
Overexpression of microRNAs miR-25-3p, miR-185-5p and miR-132-3p in Late Onset Fetal Growth Restriction, Validation of Results and Study of the Biochemical Pathways Involved

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
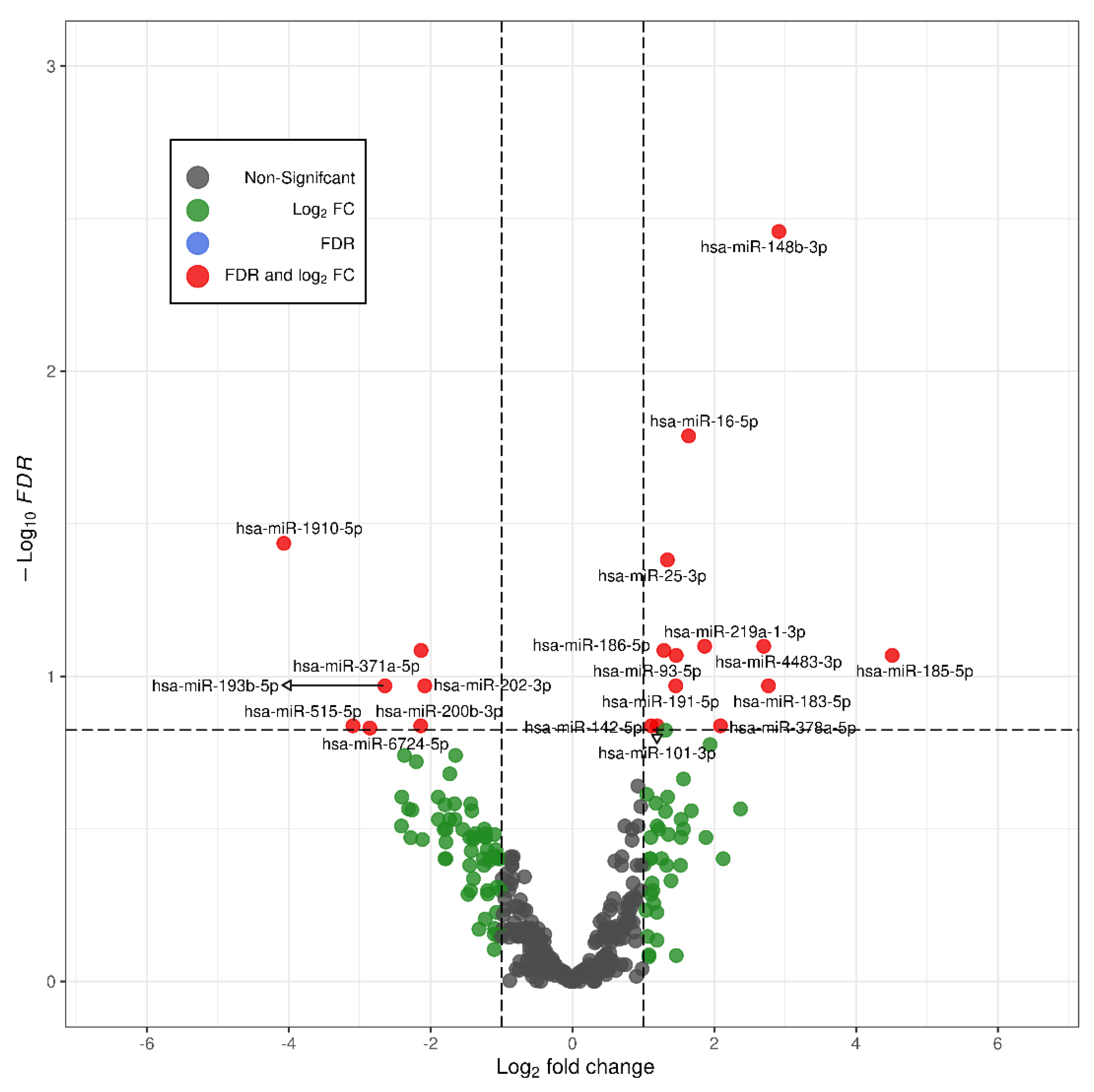
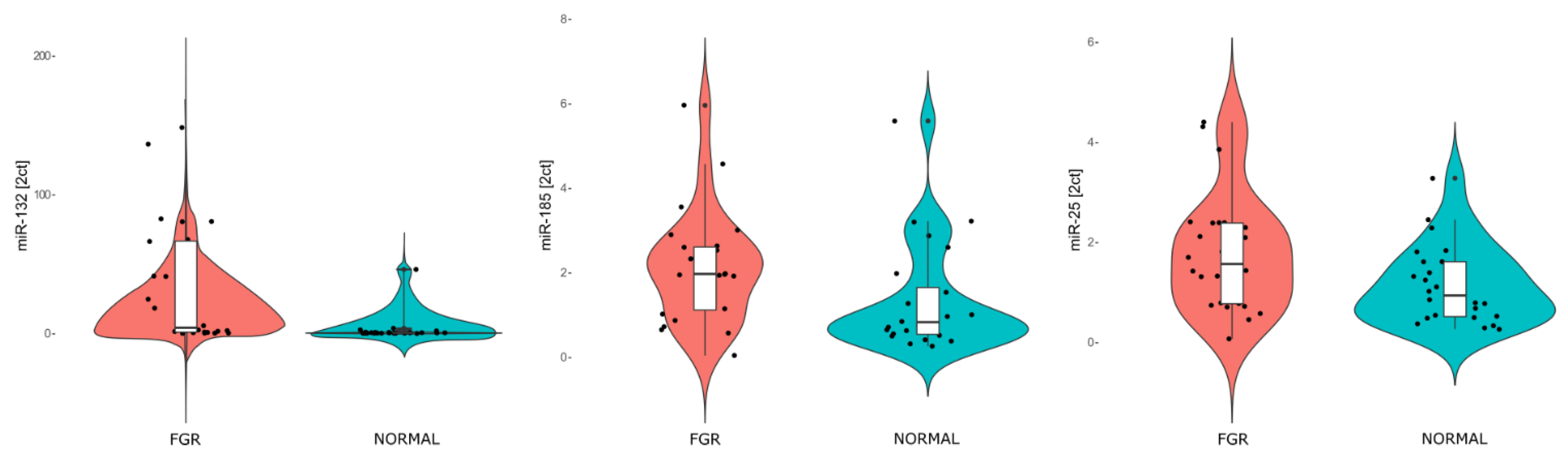
2.1. Differential Expression of miRNAs in FGR
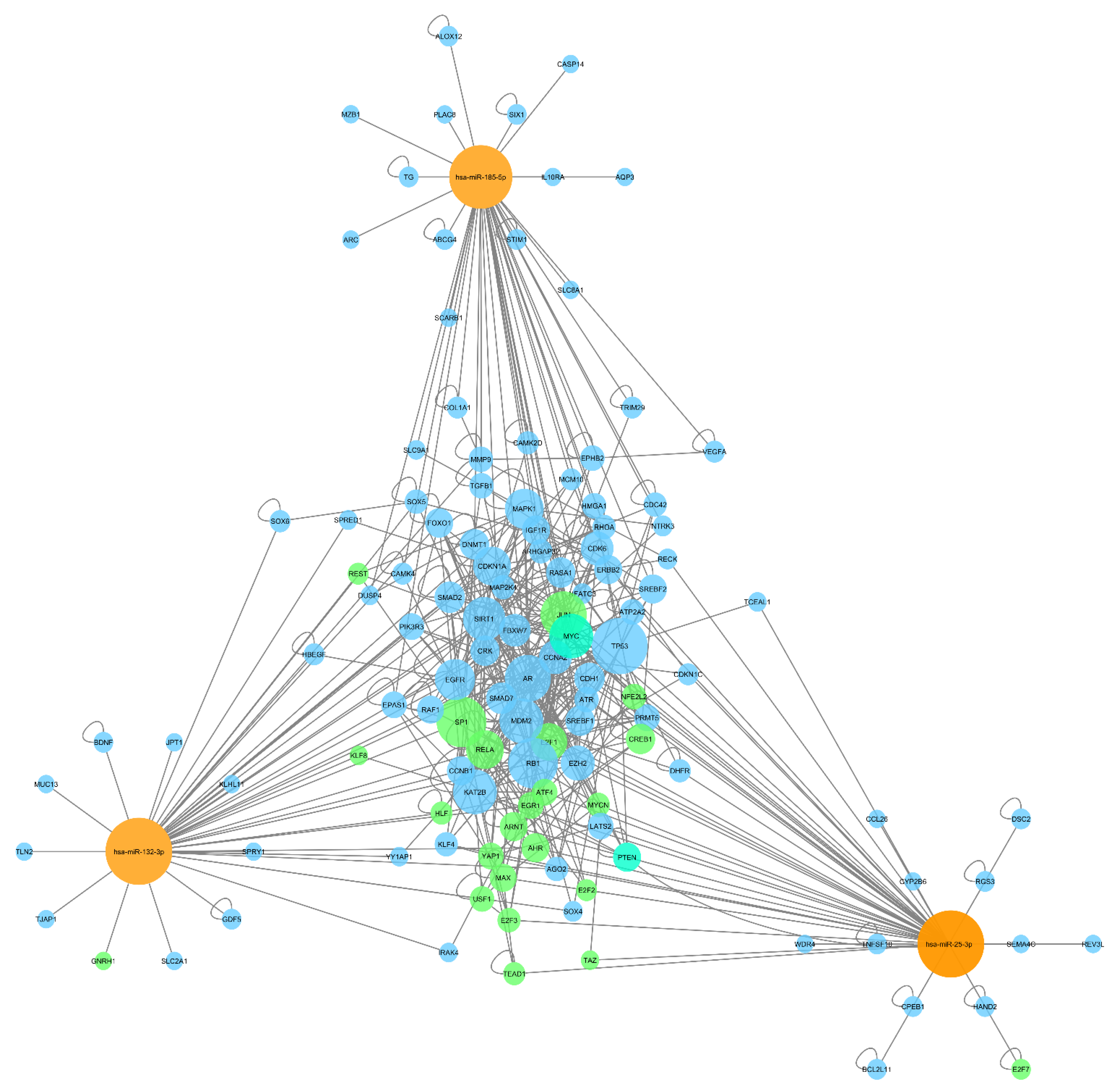
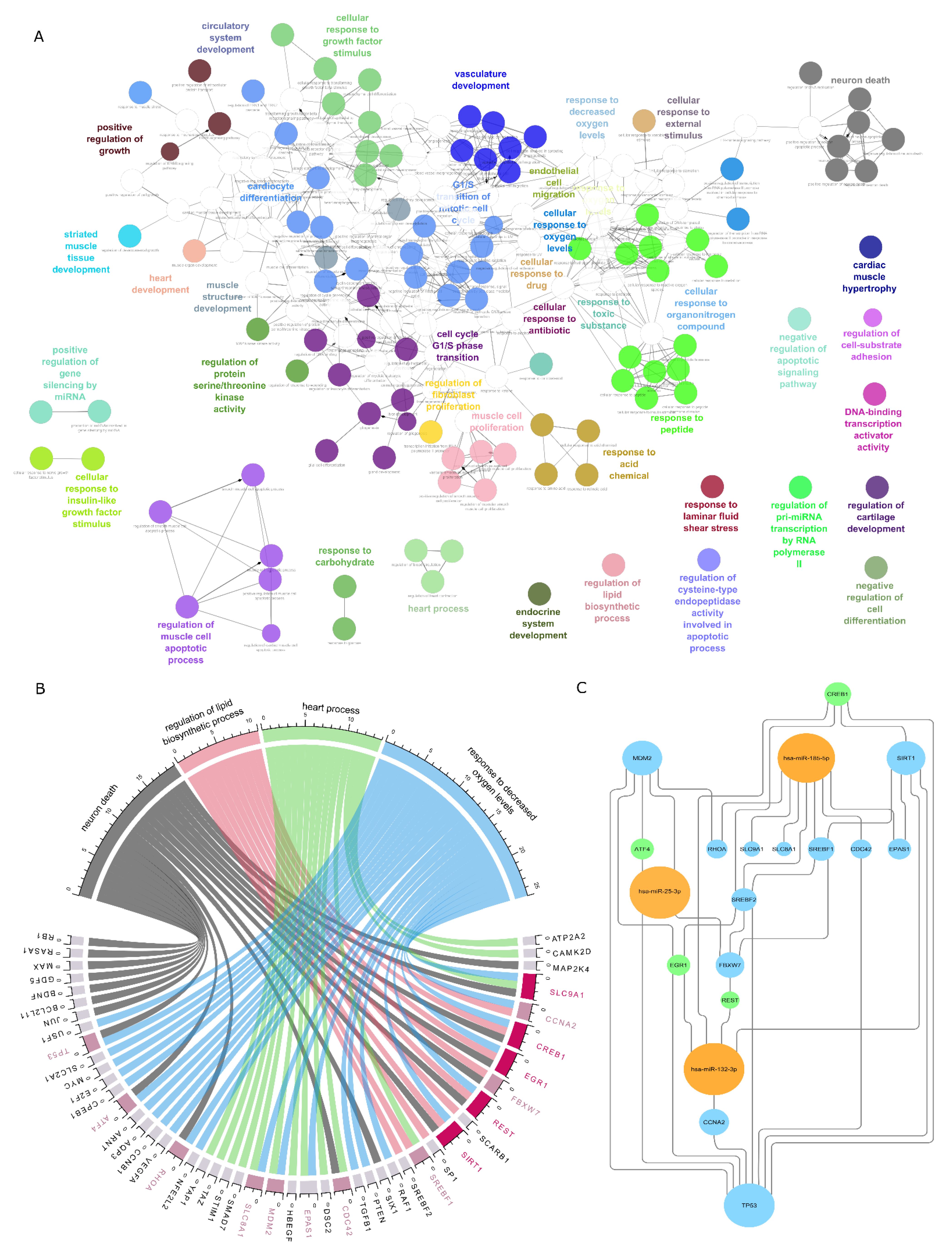
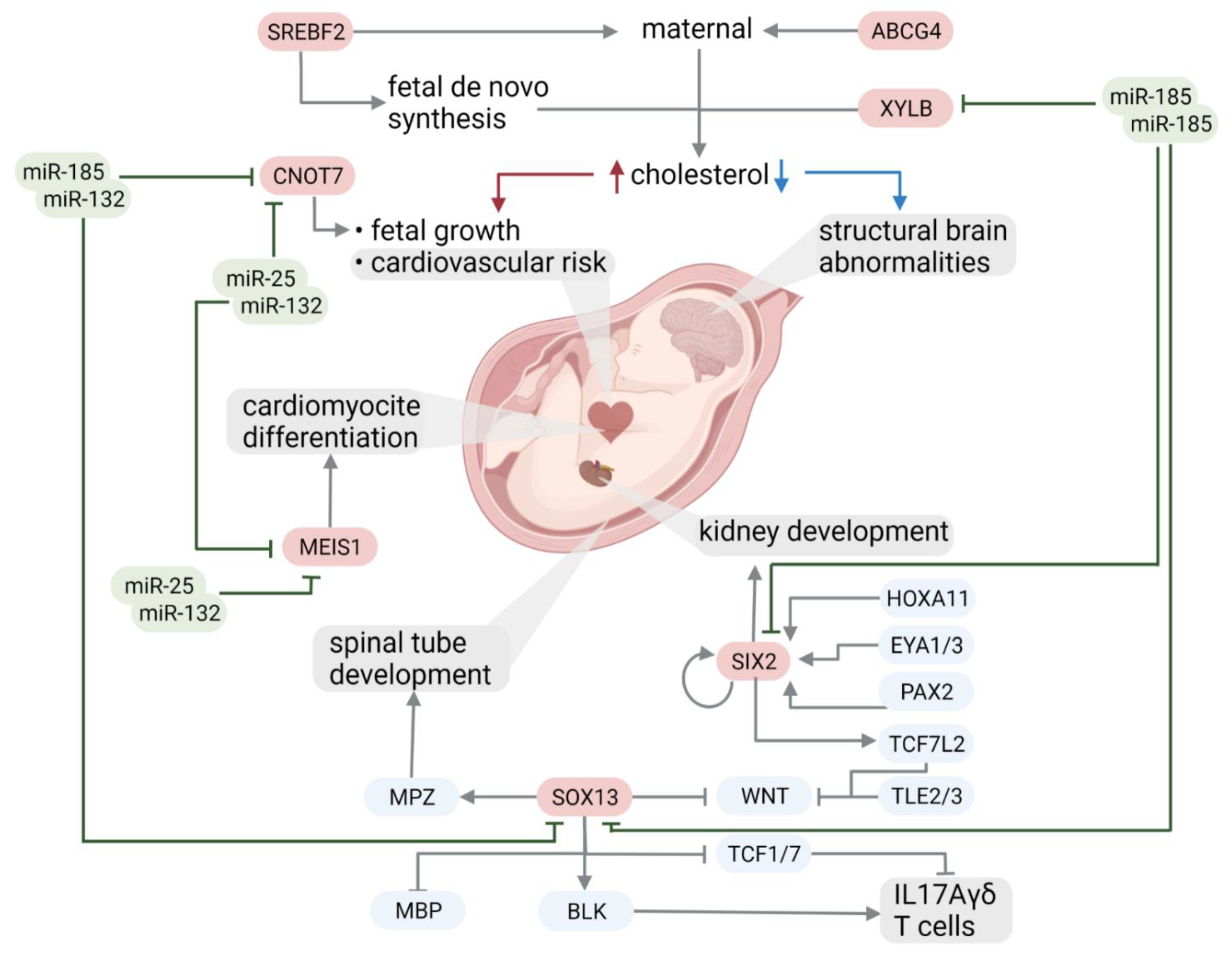
2.2. Functional Analysis of Deregulated miRNA
2.3. Interacting miRNA Triplex Formation
2.4. Limitation and Novelty
3. Discussions
3.1. Intrauterine Environment as the Origin of Adult Disease
3.2. miR-25-3p
3.3. miR-185-5p
3.4. miR-132-3p
4. Materials and Methods
4.1. Patient Recruitment
4.2. Quantitative Reverse Transcription Polymerase Chain Reaction (RT-qPCR) Validation in Umbilical Cord Samples
4.3. Gene Target Identification and Network Analysis
4.4. Overrepresentation and Functional Analysis
4.5. Synergistically Working miRNA
4.6. Statistical Analysis
5. Clinical Significance
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gordijn, S.; Beune, I.; Thilaganathan, B.; Papageorghiou, A.; Baschat, A.; Baker, P.; Silver, R.; Wynia, K.; Ganzevoort, W. Consensus definition of fetal growth restriction: A Delphi procedure. Ultrasound Obstet. Gynecol. 2016, 48, 333–339. [Google Scholar] [CrossRef] [PubMed]
- Spinillo, A.; Gardella, B.; Adamo, L.; Muscettola, G.; Fiandrino, G.; Cesari, S. Pathologic placental lesions in early and late fetal growth restriction. Acta Obstet. et Gynecol. Scand. 2019, 98, 1585–1594. [Google Scholar] [CrossRef] [Green Version]
- Morales-Roselló, J.; Buongiorno, S.; Loscalzo, G.; Alfonso, B.-R.V.; Hervas-Marín, D.; Perales-Marín, A. Mathematical simulation of Doppler changes in late-onset smallness; progression patterns of cerebral and umbilical anomalies define two types of late-onset fetal growth restriction. J. Matern. Neonatal Med. 2021, 34, 1–11. [Google Scholar] [CrossRef]
- Figueras, F.; Gardosi, J. Intrauterine growth restriction: New concepts in antenatal surveillance, diagnosis, and management. Am. J. Obstet. Gynecol. 2011, 204, 288–300. [Google Scholar] [CrossRef] [PubMed]
- Mifsud, W.; Sebire, N.J. Placental Pathology in Early-Onset and Late-Onset Fetal Growth Restriction. Fetal Diagn. Ther. 2014, 36, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Lu, Q.; Chang, C. Epigenetics in Health and Disease. Adv. Exp. Med. Biol. 2020, 1253, 3–55. [Google Scholar] [CrossRef]
- Zoghbi, H.Y.; Beaudet, A.L. Epigenetics and Human Disease. Cold Spring Harb. Perspect. Biol. 2016, 8, a019497. [Google Scholar] [CrossRef]
- Morales-Prieto, D.M.; Ospina-Prieto, S.; Chaiwangyen, W.; Schoenleben, M.; Markert, U.R. Pregnancy-associated miRNA-clusters. J. Reprod. Immunol. 2013, 97, 51–61. [Google Scholar] [CrossRef]
- Hemmatzadeh, M.; Shomali, N.; Yousefzadeh, Y.; Mohammadi, H.; Ghasemzadeh, A.; Yousefi, M. MicroRNAs: Small molecules with a large impact on pre-eclampsia. J. Cell. Physiol. 2020, 235, 3235–3248. [Google Scholar] [CrossRef] [PubMed]
- Poddar, S.; Kesharwani, D.; Datta, M. Interplay between the miRNome and the epigenetic machinery: Implications in health and disease. J. Cell. Physiol. 2017, 232, 2938–2945. [Google Scholar] [CrossRef]
- Tsochandaridis, M.; Nasca, L.; Toga, C.; Levy-Mozziconacci, A. Circulating MicroRNAs as Clinical Biomarkers in the Predictions of Pregnancy Complications. BioMed Res. Int. 2015, 2015, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayder, H.; O’Brien, J.; Nadeem, U.; Peng, C. MicroRNAs: Crucial regulators of placental development. Reproduction 2018, 155, R259–R271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gross, N.; Kropp, J.; Khatib, H. MicroRNA Signaling in Embryo Development. Biology 2017, 6, 34. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.-S.; Ishibashi, O.; Ishikawa, G.; Ishikawa, T.; Katayama, A.; Mishima, T.; Takizawa, T.; Shigihara, T.; Goto, T.; Izumi, A.; et al. Human Villous Trophoblasts Express and Secrete Placenta-Specific MicroRNAs into Maternal Circulation via Exosomes. Biol. Reprod. 2009, 81, 717–729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kochhar, P.; Vukku, M.; Rajashekhar, R.; Mukhopadhyay, A. microRNA signatures associated with fetal growth restriction: A systematic review. Eur. J. Clin. Nutr. 2021, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Liu, M.; Yu, H.; Zhang, J.; Zhou, R. Circulating microRNAs as biomarkers for diagnosis and prediction of preeclampsia: A systematic review and meta-analysis. Eur. J. Obstet. Gynecol. Reprod. Biol. 2020, 253, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Kanak, M.; Alseiari, M.; Balasubramanian, P.; Addanki, K.; Aggarwal, M.; Noorali, S.; Kalsum, A.; Mahalingam, K.; Pace, G.; Panasik, N.; et al. Triplex-forming MicroRNAs form stable complexes with HIV-1 provirus and inhibit its replication. Appl. Immunohistochem. Mol. Morphol. 2010, 18, 532–545. [Google Scholar] [CrossRef] [PubMed]
- Valadi, H.; Ekstrom, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Lötvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmitz, U.; Lai, X.; Winter, F.; Wolkenhauer, O.; Vera, J.; Gupta, S.K. Cooperative gene regulation by microRNA pairs and their identification using a computational workflow. Nucleic Acids Res. 2014, 42, 7539–7552. [Google Scholar] [CrossRef] [Green Version]
- Morales-Roselló, J.; García-Giménez, J.L.; Priego, L.M.; González-Rodríguez, D.; Mena-Mollá, S.; Catalá, A.M.; Loscalzo, G.; Buongiorno, S.; Jakaite, V.; Martínez, A.J.C.; et al. MicroRNA-148b-3p and MicroRNA-25-3p Are Overexpressed in Fetuses with Late-Onset Fetal Growth Restriction. Fetal Diagn. Ther. 2020, 47, 665–674. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.-Y.; Lin, Y.-C.-D.; Li, J.; Huang, K.-Y.; Shrestha, S.; Hong, H.-C.; Tang, Y.; Chen, Y.-G.; Jin, C.-N.; Yu, Y.; et al. miRTarBase 2020: Updates to the experimentally validated microRNA–target interaction database. Nucleic Acids Res. 2020, 48, D148–D154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, T.; Yao, J.; Liu, Z. Gene Ontology, Enrichment Analysis, and Pathway Analysis. In Bioinformatics in Aquaculture; Wiley: Hoboken, NJ, USA, 2017; pp. 150–168. [Google Scholar]
- Miller, S.L.; Hüppi, P.; Mallard, C. The consequences of fetal growth restriction on brain structure and neurodevelopmental outcome. J. Physiol. 2016, 594, 807–823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miranda, J.; Simoes, R.V.; Paules, C.; Cañueto, D.; Pardo-Cea, M.A.; García-Martín, M.L.; Crovetto, F.; Fuertes-Martin, R.; Domenech, M.; Gómez-Roig, M.D.; et al. Metabolic profiling and targeted lipidomics reveals a disturbed lipid profile in mothers and fetuses with intrauterine growth restriction. Sci. Rep. 2018, 8, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Cohen, E.; Wong, F.Y.; Horne, R.S.; Yiallourou, S. Intrauterine growth restriction: Impact on cardiovascular development and function throughout infancy. Pediatr. Res. 2016, 79, 821–830. [Google Scholar] [CrossRef] [Green Version]
- Malhotra, A.; Allison, B.J.; Castillo-Melendez, M.; Jenkin, G.; Polglase, G.R.; Miller, S.L. Neonatal Morbidities of Fetal Growth Restriction: Pathophysiology and Impact. Front. Endocrinol. 2019, 10, 55. [Google Scholar] [CrossRef] [Green Version]
- Ramos, P.I.P.; Arge, L.W.P.; Lima, N.C.B.; Fukutani, K.F.; De Queiroz, A.T.L. Leveraging User-Friendly Network Approaches to Extract Knowledge From High-Throughput Omics Datasets. Front. Genet. 2019, 10, 1120. [Google Scholar] [CrossRef]
- Kobayashi, A.; Valerius, M.T.; Mugford, J.W.; Carroll, T.J.; Self, M.; Oliver, G.; McMahon, A.P. Six2 Defines and Regulates a Multipotent Self-Renewing Nephron Progenitor Population throughout Mammalian Kidney Development. Cell Stem Cell 2008, 3, 169–181. [Google Scholar] [CrossRef] [Green Version]
- Lindgren, I.M.; Drake, R.R.; Chattergoon, N.N.; Thornburg, K.L. Down-regulation of MEIS1 promotes the maturation of oxidative phosphorylation in perinatal cardiomyocytes. FASEB J. 2019, 33, 7417–7426. [Google Scholar] [CrossRef] [PubMed]
- Marfil, V.; Moya, M.; Pierreux, C.E.; Castell, J.V.; Lemaigre, F.P.; Real, F.X.; Bort, R. Interaction between Hhex and SOX13 Modulates Wnt/TCF Activity. J. Biol. Chem. 2010, 285, 5726–5737. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.; Fukuda, Y.; Schultz, R.M. Mobilization of Dormant Cnot7 mRNA Promotes Deadenylation of Maternal Transcripts During Mouse Oocyte Maturation1. Biol. Reprod. 2015, 93, 48. [Google Scholar] [CrossRef] [PubMed]
- Bunker, R.; Bulloch, E.; Dickson, J.M.J.; Loomes, K.M.; Baker, E.N. Structure and Function of Human Xylulokinase, an Enzyme with Important Roles in Carbohydrate Metabolism. J. Biol. Chem. 2013, 288, 1643–1652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vlachos, I.S.; Paraskevopoulou, M.D.; Karagkouni, D.; Georgakilas, G.; Vergoulis, T.; Kanellos, I.; Anastasopoulos, I.-L.; Maniou, S.; Karathanou, K.; Kalfakakou, D.; et al. DIANA-TarBase v7.0: Indexing more than half a million experimentally supported miRNA:mRNA interactions. Nucleic Acids Res. 2015, 43, D153–D159. [Google Scholar] [CrossRef] [PubMed]
- Kesavan, K.; Devaskar, S.U. Intrauterine Growth Restriction. Pediatr. Clin. N. Am. 2019, 66, 403–423. [Google Scholar] [CrossRef] [PubMed]
- Doan, T.N.A.; Briffa, J.F.; Phillips, A.L.; Leemaqz, S.Y.; Burton, R.A.; Romano, T.; Wlodek, M.E.; Bianco-Miotto, T. Epigenetic mechanisms involved in intrauterine growth restriction and aberrant kidney development and function. J. Dev. Orig. Health Dis. 2020, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.-N.; Tain, Y.-L. Developmental Programming and Reprogramming of Hypertension and Kidney Disease: Impact of Tryptophan Metabolism. Int. J. Mol. Sci. 2020, 21, 8705. [Google Scholar] [CrossRef]
- Gardella, B.; Dominoni, M.; Caporali, C.; Cesari, S.; Fiandrino, G.; Longo, S.; De Vito, G.B.; Naboni, C.; Tonduti, D.; Perotti, G.; et al. Placental features of fetal vascular malperfusion and infant neurodevelopmental outcomes at 2 years of age in severe fetal growth restriction. Am. J. Obstet. Gynecol. 2021, 225, 413.e1–413.e11. [Google Scholar] [CrossRef] [PubMed]
- Crispi, F.; Miranda, J.; Gratacós, E. Long-term cardiovascular consequences of fetal growth restriction: Biology, clinical implications, and opportunities for prevention of adult disease. Am. J. Obstet. Gynecol. 2018, 218, S869–S879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Senra, J.C.; Yoshizaki, C.T.; Doro, G.F.; Ruano, R.; Gibelli, M.A.B.C.; Rodrigues, A.S.; Koch, V.; Krebs, V.L.J.; Zugaib, M.; Francisco, R.P.V.; et al. Kidney impairment in fetal growth restriction: Three-dimensional evaluation of volume and vascularization. Prenat. Diagn. 2020, 40, 1408–1417. [Google Scholar] [CrossRef]
- Herrera, E.A.; González-Candia, A. Gestational Hypoxia and Blood-Brain Barrier Permeability: Early Origins of Cerebrovascular Dysfunction Induced by Epigenetic Mechanisms. Front. Physiol. 2021, 12. [Google Scholar] [CrossRef]
- Benítez-Marín, M.J.; Marín-Clavijo, J.; Blanco-Elena, J.A.; Jiménez-López, J.; González-Mesa, E. Brain Sparing Effect on Neurodevelopment in Children with Intrauterine Growth Restriction: A Systematic Review. Children 2021, 8, 745. [Google Scholar] [CrossRef]
- Baschat, A.A. Neurodevelopment after Fetal Growth Restriction. Fetal Diagn. Ther. 2014, 36, 136–142. [Google Scholar] [CrossRef]
- Sárközy, M.; Kahán, Z.; Csont, T. A myriad of roles of miR-25 in health and disease. Oncotarget 2018, 9, 21580–21612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.; Liu, C.; Li, Q.; Li, J.; Wu, Y.; Liu, J. MicroRNA-25–5p counteracts oxidized LDL-induced pathological changes by targeting neuronal growth regulator 1 (NEGR1) in human brain micro-vessel endothelial cells. Biochimie 2019, 165, 141–149. [Google Scholar] [CrossRef]
- Zhang, B.; Zhang, G.; Wei, T.; Yang, Z.; Tan, W.; Mo, Z.; Liu, J.; Li, N.; Wei, Y.; Zhang, L.; et al. MicroRNA-25 Protects Smooth Muscle Cells against Corticosterone-Induced Apoptosis. Oxidative Med. Cell. Longev. 2019, 2019, 2691514. [Google Scholar] [CrossRef]
- Wu, X.; Zhou, H.; Yue, B.; Li, M.; Liu, F.; Qiu, C.; Chen, B.; Ma, X. Upregulation of microRNA-25-3p inhibits proliferation, migration and invasion of osteosarcoma cells in vitro by directly targeting SOX4. Mol. Med. Rep. 2017, 16, 4293–4300. [Google Scholar] [CrossRef]
- Li, R.; Wen, Y.; Wu, B.; He, M.; Zhang, P.; Zhang, Q.; Chen, Y. MicroRNA-25-3p suppresses epileptiform discharges through inhibiting oxidative stress and apoptosis via targeting OXSR1 in neurons. Biochem. Biophys. Res. Commun. 2020, 523, 859–866. [Google Scholar] [CrossRef] [PubMed]
- Yzydorczyk, C.; Armengaud, J.B.; Peyter, A.-C.; Chehade, H.; Cachat, F.; Juvet, C.; Siddeek, B.; Simoncini, S.; Sabatier, F.; Dignat-George, F.; et al. Endothelial dysfunction in individuals born after fetal growth restriction: Cardiovascular and renal consequences and preventive approaches. J. Dev. Orig. Heal. Dis. 2017, 8, 448–464. [Google Scholar] [CrossRef]
- Figueras, F.; Caradeux, J.; Crispi, F.; Eixarch, E.; Peguero, A.; Gratacos, E. Diagnosis and surveillance of late-onset fetal growth restriction. Am. J. Obstet. Gynecol. 2018, 218, S790–S802.e1. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, A.; Perkins, J. Angiogenesis and intrauterine growth restriction. Best Pr. Res. Clin. Obstet. Gynaecol. 2000, 14, 981–998. [Google Scholar] [CrossRef]
- Serocki, M.; Bartoszewska, S.; Janaszak-Jasiecka, A.; Ochocka, R.J.; Collawn, J.F.; Bartoszewski, R. miRNAs regulate the HIF switch during hypoxia: A novel therapeutic target. Angiogenesis 2018, 21, 183–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sifakis, S.; Gourvas, V.; Dalpa, E.; Konstantinidou, A.; Vrachnis, N.; Spandidos, D.A. Angiogenic factors in placentas from pregnancies complicated by fetal growth restriction (Review). Mol. Med. Rep. 2012, 6, 23–27. [Google Scholar] [CrossRef] [PubMed]
- Storkebaum, E.; Lambrechts, D.; Carmeliet, P. VEGF: Once regarded as a specific angiogenic factor, now implicated in neuroprotection. BioEssays 2004, 26, 943–954. [Google Scholar] [CrossRef] [PubMed]
- Fang, M.; Li, Y.; Wu, Y.; Ning, Z.; Wang, X.; Li, X. miR-185 silencing promotes the progression of atherosclerosis via targeting stromal interaction molecule 1. Cell Cycle 2019, 18, 682–695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Youssef, L.; Miranda, J.; Paules, C.; Garcia-Otero, L.; Vellvé, K.; Kalapotharakos, G.; Sepulveda-Martinez, A.; Crovetto, F.; Gomez, O.; Gratacós, E.; et al. Fetal cardiac remodeling and dysfunction is associated with both preeclampsia and fetal growth restriction. Am. J. Obstet. Gynecol. 2020, 222, 79.e1–79.e9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crispi, F.; Figueras, F.; Cruz-Lemini, M.; Bartrons, J.; Bijnens, B.; Gratacos, E. Cardiovascular programming in children born small for gestational age and relationship with prenatal signs of severity. Am. J. Obstet. Gynecol. 2012, 207, 121.e1–121.e9. [Google Scholar] [CrossRef] [PubMed]
- Crispi, F.; Rodríguez-López, M.; Bernardino, G.; Sepúlveda-Martínez, Á.; Prat-González, S.; Pajuelo, C.; Perea, R.J.; Caralt, M.T.; Casu, G.; Vellvé, K.; et al. Exercise Capacity in Young Adults Born Small for Gestational Age. JAMA Cardiol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Pecks, U.; Bornemann, V.; Klein, A.; Segger, L.; Maass, N.; Alkatout, I.; Eckmann-Scholz, C.; Elessawy, M.; Lütjohann, D. Estimating fetal cholesterol synthesis rates by cord blood analysis in intrauterine growth restriction and normally grown fetuses. Lipids Health Dis. 2019, 18, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pecks, U.; Rath, W.; Bauerschlag, D.O.; Maass, N.; Orlikowsky, T.; Mohaupt, M.G.; Escher, G. Serum cholesterol acceptor capacity in intrauterine growth restricted fetuses. J. Périnat. Med. 2017, 45, 829–835. [Google Scholar] [CrossRef]
- Yang, H.-X.; Zhang, M.; Long, S.-Y.; Tuo, Q.-H.; Tian, Y.; Chen, J.-X.; Zhang, C.-P.; Liao, D.-F. Cholesterol in LDL receptor recycling and degradation. Clin. Chim. Acta 2020, 500, 81–86. [Google Scholar] [CrossRef]
- Cziraky, M.J.; Watson, K.E.; Talbert, R.L. Targeting low HDL-cholesterol to decrease residual cardiovascular risk in the managed care setting. J. Manag. Care Pharmacy 2008, 14, 1–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tzschoppe, A.; Von Kries, R.; Struwe, E.; Rascher, W.; Dörr, H.-G.; Jüngert, J.; Rauh, M.; Beckmann, M.; Schild, R.; Goecke, T.W.; et al. Intrauterine Growth Restriction (IUGR) Induces Signs of Subclinical Atherosclerosis in 6-Year-Old Infants Despite Absence Of Excessive Growth. Klinische Pädiatrie 2017, 229, 209–215. [Google Scholar] [CrossRef]
- Yang, M.; Liu, W.; Pellicane, C.; Sahyoun, C.; Joseph, B.K.; Gallo-Ebert, C.; Donigan, M.; Pandya, D.; Giordano, C.; Bata, A.; et al. Identification of miR-185 as a regulator of de novo cholesterol biosynthesis and low density lipoprotein uptake. J. Lipid Res. 2014, 55, 226–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dlouha, D.; Blaha, M.; Blaha, V.; Fatorova, I.; Hubacek, J.A.; Stavek, P.; Lanska, V.; Parikova, A.; Pitha, J. Analysis of circulating miRNAs in patients with familial hypercholesterolaemia treated by LDL/Lp(a) apheresis. Atheroscler. Suppl. 2017, 30, 128–134. [Google Scholar] [CrossRef]
- Haas, J.D.; Ravens, S.; Düber, S.; Sandrock, I.; Oberdörfer, L.; Kashani, E.; Chennupati, V.; Föhse, L.; Naumann, R.; Weiss, S.; et al. Development of Interleukin-17-Producing γδ T Cells Is Restricted to a Functional Embryonic Wave. Immunity 2012, 37, 48–59. [Google Scholar] [CrossRef] [Green Version]
- Mostafa, D.; Takahashi, A.; Yanagiya, A.; Yamaguchi, T.; Abe, T.; Kureha, T.; Kuba, K.; Kanegae, Y.; Furuta, Y.; Yamamoto, T.; et al. Essential functions of the CNOT7/8 catalytic subunits of the CCR4-NOT complex in mRNA regulation and cell viability. RNA Biol. 2020, 17, 403–416. [Google Scholar] [CrossRef]
- Yu, J.; Hu, X.; Chen, X.; Zhou, Q.; Jiang, Q.; Shi, Z.; Zhu, H. CNOT7 modulates biological functions of ovarian cancer cells via AKT signaling pathway. Life Sci. 2021, 268, 118996. [Google Scholar] [CrossRef]
- Wang, N.; Yang, Q.; Gu, Y.; Zhen, X.; Shi, Y.; Gu, W.; Wang, J.; He, Y.; Wang, J. MNSFβ Promotes the Proliferation and Migration of Human Extravillous Trophoblast Cells and the Villus Expression Level of MNSFβ Is Decreased in Recurrent Miscarriage Patients. Gynecol. Obstet. Investig. 2021, 86, 27–39. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Hu, S.; Wu, Z.; Liu, J.; Li, S. The Role of MiR-132 in Regulating Neural Stem Cell Proliferation, Differentiation and Neuronal Maturation. Cell. Physiol. Biochem. 2018, 47, 2319–2330. [Google Scholar] [CrossRef] [PubMed]
- Qazi, T.J.; Lu, J.; Duru, L.; Zhao, J.; Qing, H. Upregulation of mir-132 induces dopaminergic neuronal death via activating SIRT1/P53 pathway. Neurosci. Lett. 2021, 740, 135465. [Google Scholar] [CrossRef]
- Qian, Y.; Song, J.; Ouyang, Y.; Han, Q.; Chen, W.; Zhao, X.; Xie, Y.; Chen, Y.; Yuan, W.; Fan, C. Advances in Roles of miR-132 in the Nervous System. Front. Pharmacol. 2017, 8, 770. [Google Scholar] [CrossRef] [PubMed]
- Rafat, M.; Moraghebi, M.; Afsa, M.; Malekzadeh, K. The outstanding role of miR-132-3p in carcinogenesis of solid tumors. Hum. Cell 2021, 34, 1051–1065. [Google Scholar] [CrossRef]
- Hanin, G.; Yayon, N.; Tzur, Y.; Haviv, R.; Bennett, E.R.; Udi, S.; Krishnamoorthy, Y.R.; Kotsiliti, E.; Zangen, R.; Efron, B.; et al. miRNA-132 induces hepatic steatosis and hyperlipidaemia by synergistic multitarget suppression. Gut 2017, 67, 1124–1134. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Lee, M.-S.; Jung, S.; Son, H.-Y.; Park, S.; Kang, B.; Kim, S.-Y.; Kim, I.-H.; Kim, C.-T.; Kim, Y. Ginger Extract Ameliorates Obesity and Inflammation via Regulating MicroRNA-21/132 Expression and AMPK Activation in White Adipose Tissue. Nutrients 2018, 10, 1567. [Google Scholar] [CrossRef] [Green Version]
- Ucar, A.; Gupta, S.K.; Fiedler, J.; Erikci, E.; Kardasinski, M.; Batkai, S.; Dangwal, S.; Kumarswamy, R.; Bang, C.; Holzmann, A.; et al. The miRNA-212/132 family regulates both cardiac hypertrophy and cardiomyocyte autophagy. Nat. Commun. 2012, 3, 1078. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Li, K.; Liu, L.; Li, S. MicroRNA-132 promotes oxidative stress-induced pyroptosis by targeting sirtuin 1 in myocardial ischaemia-reperfusion injury. Int. J. Mol. Med. 2020, 45, 1942–1950. [Google Scholar] [CrossRef] [PubMed]
- Figueras, F.; Meler, E.; Iraola, A.; Eixarch, E.; Coll, O.; Francis, A.; Gratacos, E.; Gardosi, J. Customized birthweight standards for a Spanish population. Eur. J. Obstet. Gynecol. Reprod. Biol. 2008, 136, 20–24. [Google Scholar] [CrossRef]
- Bhide, A.; Acharya, G.; Bilardo, C.M.; Brezinka, C.; Cafici, D.; Hernandez-Andrade, E.; Kalache, K.; Kingdom, L.; Kiserud, T.; Lee, W.; et al. ISUOG Practice Guidelines: Use of Doppler ultrasonography in obstetrics. Ultrasound Obstet. Gynecol. 2013, 41, 233–239. [Google Scholar] [CrossRef]
- Baschat, A.A.; Gembruch, U. The cerebroplacental Doppler ratio revisited. Ultrasound Obstet. Gynecol. 2003, 21, 124–127. [Google Scholar] [CrossRef] [PubMed]
- Tong, Z.; Cui, Q.; Wang, J.; Zhou, Y. TransmiR v2.0: An updated transcription factor-microRNA regulation database. Nucleic Acids Res. 2019, 47, D253–D258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A Software Environment for Integrated Models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Martin, A.; E Ochagavia, M.; Rabasa, L.C.; Miranda, J.; Fernandez-De-Cossio, J.; Bringas, R. BisoGenet: A new tool for gene network building, visualization and analysis. BMC Bioinform. 2010, 11, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bindea, G.; Mlecnik, B.; Hackl, H.; Charoentong, P.; Tosolini, M.; Kirilovsky, A.; Fridman, W.-H.; Pagès, F.; Trajanoski, Z.; Galon, J. ClueGO: A Cytoscape plug-in to decipher functionally grouped gene ontology and pathway annotation networks. Bioinformatics 2009, 25, 1091–1093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, E.Y.; Tan, C.M.; Kou, Y.; Duan, Q.; Wang, Z.; Meirelles, G.V.; Clark, N.R.; Ma’Ayan, A. Enrichr: Interactive and collaborative HTML5 gene list enrichment analysis tool. BMC Bioinform. 2013, 14, 128. [Google Scholar] [CrossRef] [Green Version]
- Walter, W.; Sánchez-Cabo, F.; Ricote, M. GOplot: An R package for visually combining expression data with functional analysis: Figure 1. Bioinformatics. 2015, 31, 2912–2914. [Google Scholar] [CrossRef] [PubMed]
- Auguie, B.; Antonov, A. gridExtra: Miscellaneous Functions for “Grid” Graphics. R package, Version 2.3.; Available online: https://CRAN.R-project.org/package=gridExtra (accessed on 13 May 2020).
- Gómez-Rubio, V. ggplot2-elegant graphics for data analysis. J. Stat. Softw. 2017, 77, 1–3. [Google Scholar] [CrossRef] [Green Version]
- Neuwirth, E.; RColorBrewer: ColorBrewer Palettes. R Package, Version 1.1-2; 2014. Available online: https://cran.r-project.org/web/packages/.10.RColorBrewer/index.html (accessed on 13 May 2020).
- Gu, Z.; Gu, L.; Eils, R.; Schlesner, M.; Brors, B. circlize implements and enhances circular visualization in R. Bioinformatics 2014, 30, 2811–2812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jassal, B.; Matthews, L.; Viteri, G.; Gong, C.; Lorente, P.; Fabregat, A.; Sidiropoulos, K.; Cook, J.; Gillespie, M.; Haw, R.; et al. The reactome pathway knowledgebase. Nucleic Acids Res. 2020, 48, D498–D503. [Google Scholar] [CrossRef]
- Pico, A.; Kelder, T.; van Iersel, M.; Hanspers, K.; Conklin, B.R.; Evelo, C. WikiPathways: Pathway Editing for the People. PLoS Biol. 2008, 6, e184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rouillard, A.D.; Gundersen, G.W.; Fernandez, N.F.; Wang, Z.; Monteiro, C.D.; McDermott, M.G.; Ma’Ayan, A. The harmonizome: A collection of processed datasets gathered to serve and mine knowledge about genes and proteins. Database 2016, 2016. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Expression Levels of the miRNAs in the Two Study Groups (Mean ± Standard Deviation) | |||
|---|---|---|---|
| MiRNAs | FGR | Normal | p-Value |
| miR-132-3p | 26.00 ± 33.78 | 1.08 ± 0.91 | 0.0002 |
| miR-185-5p | 1.80 ± 0.90 | 1.22 ± 0.95 | 0.03 |
| miR-25-3p | 1.76 ± 1.09 | 1.19 ± 0.82 | 0.05 |
| miR-148b-3p | 1.51 ± 0.80 | 1.11 ± 0.52 | 0.18 (NS) |
| miR-183-5p | 1.23 ± 0.61 | 1.27 ± 1.64 | 0.24 (NS) |
| miR-193b-5p | 1.53 ± 2.14 | 1.37 ± 1.55 | 0.18 (NS) |
| Pathway | Adj p-Value |
|---|---|
| Kegg 2021 Human | |
| Pathways in cancer | 6.709 × 10−27 |
| Bladder cancer | 1.085 × 10−23 |
| Prostate cancer | 1.085 × 10−23 |
| Cellular senescence | 1.932 × 10−22 |
| Human T-cell leukemia virus 1 infection | 5.516 × 10−22 |
| Pancreatic cancer | 6.030 × 10−21 |
| Glioma | 2.045 × 10−19 |
| Hepatitis B | 2.045 × 10−19 |
| Human cytomegalovirus infection | 3.192 × 10−19 |
| MicroRNAs in cancer | 1.116 × 10−18 |
| Reactome 2016 | |
| Cellular responses to stress Homo sapiens R-HSA-2262752 | 3.128 × 10−13 |
| Cellular Senescence Homo Sapiens R-HSA-2559583 | 7.922 × 10−13 |
| Signal Transduction Homo Sapiens R-HSA-162582 | 9.454 × 10−13 |
| Fc epsilon receptor (FCERI) signaling Homo sapiens R-HSA-2454202 | 4.211 × 10−11 |
| Signaling by NGF Homo sapiens R-HSA-166520 | 4.211 × 10−11 |
| Signaling by EGFR Homo sapiens R-HSA-177929 | 4.704 × 10−11 |
| NGF signaling via TRKA from the plasma membrane Homo sapiens R-HSA-187037 | 1.014 × 10−10 |
| Downstream signal transduction Homo sapiens R-HSA-186763 | 1.937 × 10−10 |
| Signaling by PDGF Homo sapiens R-HSA-186797 | 4.611 × 10−10 |
| Developmental Biology Homo Sapiens R-HSA-1266738 | 4.611 × 10−10 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Loscalzo, G.; Scheel, J.; Ibañez-Cabellos, J.S.; García-Lopez, E.; Gupta, S.; García-Gimenez, J.L.; Mena-Mollá, S.; Perales-Marín, A.; Morales-Roselló, J. Overexpression of microRNAs miR-25-3p, miR-185-5p and miR-132-3p in Late Onset Fetal Growth Restriction, Validation of Results and Study of the Biochemical Pathways Involved. Int. J. Mol. Sci. 2022, 23, 293. https://doi.org/10.3390/ijms23010293
Loscalzo G, Scheel J, Ibañez-Cabellos JS, García-Lopez E, Gupta S, García-Gimenez JL, Mena-Mollá S, Perales-Marín A, Morales-Roselló J. Overexpression of microRNAs miR-25-3p, miR-185-5p and miR-132-3p in Late Onset Fetal Growth Restriction, Validation of Results and Study of the Biochemical Pathways Involved. International Journal of Molecular Sciences. 2022; 23(1):293. https://doi.org/10.3390/ijms23010293
Chicago/Turabian StyleLoscalzo, Gabriela, Julia Scheel, José Santiago Ibañez-Cabellos, Eva García-Lopez, Shailendra Gupta, José Luis García-Gimenez, Salvador Mena-Mollá, Alfredo Perales-Marín, and José Morales-Roselló. 2022. "Overexpression of microRNAs miR-25-3p, miR-185-5p and miR-132-3p in Late Onset Fetal Growth Restriction, Validation of Results and Study of the Biochemical Pathways Involved" International Journal of Molecular Sciences 23, no. 1: 293. https://doi.org/10.3390/ijms23010293