotal for the energy production in eukaryotes. Mitochondrial oxidative phosphorylation relies on the mitochondrial respiratory chain, which consists of four main enzyme complexes and two mobile electron carriers. Mitochondrial enzyme complexes also assemble into respiratory chain supercomplexes (SCs) through specific interactions. The SCs not only have respiratory functions but also improve the efficiency of electron transfer and reduce the production of reactive oxygen species (ROS). Impaired assembly of SCs is closely related to various diseases, especially neurodegenerative diseases. Therefore, SCs play important roles in improving the efficiency of the mitochondrial respiratory chain, as well as maintaining the homeostasis of cellular metabolism. Here, we review the structure, assembly, and functions of SCs, as well as the relationship between mitochondrial SCs and diseases.
1. Introduction
The mitochondrial respiratory chain is the basic structure for oxidative phosphorylation and plays a central role in cellular energy metabolism [1]. The mitochondrial respiratory chain consists of four enzyme complexes, including a nicotinamide adenine dinucleotide ubiquinone reductase (NADH dehydrogenase) (Complex I, CI), a succinate ubiquinone oxidoreductase (Complex II, CII), ubiquinone cytochrome oxidoreductase (Complex III, CIII), and cytochrome c oxidase (Complex IV, CIV), as well as two mobile electron carriers, ubiquinone (Co Q) and cytochrome c (Cyt c). These enzyme complexes are embedded in the inner mitochondrial membrane to form a continuous reaction system. Generally, the mitochondrial respiratory chain transfers protons and electrons from substrates to the final electronic acceptor, oxygen, and generates energy to support the phosphorylation of adenosine diphosphate (ADP) (Figure 1).
Over the past decades, models of the organization of mitochondrial respiratory chain have been controversial [2,3]. A solid model and the liquid model are two typical models for the organization of mitochondrial respiratory chain. The solid model (Figure 2A) assumes the complexes of the mitochondrial respiratory chain are anchored within a framework to ensure contact between each other with high catalytic activity [2,4]. However, with the purification of functional respiratory complexes and new understanding of the fluid nature of the mitochondrial energy transducing membrane, the liquid model (Figure 2B) has gained acceptance [5,6]. This model assumes that the components of mitochondrial respiratory chain, which participate in electron transfer, are distributed independently and are freely diffusible. Electron transfer is a multi-collisional, hindered, and long-distant diffusion process [2,7]. Compared to the solid model, the liquid model has been widely accepted.
In 2000, Schägger et al. [8] found mitochondrial supercomplexes (SCs)for the first time by blue native-page gel electrophoresis (BN-PAGE), and the enzyme activity of SCs was also confirmed. SCs are functional quaternary structures consisting of several free complexes both in prokaryotes and eukaryotes [9,10]. Compared with a free complex, SCs increase the efficiency of electron transport, reduce the production of reactive oxygen species (ROS), and stabilize the structure of free complexes [11,12,13,14]. Further studies suggested that free complexes and SCs are co-located in the inner mitochondrial membrane, have enzymatic activity and exert respiratory functions [15,16,17,18]. Moreover, SCs can be observed in BN-PAGE bands from different detergent-treated protein samples, which excludes the possibility that SCs were produced by electrophoretic techniques [15]. Furthermore, SCs have been observed in mitochondria from other species, such as yeast, plants, and vertebrates, by BN-PAGE [2]. Therefore, Rebeca Acín-Pérez et al. [15] proposed a plasticity model (Figure 2C) that is more flexible than the solid model or liquid model, with more complex processes for organization of the respiratory chain. Physiological activities suggest the critical roles of SCs in mitochondrial respiratory functions, and support the plasticity model for organization of the mitochondrial respiratory chain. Overall, available evidence suggests the mitochondrial respiratory chain is composed of individual complexes and SCs, both of which participate in electron transfer and have oxidative phosphorylation functions. However, the ratio of free complexes and SCs can be adjusted to adapt internal environment [16].
The assembly of SCs may be regulated by numerous factors, such as mitochondrial cristae morphology, phospholipids, assembly factors, and others. The failure of SC assembly can contribute to the development of various diseases. It has been reported that disorder of SC assembly occurs in various disease models, including neurodegenerative diseases, genetic diseases, heart failure, and cancer [1,19,20,21,22]. Therefore, the assembly and regulation of SCs are crucial for maintaining mitochondrial homeostasis and cellular functions. Here, we review the structure and assembly, the regulation mechanisms, and the functions of SCs, which will provide valuable clues for studying the efficiency of the mitochondrial respiratory chain, regulation mechanisms of cellular metabolism, and of organismal metabolic homeostasis.
2. Structure and Assembly of SCs
SCs can be divided into two categories: those containing CI and those without CI. The three SCs containing CI include CI + CIII, CI + CIII + CIV, and CI + CII + CIII + CIV, while the other two SCs without CI include CIII + CIV, and CII + CIII + CIV [15,16,23] (Table 1). However, SCs are heterogeneous and vary among different species, different tissue structures, and even different physiological states. In yeast cells, almost all the CIV binds to CIII2 to form SCs, and the available CIV determines the number of SCs, while in mammals, almost all the CI is bound in SCs, among which 54% is CICIII2CIV and 17% is CICIII2 [8,24].
There are two hypotheses concerning the assembly of SCs. One model assumes that the SCs are assembled after the individual complexes are formed [15], and another model assumes that the assembly of SCs starts when intermediates of individual complexes are synthesized, followed by successive assembly of the remaining subunits [13,28]. Margherita Protasoni et al. [13] found the accumulation of incompletely assembled CIII2 in a cell line with an MT-CyB deletion mutation. The partial subunit of CIV was found in this incompletely assembled CIII2. These data provided an experimental basis for the “cooperative-assembly model” for assembly as shown in Figure 3. In this model, CIII2, which acts as the central entity for SCs, promotes the sequential assembly of the pre-CI and CIV structural subunit modules with CIII2 [13,28]’ leading to a completely assembled CICIII2CIV. Dynamic assembly of SCs allows the cell to adapt to the internal environment and different carbon sources [10]. In yeast, lactate can induce the biosynthesis of CIV, which, in turn, increase the assembly of CIII2CIV1-2 and enhances OXPHOS [8]. In mammalian cell lines, increased galactose promotes the assembly of SCs to ensure that the ETC works at maximum capacity [10]. Using BN-PAGE, Chiara Greggio et al. [29] found that after 4 months exercise training, the number of CI and the assembly of SCs increased in 26 healthy, sedentary older adults.
2.1. CICIII2CIV
CICIII2CIV consists of a CI, a CIII dimer, and a CIV. The SC is also known as respirasome because it has all the components required to transmit electrons from NADH to oxygen [8,15,30]. Saccharomyces cerevisiae lacks CI, while CIV is almost absent in plants. Therefore, the respirasome is mainly detected in mammalian mitochondria. However, the number of mitochondrial respirasome varies among different species in mammals [31].
In 2016, Jinke Gu et al. [25,32] reported the structure of 1.7 mDa CICIII2IV in porcine heart using cryo-electron microscopy at resolutions up to 5.4 Å and 4.0 Å. The CIII dimer and CIV bind on the membrane arm of the L-shaped CI to form a transmembrane disk [25]. Interactions between the three complexes mainly occur at positions close to the membrane surface, and the interaction between CI and CIII is most obvious [33]. The subunits involved in the interaction between CI and CIII include NDUFB9 (NADH: ubiquinone oxidoreductase subunit B9), NDUFA11 (NADH: ubiquinone oxidoreductase subunit B11), NDUFB8 (NADH: ubiquinone oxidoreductase subunit B8) and NDUFB4 (NADH: ubiquinone oxidoreductase subunit B4) of CI, as well as UQCRC1 (ubiquinol-cytochrome c reductase core protein 1) of CIII [12,25,32]. CI directly interacts with CIII through the NDUFA11 subunit. In addition, the UQCRC1S251-L265, a short loop of UQCRC1, inserts into the N terminus of NDUFB9 subunit to stabilize the interaction of CI with CIII [25,32]. Moreover, the ND5 (NADH dehydrogenase subunit 5), located at the distal end of CI membrane arm, interacts with COX7C (cytochrome c oxidase subunit 7C) of CIV [12,25]. Furthermore, the COX7A subunit of CIV interacts with CIII through UQCRC1 and UQCR11 (ubiquinol-cytochrome c reductase, complex III subunit XI) [12,25,32]. Studies have demonstrated that disruption of the structure of NDUFA11 subunit, or inhibition of NDUFA11 expression can affect CI stability and result in disorder of SC assembly [34,35]. Other studies have suggested that mutations in NDUFB9 abolish CI synthesis [36], disturb NAD+/NADH balance, and promote tumor metastasis [37].
2.2. Other Types of SCs
The structure of CI and CIII2 is conserved in plants, yeast, and mammals. Therefore, the assembly of CICIII2 is similar to that in the respirasome, in which the NDUFB9, NDUFA11, NDUFB8, and NDUFB4 of CI interact with UQCRC1 of CIII to form CICIII2 [26].
CIII and CIV are assembled differently in multiple types of SCs. CIII2CIV1-2 consists of two CIII and one or two CIV. Irene Vercellino et al. [24] revealed the structures of CIII2CIV1-2 in mice by cryo-electron microscopy. Based on the point at which CIV interacted with CIII2, the structure could be divided into two types, a mature unlocked class and locked class. Moreover, the locked structures could be further divided into a locked intermediate class and a locked assembled class. The locked intermediate class contains assembled CIII2 intermediates, while the locked assembled class forms fully assembled CIII2CIV1-2. Unlike the mature unlocked class, the CIV of locked assembled class moves laterally much closer to CIII2. In the conformation of the mature unlocked class, CIV is almost rotated by 180° [24]. The assembly of CIII2CIV1-2 starts by forming the locked intermediate class, then the locked assembled class, and finally forms the mature unlocked class. During the transition, supercomplex assembly factor I (SCAFI) provides an anchor or pivot point for the rotation of CIV and plays a crucial role in stabilizing the structure of SCs [24]. In mice, CIII2CIV1-2 is composed 41% of the mature unlocked class and 59% Iof the locked class, which includes 21% locked intermediate class and 38% locked assembled class. However, in sheep, the unlocked class reaches 93% of the total CIII2CIV1-2, and only 7% is in the locked class [24].
Due to the lack of CI in yeast, the SCs are mainly CII2CIV1-2 and protein interaction is also different than in mammals. The Cor1 subunit of CIII linked to the Cox5a subunit of CIV [27]. Furthermore, Sorbhi Rathore et al. [27] speculated that the surface of these subunits further stabilizes the interactions of CIII2CIV1-2 by binding to cardiolipin molecules by observing the interface densities of Cox5a, Qcr8, Qcr9, and Rip1.
3. Potential Functions of SCs
Numerous studies have proposed several roles of SCs, such as maintaining the integrity of free complexes, improving the efficiency of electron transfer and regulating ROS production. The functions of SCs are shown in Table 2. Each functional advantage is complementary to each other. The loss of one function affects other functional advantages, and ultimately hinders mitochondrial energy metabolism (Figure 4).
3.1. Maintaining the Integrity of Free Complexes
The assembly of SCs is crucial for the assembly and stability of each free complex. Margherita Protasoni et al. [13] found that the number and activity of both CIII2 and CI in SCs significantly decreased in CIII2 subunit MT-CyB-deleted human cell lines, by using BN-GAGE, proteomics and in-gel activity assays. In addition, the subunits of CI and CIV involved in the integration of free complex, were also significantly decreased, and the assembly of CI and CIV was brought to a standstill. Both in Paracoccus denitrificans and the CIII-deleted mouse cell line L929, researchers found that the assembly of SCs was reduced. Moreover, the assembly and activity of CI was decreased, and the stability of assembled CI was reduced due to susceptibility to degradation. In addition to CI, the assembly of CIVs was also inhibited [14,38]. Moreover, Rebeca Acín-Pérez et al. [38] reported that pharmacological inhibition of CIII activity did not affect CI assembly, but physical deletion of CIII resulted in reduced CI assembly. Therefore, CIII is the center for the assembly of SCs, providing a structural and functional platform for the biogenesis of the complete mitochondrial respiratory chain [13].
3.2. Improving the Efficiency of Electron Transfer
The assembly of SCs facilitates the diffusion of ubiquinone and cytochrome c by shortening the distance between the complexes, and by increased electron transfer efficiency.
Cristina Bianchi et al. [11] proposed a model in which the existence of electron channels between mitochondrial respiratory chain complexes, and specific ubiquinone and cytochrome c could spread and transfer electrons, and the mitochondrial respiratory chain complex could control electron flux. Although the substrate channel theory is still controversial, growing evidence suggests that ubiquinone and cytochrome c are free to diffuse between the complexes [1]. James A. Letts et al. [12] reported that the active sites of each complex opened to the inner mitochondrial membrane at an interval of 10 nm in sheep mitochondria, detected by cryo-electron microscopy at a resolution of 5.8 Å. Although no protein-mediated substrate channels connect the individual SCs, ubiquinone and cytochrome can flow freely between them for electron transfer. These structural arrangements not only shorten the diffusion of the transmitters, but also reduce electron leakage [12]. Jens Berndtsson et al. [40] found that the efficiency of electron transfer was reduced in yeast when the interaction between CIII and CIV was disrupted by altering the COR1 subunit. Introducing endogenous or exogenous cytochrome c significantly increased the growth of yeast. Agnes Moe et al. [39] also reported that cytochrome c with a positive charge can slide along the negatively charged surface of CIII2CIV in yeast cells. These results suggest that SCs may shorten the diffusion distance of cytochrome c to enhance electron transfer efficiency.
Studies have demonstrated that the enzyme activities of CIII2 and CIV in SCs are 1.7 and 1.9 times higher than those in free complexes in mammals, resulting in stronger catalytic activity for the redox reactions of cytochrome c and higher efficiency for electron transfer [24]. Compared to free CIII and CIV, CIII2CIV improved efficiency of mitochondrial respiration and ATP synthesis [16].
3.3. Regulating ROS Production
Mitochondria-derived ROS are mainly produced by CI and CIII, which may due to decreased electron flux between complexes [41,42]. The assembly of SCs can reduce ROS production by increasing electron transfer efficiency [11,12].
Irene Lopez-Fabuela et al. found that ROS production in astrocytes was higher than that in neurons. Further analysis showed that mitochondrial complex I is predominantly assembled into supercomplexes in neurons, whereas abundant free complex I can be detected in astrocytes. In CI over-expressed and knockdown models, it has been demonstrated that assembly of SCs can decrease ROS production [43]. In the respirasome, UQCRFS1, the distal subunit of CIII, COX6a2, and the subunit of CIV, were tightly associated to prevent the oxidation of ubiquinone and reduce ROS production in CIII [12,33]. Moreover, disrupting the structure of SCs increased the production of ROS. Evelina Maranzana et al. [41] found that dissociating CI from SCs by dodecyl maltoside (DDM) increased ROS production. In yeast cells that lacked regulatory SCs assembly factors, elevated ROS could be detected [44].
4. Regulation of SCs
Many studies have investigated the structure of SCs, and have found several factors regulating SC assembly. The regulation of SCs is shown in Table 3.
4.1. SCAFI
The cytochrome c oxidase subunit 7A-related protein (COX7a2L), also named SCAFI, plays critical roles in regulating the assembly of CIII and CIV. However, SCAFI is only present in SCs containing CIII2CIV, but not in free CIII and CIV [16]. SCAFI has two phenotypes, a long type containing 113 amino acids, and a short type encoding 111 amino acids. The phenotype of SCAFI varies among species and strains. For example, the phenotype is different between C57/6J and Balb/CJ mice [16,17]. The long-type SCAFI in mammals prompts the assembly of CIII2 and CIV into SCs, whereas the short-type SCAFI results in deficiency of CIII2CIV in strains [16,23].
SCAFI, a homolog of the COX7a subunit of CIV [12], has a similar amino acid sequence to COX7a. However, SCAFI contains an extra domain that can bind to CIII [23]. Mouse CIII2CIV was analyzed by cryo-electron microscopy and shows that SCAFI forms a physical linkage between both complexes [24]. Studies have revealed that the N terminus of SCAFI interacts with the N terminus of sub9 by electrostatic effects and is embedded in the cavity of CIII2. SCAFI also binds to CIV via the C terminus of the transmembrane helix at the COX7a subunit. Therefore, COX7a is replaced by SCAFI during the assembly of SCs containing CIII and CIV, and SCAFI bridges CIII and CIV to form CIII2CIV [12,16,23,24]. SCAFI is not only an assembly factor, but also participates in the interaction between CIII and CIV to stabilize SCs [24].
Although CIII2CIV does not assemble in mice because mice congenitally express short-type SCAFI, a certain number of respirasomes and free complexes can be detected in the hearts of C57 mice. No severe abnormalities of respiratory chain functions have been observed in mice [2,17,23]. In a SCAFI-deficient zebrafish model, the interaction between CIII and CIV was disrupted, which caused a significant reduction in CIII2CIV, as well as respirasomes. These changes resulted in a substantial impairment of respiratory functions, and finally led to a significant reduction in body size and fertility [45]. It is assumed that congenital deletion of SCAFI in mice regulates the type and ratio of SCs to compensate for the deficiency of CIII2CIV. However, gene silencing of SCAFI may cause serious disorder of mitochondrial respiratory functions due to limited compensatory capacity. Further analysis has shown that SCAFI was absent in both CICIII2 and the respirasome, suggesting that SCAFI is an CIII2CIV specific assembly factor [24].
4.2. Respiratory Supercomplex Factor 1
Respiratory supercomplex factor 1 (Rcf1) and respiratory supercomplex factor 2 (Rcf2) belong to the hypoxia inducible gene domain (HIGD) family. Rcf2 is expressed in yeast cells, while Rcf1 and its homologs are expressed in the plants and mammals.
In yeast cells, both Rcf1 and Rcf2 are subunits that made up the CIV and SCs. Rcf1 is necessary to maintain cellular respiration, and affects the assembly of CIV and SCs by regulating cytochrome c oxidase subunit VIa (COX13) [46]. Moreover, Rcf1 also directly interacts with CIII and CIV to form CIII2CIV [44]. Rcf1 knockout results in a significant decrease of CIII2CIV and an obvious increase of free CIII2 and CIV, which increases ROS production and elevates oxidative stress in yeast cells [44,46]. In an Rcf1-overexpressed model, both CIII2CIV and CIII2CIV2 are significantly increased while free CIV is absent, suggesting that elevated Rcf1 expression can promote the formation of SCs [44].
HIGD1A and HIGD2A, two homologs of Rcf1 in mammals [49], are two important subunits of CIV that play similar roles as Rcf1 [44,46,50]. It was initially reported that HIG1A could be strongly induced by hypoxia in an HIF-1-dependent manner. Although HIGD1A is not essential for the basic functions of mitochondria in mammals, it is critical for enhancement of electron transport and improving respiratory efficiency. Studies also showed that HIG2A knockdown led to depletion of CIV-containing SCs, especially CICIICIIICIV, CICIII2CIV, and CIII2CIV1-2 [44,46]
4.3. Other Regulatory Factors
In addition to the above-mentioned factors that regulate the assembly of SCs, other regulatory networks containing phospholipids and cytochrome c, have also been identified. Several studies found large amounts of phospholipids, such as cardiolipin (CL), phosphatidylethanolamine (PC), and phosphatidylcholine (PE), in mitochondrial SCs [30,47,51]. Cryo-electron microscopy with high-resolution has revealed that lipid molecules affect the enzyme complex catalytic activity by participating in interactions between complexes in SCs. For example, CL, PC, and PE interact with NDUFA11 [32,51,52]. Moreover, CL is present in the inner mitochondrial membrane and plays important roles in stabilizing the structure of SCs [53,54,55]. Knockdown of the Crd1 gene that encodes cardiolipin synthase, impairs CL synthesis, reduces assembly of CIII2CIV2 complexes, and increases free CIII2 and CIV2 [47]. Moreover, a small number of assembled SCs are not as stable as those in wild-type cell lines [9,30].
Cytochrome c is not only a transmitter of electrons in the mitochondrial respiratory chain, but a pivotal molecule in the apoptosis-associated signaling pathway. It is well-known that cytochrome c is required for CIV assembly [48]. Uma D. Vempati et al. [53] found that knockdown of cytochrome c gene completely abolished CIV activity, decreased CIII activity to 50% of the normal level, and significantly reduced the activity of cardiolipin synthase, which in turn affected assembly of SCs.
5. SCs in the Development of Diseases
Disorder of the assembly of SCs, due to gene mutation or environmental stimulation, had been widely reported in various diseases. The decrease or absence of SCs has emerged as a possible secondary cause of these diseases (Table 4). Failure of SC assembly might result in weak adaptability to environmental changes, due to impaired mitochondrial energy metabolism.
Alzheimer’s and Parkinson’s disease are two major neurodegenerative diseases. Obvious amyloid deposition, mitochondrial dysfunction, and oxidative stress can be observed in brain tissue of AD and PD patients [21]. Amyloid accumulation alters phospholipids of the mitochondrial inner membrane, such as increasing ceramide, which results in dysfunction of mitochondrial energy metabolism by affecting assembly and function of SCs [22]. Studies have also found that the SCs are lower in the cerebral cortex of aging rats than that in young rats [57]. It had been demonstrated that functional deficiencies of PINK1 and DJ-1 are closely associated with PD. PINK1 knockout impaired mitochondrial division, decreased the enzymatic activity of CI and CIV, as well as reduced the assembly of SCs [21,58], while deletion of DJ-1 in dopaminergic neurons decreased the assembly and activity of CI in mitochondria [21].
Barth syndrome is an X-linked recessive inheritable disease mainly caused by mutations in the Tafazzin (TAZ) gene, which leads to disorder of cardiolipin remodeling [59]. In mouse model of Barth syndrome, Tafazzin knockout by doxycycline significantly impaired cardiolipin metabolism, reduced SCs and increased free CI. Moreover, it was reported that Tafazzin knockout resulted in disruption of mitochondrial structure, and mitochondrial strings and vesicular mitochondrial cristae were observed [19]. Leigh syndrome (LS), also known as subacute necrotizing encephalomyelopathy (SNEM), is a rare mitochondrial disease caused by a genetic deficiency [60]. Fabian Baertling et al. [20] found impaired CI synthesis in fibroblasts from NDUFAF4-mutated Leigh syndrome patients that decreased the assembly of CICIII2 and CICIII2CIV.
6. Conclusions and Future Perspectives
With the development of various advanced technologies, such as BN-PAGE, 2D BN-PAGE and cryo-electron microscopy, the location of SCs in the inner mitochondrial membrane has been demonstrated, and researchers have focused on exploring the structure and assembly of SCs [15,25,32,33]. The plasticity model of respiratory chain has been attracting more and more attention [2,3]. However, more experimental evidence is required to support the plasticity model, and the processes of assembly should be further investigated. There are several limitations to study of the assembly of SCs. First, the detergents used in isolation and purification of SCs inevitably impact the components of SCs. Therefore, isolated SCs are always different from intact SCs. Second, the varied methods of sample preparation used for cryo-electron microscopy techniques may affect results [21]. Therefore, it is necessary to optimize current technologies or develop novel methods for uncovering the assembly and functions of SCs.
Moreover, the functions of SCs, such as reducing ROS production and increasing electron transfer efficiency through substrate channels, still need to be verified in further in vitro and in vivo models [1]. It also has been widely demonstrated that abnormalities of SCs participate in the pathological processes of various diseases, especially neurodegenerative diseases, and impaired assembly of SCs has been emerged as a major cause. However, the potential mechanisms still need to be further investigated.
Author Contributions
S.G. wrote the first draft. L.Z. and R.P. edited the draft. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the (National Natural Science Foundation of China) under Grant (number 82073511).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare that they have no competing interest.
References
- Vercellino, I.; Sazanov, L.A. The assembly, regulation and function of the mitochondrial respiratory chain. Nat. Rev. Mol. Cell Biol. 2022, 23, 141–161. [Google Scholar] [CrossRef] [PubMed]
- Enríquez, J.A. Supramolecular Organization of Respiratory Complexes. Annu. Rev. Physiol. 2016, 78, 533–561. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Loshuertos, R.; Enríquez, J.A. Respiratory supercomplexes and the functional segmentation of the CoQ pool. Free Radic. Biol. Med. 2016, 100, 5–13. [Google Scholar] [CrossRef]
- Green, D.E.; Tzagoloff, A. The mitochondrial electron transfer chain. Arch. Biochem. Biophys. 1966, 116, 293–304. [Google Scholar] [CrossRef]
- Hatefi, Y.; Haavik, A.G.; Fowler, L.R.; Griffiths, D.E. Studies on the electron transfer system. XLII. Reconstitution of the electron transfer system. J. Biol. Chem. 1962, 237, 2661–2669. [Google Scholar] [CrossRef]
- Hackenbrock, C.R. Molecular Organization and the Fluid Nature of the Mitochondrial Energy Transducing Membrane. Struct. Biol. Membr. 1977, 34, 199–234. [Google Scholar]
- Höchli, M.; Hackenbrock, C.R. Fluidity in mitochondrial membranes: Thermotropic lateral translational motion of intramembrane particles. Proc. Natl. Acad. Sci. USA 1976, 73, 1636–1640. [Google Scholar] [CrossRef] [Green Version]
- Schägger, H.; Pfeiffer, K. Supercomplexes in the respiratory chains of yeast and mammalian mitochondria. EMBO J. 2000, 19, 1777–1783. [Google Scholar] [CrossRef] [Green Version]
- Cogliati, S.; Frezza, C.; Soriano, M.E.; Varanita, T.; Quintana-Cabrera, R.; Corrado, M.; Cipolat, S.; Costa, V.; Casarin, A.; Gomes, L.C.; et al. Mitochondrial Cristae Shape Determines Respiratory Chain Supercomplexes Assembly and Respiratory Efficiency. Cell 2013, 155, 160–171. [Google Scholar] [CrossRef] [Green Version]
- Acin-Perez, R.; Enriquez, J.A. The function of the respiratory supercomplexes: The plasticity model. Biochim. Biophys. Acta 2014, 1837, 444–450. [Google Scholar] [CrossRef] [Green Version]
- Bianchi, C.; Genova, M.L.; Castelli, G.P.; Lenaz, G. The mitochondrial respiratory chain is partially organized in a supercomplex assembly: Kinetic evidence using flux control analysis. J. Biol. Chem. 2004, 279, 36562–36569. [Google Scholar] [CrossRef] [PubMed]
- Letts, J.A.; Fiedorczuk, K.; Sazanov, L.A. The architecture of respiratory supercomplexes. Nature 2016, 537, 644. [Google Scholar] [CrossRef]
- Protasoni, M.; Pérez-Pérez, R.; Lobo-Jarne, T.; Harbour, M.E.; Ding, S.; Peñas, A.; Diaz, F.; Moraes, C.T.; Fearnley, I.M.; Zeviani, M.; et al. Respiratory supercomplexes act as a platform for complex III-mediated maturation of human mitochondrial complexes I and IV. EMBO J. 2020, 39, e102817. [Google Scholar] [CrossRef] [PubMed]
- Stroh, A.; Anderka, O.; Pfeiffer, K.; Yagi, T.; Finel, M.; Ludwig, B.; Schägger, H. Assembly of respiratory complexes I, III, and IV into NADH oxidase supercomplex stabilizes complex I in Paracoccus denitrificans. J. Biol. Chem. 2004, 279, 5000–5007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acin-Perez, R.; Fernandez-Silva, P.; Peleato, M.L.; Perez-Martos, A.; Enriquez, J.A. Respiratory Active Mitochondrial Supercomplexes. Mol. Cell 2008, 32, 529–539. [Google Scholar] [CrossRef]
- Lapuente-Brun, E.; Moreno-Loshuertos, R.; Acin-Perez, R.; Latorre-Pellicer, A.; Colas, C.; Balsa, E.; Perales-Clemente, E.; Quiros, P.M.; Calvo, E.; Rodriguez-Hernandez, M.A.; et al. Supercomplex Assembly Determines Electron Flux in the Mitochondrial Electron Transport Chain. Science 2013, 340, 1567–1570. [Google Scholar] [CrossRef]
- Mourier, A.; Matic, S.; Ruzzenente, B.; Larsson, N.G.; Milenkovic, D. The respiratory chain supercomplex organization is independent of COX7a2l isoforms. Cell Metab. 2014, 20, 1069–1075. [Google Scholar] [CrossRef] [Green Version]
- Wittig, I.; Braun, H.; Schagger, H. Blue Native PAGE; Nature Publishing Group: Berlin, Germany, 2006. [Google Scholar]
- Kim, J.; Lee, K.; Fujioka, H.; Tandler, B.; Hoppel, C.L. Cardiac mitochondrial structure and function in tafazzin-knockdown mice. Mitochondrion 2018, 43, 53–62. [Google Scholar] [CrossRef]
- Baertling, F.; Sánchez-Caballero, L.; van den Brand, M.A.M.; Wintjes, L.T.; Brink, M.; van den Brandt, F.A.; Wilson, C.; Rodenburg, R.J.T.; Nijtmans, L.G.J. NDUFAF4 variants are associated with Leigh syndrome and cause a specific mitochondrial complex I assembly defect. Eur. J. Hum. Genet. 2017, 25, 1273–1277. [Google Scholar] [CrossRef]
- Novack, G.V.; Galeano, P.; Castaño, E.M.; Morelli, L. Mitochondrial Supercomplexes: Physiological Organization and Dysregulation in Age-Related Neurodegenerative Disorders. Front. Endocrinol. 2020, 11, 600. [Google Scholar] [CrossRef]
- Pera, M.; Larrea, D.; Guardia-Laguarta, C.; Montesinos, J.; Velasco, K.R.; Agrawal, R.R.; Xu, Y.; Chan, R.B.; Di Paolo, G.; Mehler, M.F.; et al. Increased localization of APP-C99 in mitochondria-associated ER membranes causes mitochondrial dysfunction in Alzheimer disease. EMBO J. 2017, 36, 3356–3371. [Google Scholar] [CrossRef] [PubMed]
- Cogliati, S.; Calvo, E.; Loureiro, M.; Guaras, A.M.; Nieto-Arellano, R.; Garcia-Poyatos, C.; Ezkurdia, I.; Mercader, N.; Vázquez, J.; Enriquez, J.A. Mechanism of super-assembly of respiratory complexes III and IV. Nature 2016, 539, 579–582. [Google Scholar] [CrossRef] [PubMed]
- Vercellino, I.; Sazanov, L.A. Structure and assembly of the mammalian mitochondrial supercomplex CIII2CIV. Nature 2021, 598, 364. [Google Scholar] [CrossRef]
- Gu, J.; Wu, M.; Guo, R.; Yan, K.; Lei, J.; Gao, N.; Yang, M. The architecture of the mammalian respirasome. Nature 2016, 537, 639. [Google Scholar] [CrossRef] [PubMed]
- Davies, K.M.; Blum, T.B.; Kühlbrandt, W. Conserved in situ arrangement of complex I and III2 in mitochondrial respiratory chain supercomplexes of mammals, yeast, and plants. Proc. Natl. Acad. Sci. USA 2018, 115, 3024–3029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rathore, S.; Berndtsson, J.; Marin-Buera, L.; Conrad, J.; Carroni, M.; Brzezinski, P.; Ott, M. Cryo-EM structure of the yeast respiratory supercomplex. Nat. Struct. Mol. Biol. 2019, 26, 50–57. [Google Scholar] [CrossRef]
- Moreno-Lastres, D.; Fontanesi, F.; García-Consuegra, I.; Martín, M.A.; Arenas, J.; Barrientos, A.; Ugalde, C. Mitochondrial complex I plays an essential role in human respirasome assembly. Cell Metab. 2012, 15, 324–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greggio, C.; Jha, P.; Kulkarni, S.S.; Lagarrigue, S.; Broskey, N.T.; Boutant, M.; Wang, X.; Conde Alonso, S.; Ofori, E.; Auwerx, J.; et al. Enhanced Respiratory Chain Supercomplex Formation in Response to Exercise in Human Skeletal Muscle. Cell Metab. 2017, 25, 301–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shinzawa-Itoh, K.; Shimomura, H.; Yanagisawa, S.; Shimada, S.; Takahashi, R.; Oosaki, M.; Ogura, T.; Tsukihara, T. Purification of Active Respiratory Supercomplex from Bovine Heart Mitochondria Enables Functional Studies. J. Biol. Chem. 2016, 291, 4178–4184. [Google Scholar] [CrossRef] [Green Version]
- Protasoni, M.; Zeviani, M. Mitochondrial Structure and Bioenergetics in Normal and Disease Conditions. Int. J. Mol. Sci. 2021, 22, 586. [Google Scholar] [CrossRef]
- Wu, M.; Gu, J.; Guo, R.; Huang, Y.; Yang, M. Structure of Mammalian Respiratory Supercomplex I1III2IV1. Cell 2016, 167, 1598. [Google Scholar] [CrossRef] [PubMed]
- Sousa, J.S.; Mills, D.J.; Vonck, J.; Kühlbrandt, W. Functional asymmetry and electron flow in the bovine respirasome. Elife 2016, 5, e21290. [Google Scholar] [CrossRef] [PubMed]
- Berger, I.; Hershkovitz, E.; Shaag, A.; Edvardson, S.; Saada, A.; Elpeleg, O. Mitochondrial complex I deficiency caused by a deleterious NDUFA11 mutation. Ann. Neurol. 2008, 63, 405–408. [Google Scholar] [CrossRef] [PubMed]
- Andrews, B.; Carroll, J.; Ding, S.; Fearnley, I.M.; Walker, J.E. Assembly factors for the membrane arm of human complex I. Proc. Natl. Acad. Sci. USA 2013, 110, 18934–18939. [Google Scholar] [CrossRef] [Green Version]
- Haack, T.B.; Madignier, F.; Herzer, M.; Lamantea, E.; Danhauser, K.; Invernizzi, F.; Koch, J.; Freitag, M.; Drost, R.; Hillier, I.; et al. Mutation screening of 75 candidate genes in 152 complex I deficiency cases identifies pathogenic variants in 16 genes including NDUFB9. J. Med. Genet. 2012, 49, 83–89. [Google Scholar] [CrossRef] [Green Version]
- Tan, A.S.; Baty, J.W.; Berridge, M.V. The role of mitochondrial electron transport in tumorigenesis and metastasis. Biochim. Biophys. Acta 2014, 1840, 1454–1463. [Google Scholar] [CrossRef]
- Acín-Pérez, R.; Bayona-Bafaluy, M.P.; Fernández-Silva, P.; Moreno-Loshuertos, R.; Pérez-Martos, A.; Bruno, C.; Moraes, C.T.; Enríquez, J.A. Respiratory complex III is required to maintain complex I in mammalian mitochondria. Mol. Cell 2004, 13, 805–815. [Google Scholar] [CrossRef]
- Moe, A.; Di Trani, J.; Rubinstein, J.L.; Brzezinski, P. Cryo-EM structure and kinetics reveal electron transfer by 2D diffusion of cytochrome c in the yeast III–IV respiratory supercomplex. Proc. Natl. Acad. Sci. USA 2021, 118, e2021157118. [Google Scholar] [CrossRef]
- Berndtsson, J.; Aufschnaiter, A.; Rathore, S.; Marin-Buera, L.; Dawitz, H.; Diessl, J.; Kohler, V.; Barrientos, A.; Büttner, S.; Fontanesi, F.; et al. Respiratory supercomplexes enhance electron transport by decreasing cytochrome c diffusion distance. EMBO Rep. 2020, 21, e51015. [Google Scholar] [CrossRef]
- Maranzana, E.; Barbero, G.; Falasca, A.I.; Lenaz, G.; Genova, M.L. Mitochondrial respiratory supercomplex association limits production of reactive oxygen species from complex I. Antioxid. Redox Signal. 2013, 19, 1469–1480. [Google Scholar] [CrossRef] [Green Version]
- Lenaz, G. Mitochondria and reactive oxygen species. Which role in physiology and pathology? Adv. Exp. Med. Biol. 2012, 942, 93–136. [Google Scholar] [PubMed]
- Lopez-Fabuel, I.; Le Douce, J.; Logan, A.; James, A.M.; Bonvento, G.; Murphy, M.P.; Almeida, A.; Bolaños, J.P. Complex I assembly into supercomplexes determines differential mitochondrial ROS production in neurons and astrocytes. Proc. Natl. Acad. Sci. USA 2016, 113, 13063–13068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.C.; Taylor, E.B.; Dephoure, N.; Heo, J.M.; Tonhato, A.; Papandreou, I.; Nath, N.; Denko, N.C.; Gygi, S.P.; Rutter, J. Identification of a protein mediating respiratory supercomplex stability. Cell Metab. 2012, 15, 348–360. [Google Scholar] [CrossRef] [Green Version]
- García-Poyatos, C.; Cogliati, S.; Calvo, E.; Hernansanz-Agustín, P.; Lagarrigue, S.; Magni, R.; Botos, M.; Langa, X.; Amati, F.; Vázquez, J.; et al. Scaf1 promotes respiratory supercomplexes and metabolic efficiency in zebrafish. EMBO Rep. 2020, 21, e50287. [Google Scholar] [CrossRef]
- Vukotic, M.; Oeljeklaus, S.; Wiese, S.; Voegtle, F.N.; Meisinger, C.; Meyer, H.E.; Zieseniss, A.; Katschinski, D.M.; Jans, D.C.; Jakobs, S.; et al. Rcf1 Mediates Cytochrome Oxidase Assembly and Respirasome Formation, Revealing Heterogeneity of the Enzyme Complex. Cell Metab. 2012, 15, 336–347. [Google Scholar] [CrossRef] [Green Version]
- Pfeiffer, K.; Gohil, V.; Stuart, R.A.; Hunte, C.; Brandt, U.; Greenberg, M.L.; Schägger, H. Cardiolipin stabilizes respiratory chain supercomplexes. J. Biol. Chem. 2003, 278, 52873–52880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrientos, A.; Pierre, D.; Lee, J.; Tzagoloff, A. Cytochrome oxidase assembly does not require catalytically active cytochrome C. J. Biol. Chem. 2003, 278, 8881–8887. [Google Scholar] [CrossRef] [Green Version]
- Strogolova, V.; Furness, A.; Robb-McGrath, M.; Garlich, J.; Stuart, R.A. Rcf1 and Rcf2, members of the hypoxia-induced gene 1 protein family, are critical components of the mitochondrial cytochrome bc1-cytochrome c oxidase supercomplex. Mol. Cell. Biol. 2012, 32, 1363–1373. [Google Scholar] [CrossRef] [Green Version]
- Timón-Gómez, A.; Bartley-Dier, E.L.; Fontanesi, F.; Barrientos, A. HIGD-Driven Regulation of Cytochrome c Oxidase Biogenesis and Function. Cells 2020, 9, 2620. [Google Scholar] [CrossRef]
- Sharpley, M.S.; Shannon, R.J.; Draghi, F.; Hirst, J. Interactions between phospholipids and NADH: Ubiquinone oxidoreductase (complex I) from bovine mitochondria. Biochemistry 2006, 45, 241–248. [Google Scholar] [CrossRef]
- Guo, R.; Gu, J.; Zong, S.; Wu, M.; Yang, M. Structure and mechanism of mitochondrial electron transport chain. Biomed. J. 2018, 41, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Vempati, U.D.; Han, X.; Moraes, C.T. Lack of cytochrome c in mouse fibroblasts disrupts assembly/stability of respiratory complexes I and IV. J. Biol. Chem. 2009, 284, 4383–4391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, B.H. Human cytochrome oxidase deficiency. Pediatr. Res. 2000, 48, 581–585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lobo-Jarne, T.; Ugalde, C. R1-2018-Respiratory chain supercomplexes: Structures, function and biogenesis. Semin. Cell Dev. Biol. 2018, 76, 179–190. [Google Scholar] [CrossRef]
- Heo, J.Y.; Park, J.H.; Kim, S.J.; Seo, K.S.; Han, J.S.; Lee, S.H.; Kim, J.M.; Park, J.I.; Park, S.K.; Lim, K.; et al. DJ-1 null dopaminergic neuronal cells exhibit defects in mitochondrial function and structure: Involvement of mitochondrial complex I assembly. PLoS ONE 2012, 7, e32629. [Google Scholar] [CrossRef]
- Frenzel, M.; Rommelspacher, H.; Sugawa, M.D.; Dencher, N.A. Ageing alters the supramolecular architecture of OxPhos complexes in rat brain cortex. Exp. Gerontol. 2010, 45, 563–572. [Google Scholar] [CrossRef]
- Liu, W.; Acín-Peréz, R.; Geghman, K.D.; Manfredi, G.; Lu, B.; Li, C. Pink1 regulates the oxidative phosphorylation machinery via mitochondrial fission. Proc. Natl. Acad. Sci. USA 2011, 108, 12920–12924. [Google Scholar] [CrossRef]
- Ren, H.; Wang, W.; Lin, Y.; Lou, J.; Ji, K.; Zhao, Y.; Yan, C. A case report of Barth syndrome. J. Shandong Univ. (Health Sci.) 2022, 60, 123–127. [Google Scholar]
- Zhao, H.; Wang, X.; Liang, Y.; Liu, H.; Wu, C. A case Child Leigh Syndrome. Chin. J. Lab. Diagn. 2019, 23, 889–892. [Google Scholar]
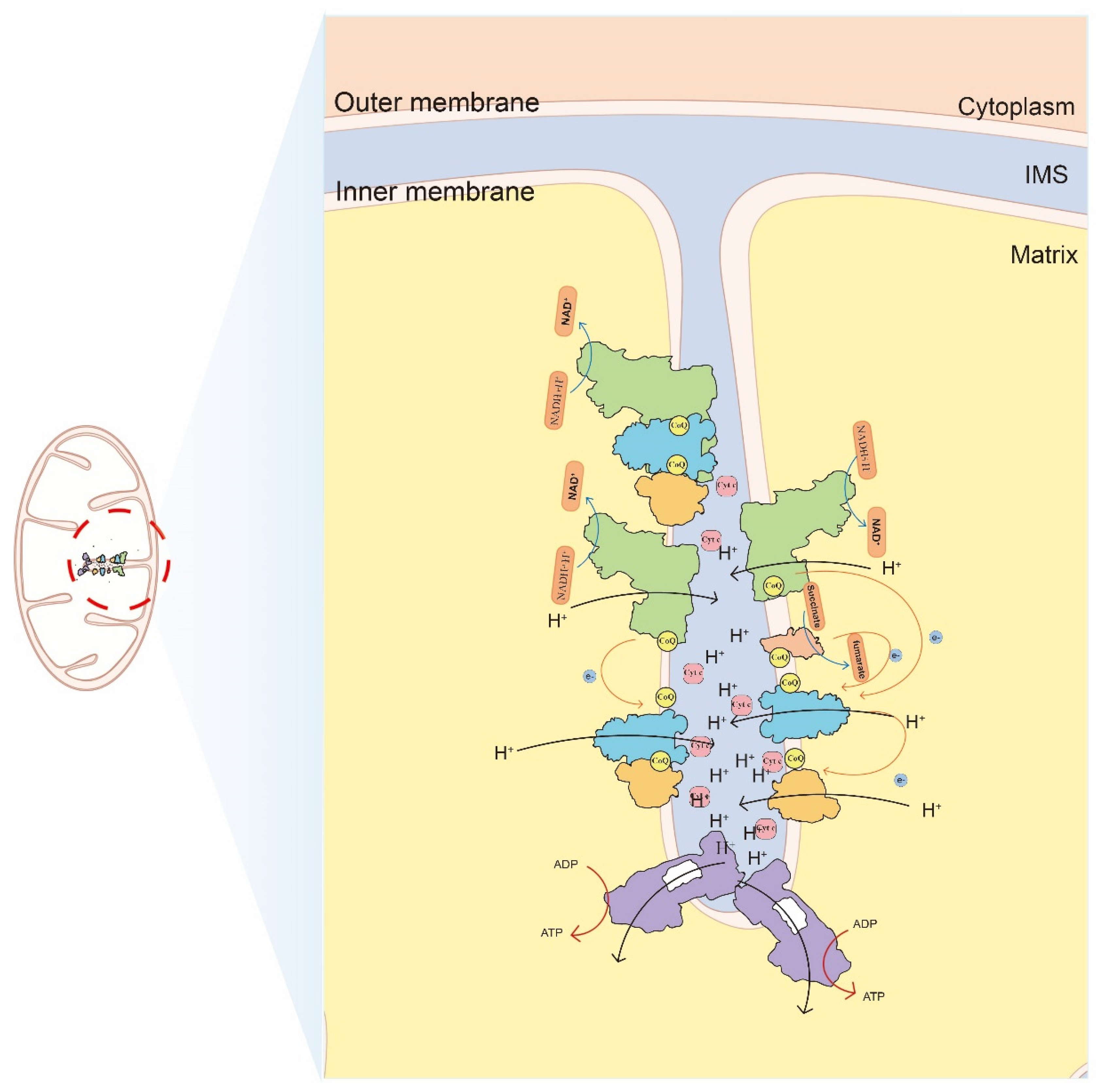
Figure 1.
Mitochondrial cristae and the oxidative phosphorylation located on cristae. The free complexes and SCs transport electron to others though two mobile electron carriers and generate ATP. The position of cytoplasm, matrix, intermembrane space (IMS), cristae and inner membrane are indicated. The complexes are distinguished by different colors: CI: green, CII: pink, CIII dimer: blue, CIV: orange, CV: purple.
Figure 1.
Mitochondrial cristae and the oxidative phosphorylation located on cristae. The free complexes and SCs transport electron to others though two mobile electron carriers and generate ATP. The position of cytoplasm, matrix, intermembrane space (IMS), cristae and inner membrane are indicated. The complexes are distinguished by different colors: CI: green, CII: pink, CIII dimer: blue, CIV: orange, CV: purple.

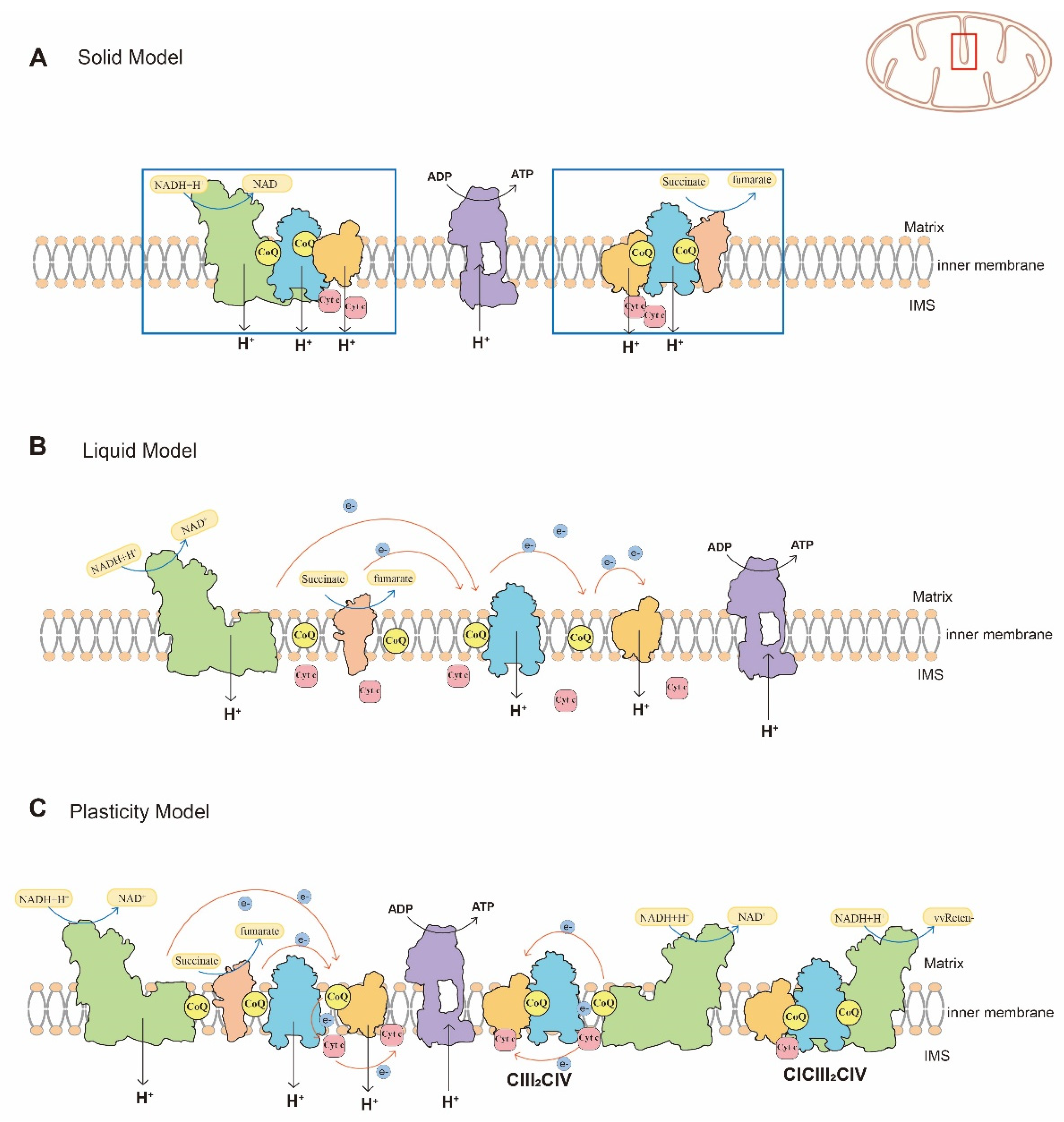
Figure 2.
Schematic representation of the different assembly models of the mitochondrial respiratory chain. (A) The solid model of mitochondrial respiratory chain. The mitochondrial respiratory chain complexes and electron carriers are anchored within a framework and interact with others tightly to ensure high catalytic activity. (B) The liquid model of mitochondrial respiratory chain. Respiratory chain complexes are freely located on the inner mitochondrial membrane and are connected by freely diffusible electron carriers. (C) The plasticity model of the mitochondrial respiratory chain. In this model, individual complexes and SCs participate in electron transfer collectively or individually to adapt to the internal environment. The position of cytoplasm, matrix, the intermembrane space (IMS), cristae and the inner membrane are indicated. Green = CI, pink = CII, blue = CIII dimer, orange = CIV, purple = CV.
Figure 2.
Schematic representation of the different assembly models of the mitochondrial respiratory chain. (A) The solid model of mitochondrial respiratory chain. The mitochondrial respiratory chain complexes and electron carriers are anchored within a framework and interact with others tightly to ensure high catalytic activity. (B) The liquid model of mitochondrial respiratory chain. Respiratory chain complexes are freely located on the inner mitochondrial membrane and are connected by freely diffusible electron carriers. (C) The plasticity model of the mitochondrial respiratory chain. In this model, individual complexes and SCs participate in electron transfer collectively or individually to adapt to the internal environment. The position of cytoplasm, matrix, the intermembrane space (IMS), cristae and the inner membrane are indicated. Green = CI, pink = CII, blue = CIII dimer, orange = CIV, purple = CV.

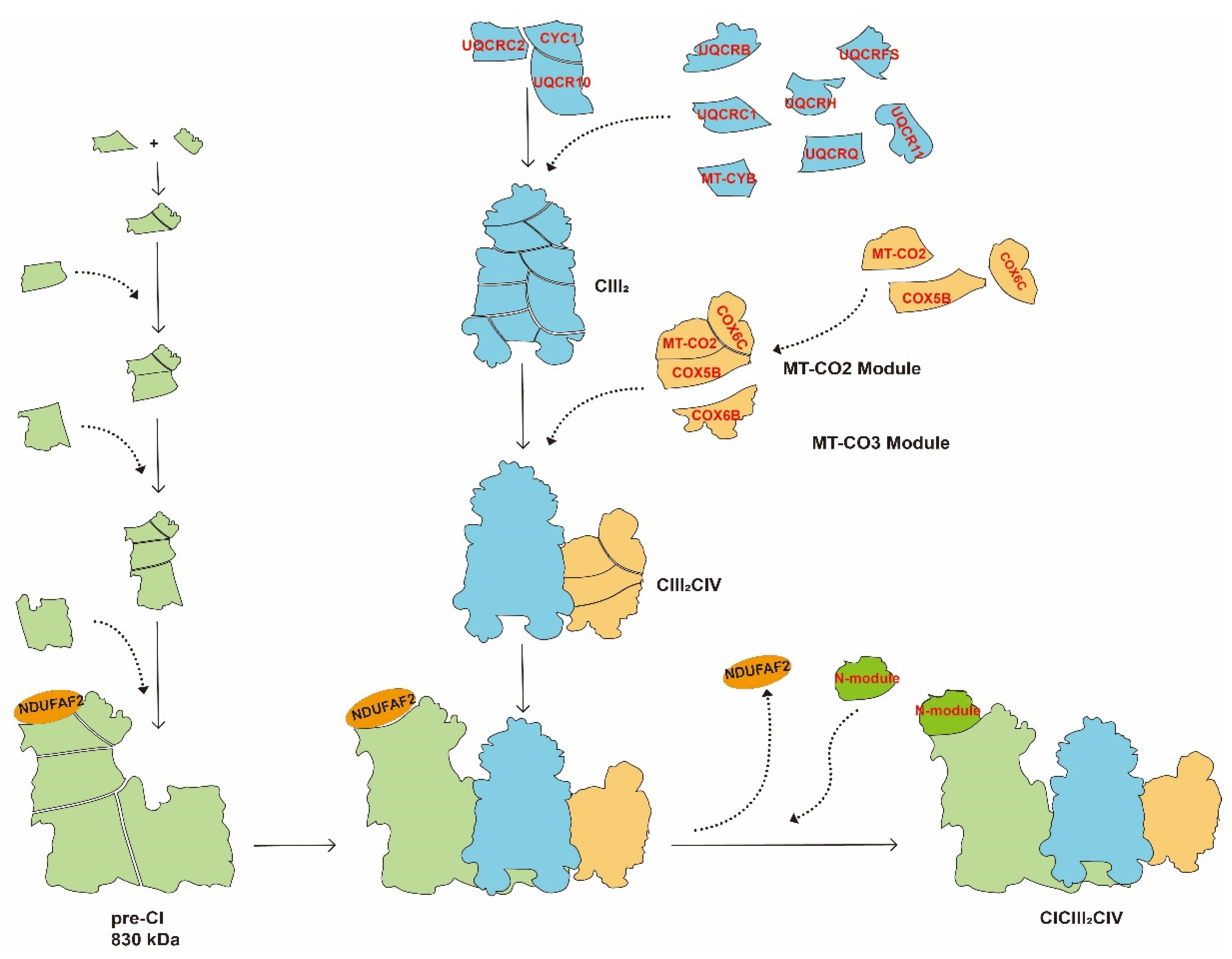
Figure 3.
Assembly of CICIII2CIV by the cooperative-assembly model. CYC1, QUCRC2, and UQCRC10 are assembled to the pre-CIII2 stage and then other subunits are assembled successively to generate CIII2, which acts as the central entity for SCs. At the early stage of assembly of CIII2, CI builds up as a pre-CI of ~830 kDa in which the binding site of the N-module subunit is occupied by NDUFA12 to stabilize the pre-CI. In addition, several CIV subunits belonging to the MT-CO2 module (MT-CO2, COX5B, and COX6C) and the MT-CO3 module (COX6B1) interact with CYC1 and UQCR10. Then, CIII2 binds with pre-CI. Finally, NDUFAF2 is released from pre-CI, and the N-module is assembled to form the complete CICIII2CIV. The SCs are distinguished by different colors: CICIII2CIV: green, blue, and orange; CIII2CIV: blue and orange; CIII2: blue.
Figure 3.
Assembly of CICIII2CIV by the cooperative-assembly model. CYC1, QUCRC2, and UQCRC10 are assembled to the pre-CIII2 stage and then other subunits are assembled successively to generate CIII2, which acts as the central entity for SCs. At the early stage of assembly of CIII2, CI builds up as a pre-CI of ~830 kDa in which the binding site of the N-module subunit is occupied by NDUFA12 to stabilize the pre-CI. In addition, several CIV subunits belonging to the MT-CO2 module (MT-CO2, COX5B, and COX6C) and the MT-CO3 module (COX6B1) interact with CYC1 and UQCR10. Then, CIII2 binds with pre-CI. Finally, NDUFAF2 is released from pre-CI, and the N-module is assembled to form the complete CICIII2CIV. The SCs are distinguished by different colors: CICIII2CIV: green, blue, and orange; CIII2CIV: blue and orange; CIII2: blue.

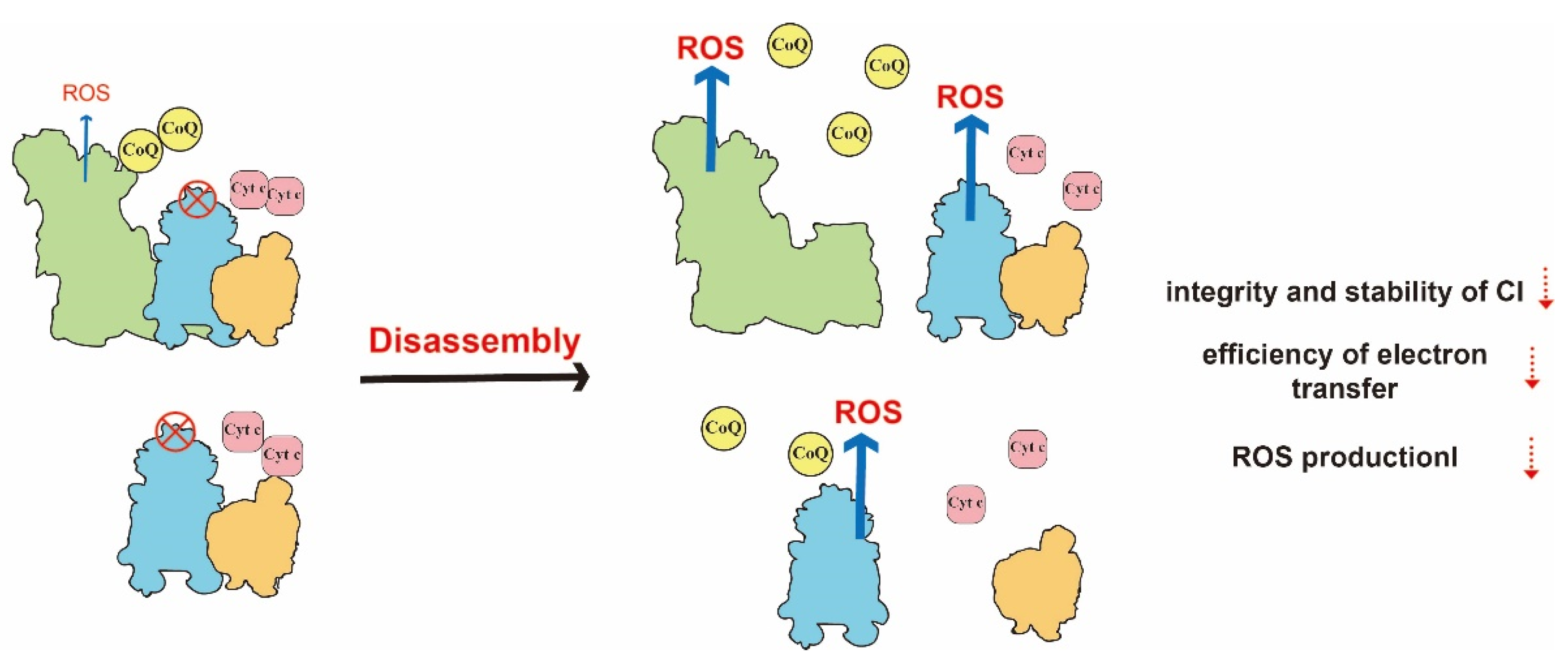
Figure 4.
Illustration of the dysfunction of the respiratory chain induced by the disassembly of SCs. With the defect of CIII as an example, the failed assembly of SCs results in a longer diffusion distance decreased efficiency of electron transfer, and increased ROS production. In addition, the stability of CI is reduced.
Figure 4.
Illustration of the dysfunction of the respiratory chain induced by the disassembly of SCs. With the defect of CIII as an example, the failed assembly of SCs results in a longer diffusion distance decreased efficiency of electron transfer, and increased ROS production. In addition, the stability of CI is reduced.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Structure and important subunits of various SCs in different models.
| Type | Structure | Important Subunit | Model | Ref. |
|---|---|---|---|---|
| CI-CIII-CIV | CICIII2CIV | NDUFB9, NDUFA11, NDUFB8, NDUFB4, ND5, UQCRC1, UQCR11, COX7A, COX7C. | Porcine | [25] |
| CICIII2CIV | NDUFB2, NDUFB3, NDUFB4, NDUFB7, NDUFB9, NDUFB10, NDUFB11, ND5; UQCRQ, UQCRC1, UQCRH, UQCR11, UQCRFS1, COX5b, COX7a, COX7c, COX8b. | Ovine | [12] | |
| CI-CIII | CICIII2 | NDUFB9, NDUFA11, UQCR10, UQCRQ, UQCRC1. | Mammalian | [26] |
| CIII-CIV | CIII2CIV | UQCRFS1, Sub7, Sub10, COX3, COX5B, COX6A2. | Mammalian (mouse and ovine) | [24] |
| CIII2CIV1-2 | Cor1, Cox5a. | Yeast | [27] | |
| CI-CII-CIII-CIV | NA | NA | Mouse | [15] |
| CII-CIII-CIV | NA | NA | Mouse | [15] |
Table 2.
Functions of SCs in various models.
| Function | SCs | Model | Ref. |
|---|---|---|---|
| Maintaining the integrity of free complexes | CICIII2CIV | MT-CYB-deficient human cell line | [13] |
| CICIII2CIV | Mouse L929 cells | [38] | |
| CICIII2CIV | Paracoccus denitrificans | [14] | |
| Improving the efficiency of electron transfer | CIII2CIV | Mammalian (mouse&ovine) | [24] |
| CICIII2CIV; CICIII2; CIII2CIV | Mouse | [16] | |
| CIII2CIV | Yeast | [39] | |
| CICIII2CIV | Ovine | [12] | |
| CIII2CIV1-2 | Yeast | [40] | |
| Regulating ROS production | CICIII2CIV | Ovine | [12] |
| CICIII2CIV | Bovine | [33] | |
| CICIII2 | Bovine | [41] |
Table 3.
Regulatory factors at different regulatory site of SCs in diverse models.
| Regulatory Factors | Regulatory Site | Model | Ref. |
|---|---|---|---|
| SCAF1 | CIII2CIV | Mouse | [16,17,23] |
| CIII2CIV | Ovine | [12] | |
| CIII2CIV | Zebrafish | [45] | |
| HIGD family | |||
| Rcf 1 | CIV | Yeast | [46] |
| HIGD2A | CIV | Mouse C2C12 cells | [44] |
| Phospholipids | CICIII2CIV | Porcine | [32] |
| Cytochrome c | CIII2CIV | Saccharomyces Cerevisiae | [47] |
| CIV | Saccharomyces Cerevisiae | [48] |
Table 4.
The role of SCs in the progression of diseases.
| Disease | SCs | Progress | Ref. |
|---|---|---|---|
| AD | CI–CIII–CIV; CI–CIII; CIII–CIV. | Retention of amyloid at MAM→Failure of the assembly of SCs→Dysfunction of OXPHOS | [22] |
| PD | CICIII2CIV; CICIII2; CIII2CIV. | Absent of Dj1→Defect in the assembly of complex I→Hind assembly of SCs | [56] |
| Barth Syndrome | All SCs | Absent of cardiolipin | [19] |
| Leigh Syndrome | CICIII2CIVn; CICIII2; CIII2CIV. | Mutation in NDUFAF4 | [20] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Guan, S.; Zhao, L.; Peng, R. Mitochondrial Respiratory Chain Supercomplexes: From Structure to Function. Int. J. Mol. Sci. 2022, 23, 13880. https://doi.org/10.3390/ijms232213880
AMA Style
Guan S, Zhao L, Peng R. Mitochondrial Respiratory Chain Supercomplexes: From Structure to Function. International Journal of Molecular Sciences. 2022; 23(22):13880. https://doi.org/10.3390/ijms232213880
Chicago/Turabian StyleGuan, Shuting, Li Zhao, and Ruiyun Peng. 2022. "Mitochondrial Respiratory Chain Supercomplexes: From Structure to Function" International Journal of Molecular Sciences 23, no. 22: 13880. https://doi.org/10.3390/ijms232213880
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.