
Long Preservation of AAV-Transduced Fluorescence by a Modified Organic Solvent-Based Clearing Method


Abstract
:1. Introduction
2. Results
2.1. tDISCO Preserves Endogenous EGFP Fluorescence for Months
2.2. tDISCO Shows the Fast and Effective Clearing Performance on Adult Mouse Brain
2.3. tDISCO Can Be Applied to Clear Various Organs and Visualize Neuronal Morphology
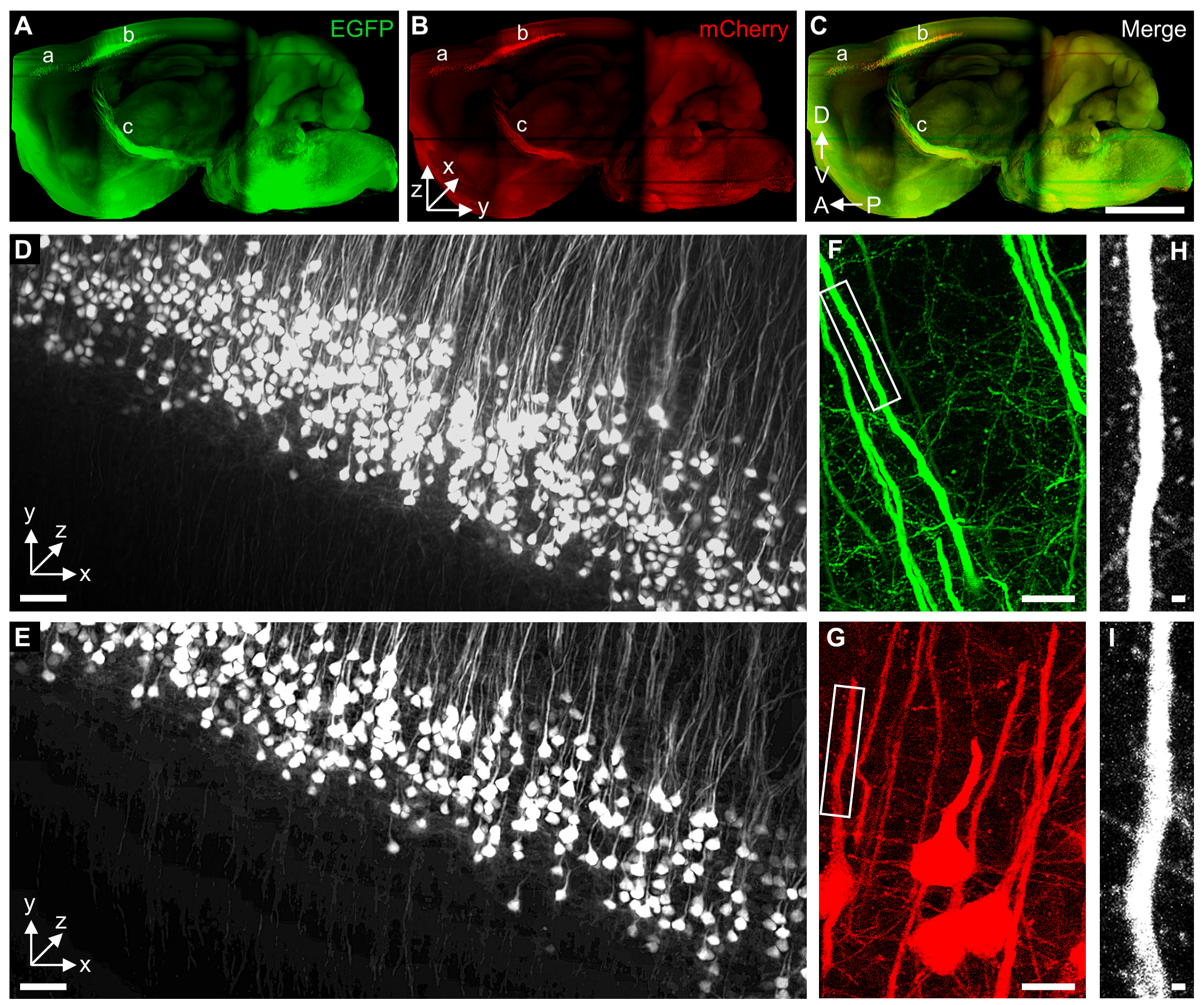
2.4. tDISCO Allows 3D Visualization of Supraspinal Neurons with High-Resolution
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Retrograde Tracing of Supraspinal Inputs to the Cervical Spinal Cord
4.3. Tissue Processing and Imaging
4.4. Modified Organic Solvent-Based Tissue Clearing Method
4.5. Other Organic Solvent-Based Tissue Clearing Methods
4.6. Measurement of Transparency and Tissue Size
4.7. Confocal Fluorescence Microscopy Imaging
4.8. Light-Sheet Fluorescence Microscopy (LSFM) Imaging
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brenna, C.; Simioni, C.; Varano, G.; Conti, I.; Costanzi, E.; Melloni, M.; Neri, L.M. Optical tissue clearing associated with 3D imaging: Application in preclinical and clinical studies. Histochem. Cell Biol. 2022, 157, 497–511. [Google Scholar] [CrossRef] [PubMed]
- Lai, H.M.; Ng, W.L.; Gentleman, S.M.; Wu, W. Chemical Probes for Visualizing Intact Animal and Human Brain Tissue. Cell Chem. Biol. 2017, 24, 659–672. [Google Scholar] [CrossRef] [PubMed]
- Ueda, H.R.; Erturk, A.; Chung, K.; Gradinaru, V.; Chedotal, A.; Tomancak, P.; Keller, P.J. Tissue clearing and its applications in neuroscience. Nat. Rev. Neurosci. 2020, 21, 61–79. [Google Scholar] [CrossRef] [PubMed]
- Erturk, A.; Becker, K.; Jahrling, N.; Mauch, C.P.; Hojer, C.D.; Egen, J.G.; Hellal, F.; Bradke, F.; Sheng, M.; Dodt, H.U. Three-dimensional imaging of solvent-cleared organs using 3DISCO. Nat. Protoc. 2012, 7, 1983–1995. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.; Yu, T.; Xu, J.; Wan, P.; Ma, Y.; Zhu, J.; Li, Y.; Gong, H.; Luo, Q.; Zhu, D. FDISCO: Advanced solvent-based clearing method for imaging whole organs. Sci. Adv. 2019, 5, eaau8355. [Google Scholar] [CrossRef]
- Schwarz, M.K.; Scherbarth, A.; Sprengel, R.; Engelhardt, J.; Theer, P.; Giese, G. Fluorescent-protein stabilization and high-resolution imaging of cleared, intact mouse brains. PLoS ONE 2015, 10, e0124650. [Google Scholar] [CrossRef]
- Pan, C.; Cai, R.; Quacquarelli, F.P.; Ghasemigharagoz, A.; Lourbopoulos, A.; Matryba, P.; Plesnila, N.; Dichgans, M.; Hellal, F.; Erturk, A. Shrinkage-mediated imaging of entire organs and organisms using uDISCO. Nat. Methods 2016, 13, 859–867. [Google Scholar] [CrossRef]
- Susaki, E.A.; Tainaka, K.; Perrin, D.; Kishino, F.; Tawara, T.; Watanabe, T.M.; Yokoyama, C.; Onoe, H.; Eguchi, M.; Yamaguchi, S.; et al. Whole-brain imaging with single-cell resolution using chemical cocktails and computational analysis. Cell 2014, 157, 726–739. [Google Scholar] [CrossRef]
- Hama, H.; Hioki, H.; Namiki, K.; Hoshida, T.; Kurokawa, H.; Ishidate, F.; Kaneko, T.; Akagi, T.; Saito, T.; Saido, T.; et al. ScaleS: An optical clearing palette for biological imaging. Nat. Neurosci. 2015, 18, 1518–1529. [Google Scholar] [CrossRef]
- Ke, M.T.; Fujimoto, S.; Imai, T. SeeDB: A simple and morphology-preserving optical clearing agent for neuronal circuit reconstruction. Nat. Neurosci. 2013, 16, 1154–1161. [Google Scholar] [CrossRef]
- Yang, B.; Treweek, J.B.; Kulkarni, R.P.; Deverman, B.E.; Chen, C.K.; Lubeck, E.; Shah, S.; Cai, L.; Gradinaru, V. Single-cell phenotyping within transparent intact tissue through whole-body clearing. Cell 2014, 158, 945–958. [Google Scholar] [CrossRef] [PubMed]
- Chung, K.; Wallace, J.; Kim, S.Y.; Kalyanasundaram, S.; Andalman, A.S.; Davidson, T.J.; Mirzabekov, J.J.; Zalocusky, K.A.; Mattis, J.; Denisin, A.K.; et al. Structural and molecular interrogation of intact biological systems. Nature 2013, 497, 332–337. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.G.; Sohn, C.H.; Chen, R.; McCue, M.; Yun, D.H.; Drummond, G.T.; Ku, T.; Evans, N.B.; Oak, H.C.; Trieu, W.; et al. Protection of tissue physicochemical properties using polyfunctional crosslinkers. Nat. Biotechnol. 2019, 37, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Richardson, D.S.; Guan, W.; Matsumoto, K.; Pan, C.; Chung, K.; Ertürk, A.; Ueda, H.R.; Lichtman, J.W. Tissue Clearing. Nat. Rev. Methods Primers 2021, 1, 1–24. [Google Scholar] [CrossRef]
- Zhan, Y.; Wu, H.; Liu, L.; Lin, J.; Zhang, S. Organic solvent-based tissue clearing techniques and their applications. J. Biophotonics 2021, 14, e202000413. [Google Scholar] [CrossRef]
- Molbay, M.; Kolabas, Z.I.; Todorov, M.I.; Ohn, T.L.; Erturk, A. A guidebook for DISCO tissue clearing. Mol. Syst. Biol. 2021, 17, e9807. [Google Scholar] [CrossRef]
- Soderblom, C.; Lee, D.H.; Dawood, A.; Carballosa, M.; Jimena Santamaria, A.; Benavides, F.D.; Jergova, S.; Grumbles, R.M.; Thomas, C.K.; Park, K.K.; et al. 3D Imaging of Axons in Transparent Spinal Cords from Rodents and Nonhuman Primates. eNeuro 2015, 2. [Google Scholar] [CrossRef]
- Alnuami, A.A.; Zeedi, B.; Qadri, S.M.; Ashraf, S.S. Oxyradical-induced GFP damage and loss of fluorescence. Int. J. Biol. Macromol. 2008, 43, 182–186. [Google Scholar] [CrossRef]
- Hahn, C.; Becker, K.; Saghafi, S.; Pende, M.; Avdibasic, A.; Foroughipour, M.; Heinz, D.E.; Wotjak, C.T.; Dodt, H.U. High-resolution imaging of fluorescent whole mouse brains using stabilised organic media (sDISCO). J. Biophotonics 2019, 12, e201800368. [Google Scholar] [CrossRef]
- Becker, K.; Jahrling, N.; Saghafi, S.; Weiler, R.; Dodt, H.U. Chemical clearing and dehydration of GFP expressing mouse brains. PLoS ONE 2012, 7, e33916. [Google Scholar] [CrossRef]
- Renier, N.; Wu, Z.; Simon, D.J.; Yang, J.; Ariel, P.; Tessier-Lavigne, M. iDISCO: A simple, rapid method to immunolabel large tissue samples for volume imaging. Cell 2014, 159, 896–910. [Google Scholar] [CrossRef] [PubMed]
- Cai, R.; Pan, C.; Ghasemigharagoz, A.; Todorov, M.I.; Forstera, B.; Zhao, S.; Bhatia, H.S.; Parra-Damas, A.; Mrowka, L.; Theodorou, D.; et al. Panoptic imaging of transparent mice reveals whole-body neuronal projections and skull-meninges connections. Nat. Neurosci. 2019, 22, 317–327. [Google Scholar] [CrossRef] [PubMed]
- Susaki, E.A.; Shimizu, C.; Kuno, A.; Tainaka, K.; Li, X.; Nishi, K.; Morishima, K.; Ono, H.; Ode, K.L.; Saeki, Y.; et al. Versatile whole-organ/body staining and imaging based on electrolyte-gel properties of biological tissues. Nat. Commun. 2020, 11, 1982. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Xu, J.; Wan, P.; Yu, T.; Zhu, D. Optimization of GFP Fluorescence Preservation by a Modified uDISCO Clearing Protocol. Front. Neuroanat. 2018, 12, 67. [Google Scholar] [CrossRef]
- Tervo, D.G.; Hwang, B.Y.; Viswanathan, S.; Gaj, T.; Lavzin, M.; Ritola, K.D.; Lindo, S.; Michael, S.; Kuleshova, E.; Ojala, D.; et al. A Designer AAV Variant Permits Efficient Retrograde Access to Projection Neurons. Neuron 2016, 92, 372–382. [Google Scholar] [CrossRef]
- Wang, Z.; Maunze, B.; Wang, Y.; Tsoulfas, P.; Blackmore, M.G. Global Connectivity and Function of Descending Spinal Input Revealed by 3D Microscopy and Retrograde Transduction. J. Neurosci. 2018, 38, 10566–10581. [Google Scholar] [CrossRef]
- Wang, X.; Liu, Y.; Li, X.; Zhang, Z.; Yang, H.; Zhang, Y.; Williams, P.R.; Alwahab, N.S.A.; Kapur, K.; Yu, B.; et al. Deconstruction of Corticospinal Circuits for Goal-Directed Motor Skills. Cell 2017, 171, 440–455 e414. [Google Scholar] [CrossRef]
- Alkaabi, K.M.; Yafea, A.; Ashraf, S.S. Effect of pH on thermal- and chemical-induced denaturation of GFP. Appl Biochem. Biotechnol. 2005, 126, 149–156. [Google Scholar] [CrossRef]
- Laroche, T.; Burri, O.; Dubey, L.K.; Seitz, A. Development of Sample-Adaptable Holders for Lightsheet Microscopy. Front. Neuroanat. 2019, 13, 26. [Google Scholar] [CrossRef]
- Doherty, G.P.; Bailey, K.; Lewis, P.J. Stage-specific fluorescence intensity of GFP and mCherry during sporulation In Bacillus Subtilis. BMC Res. Notes 2010, 3, 303. [Google Scholar] [CrossRef] [Green Version]
- Shaner, N.C.; Steinbach, P.A.; Tsien, R.Y. A guide to choosing fluorescent proteins. Nat. Methods 2005, 2, 905–909. [Google Scholar] [CrossRef] [PubMed]
- Lu, T.; Shinozaki, M.; Nagoshi, N.; Nakamura, M.; Okano, H. 3D imaging of supraspinal inputs to the thoracic and lumbar spinal cord mapped by retrograde tracing and light-sheet microscopy. J. Neurochem. 2022, 162, 352–370. [Google Scholar] [CrossRef]
- Peters, A.J.; Lee, J.; Hedrick, N.G.; O’Neil, K.; Komiyama, T. Reorganization of corticospinal output during motor learning. Nat. Neurosci. 2017, 20, 1133–1141. [Google Scholar] [CrossRef] [PubMed]
- Bekkouche, B.M.B.; Fritz, H.K.M.; Rigosi, E.; O’Carroll, D.C. Comparison of Transparency and Shrinkage During Clearing of Insect Brains Using Media With Tunable Refractive Index. Front. Neuroanat. 2020, 14, 599282. [Google Scholar] [CrossRef]
- Wang, Z.; Bovik, A.C.; Sheikh, H.R.; Simoncelli, E.P. Image quality assessment: From error visibility to structural similarity. IEEE Trans. Image Process. 2004, 13, 600–612. [Google Scholar] [CrossRef] [PubMed]
- Royer, L.A.; Weigert, M.; Gunther, U.; Maghelli, N.; Jug, F.; Sbalzarini, I.F.; Myers, E.W. ClearVolume: Open-source live 3D visualization for light-sheet microscopy. Nat. Methods 2015, 12, 480–481. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reagents (Alkaline pH) | Brain Slices (1 mm); Spinal Cord | Brain Hemisphere; Internal Organs a | Whole Adult Brain |
|---|---|---|---|
| 50% (v/v) a-THF | 30 min | 3 h | 6 h |
| 70% (v/v) a-THF | - | - | 6 h |
| 80% (v/v) a-THF | 30 min | 3 h | 12 h |
| 100% (v/v) a-THF | 2 × 30 min | 3 h, ON | 2 × 12 h |
| a-BABB-D4 | ≥15 min | ≥1.5 h | ≥3 h |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, T.; Shinozaki, M.; Nagoshi, N.; Nakamura, M.; Okano, H. Long Preservation of AAV-Transduced Fluorescence by a Modified Organic Solvent-Based Clearing Method. Int. J. Mol. Sci. 2022, 23, 9637. https://doi.org/10.3390/ijms23179637
Lu T, Shinozaki M, Nagoshi N, Nakamura M, Okano H. Long Preservation of AAV-Transduced Fluorescence by a Modified Organic Solvent-Based Clearing Method. International Journal of Molecular Sciences. 2022; 23(17):9637. https://doi.org/10.3390/ijms23179637
Chicago/Turabian StyleLu, Tao, Munehisa Shinozaki, Narihito Nagoshi, Masaya Nakamura, and Hideyuki Okano. 2022. "Long Preservation of AAV-Transduced Fluorescence by a Modified Organic Solvent-Based Clearing Method" International Journal of Molecular Sciences 23, no. 17: 9637. https://doi.org/10.3390/ijms23179637