CB1 Activity Drives the Selection of Navigational Strategies: A Behavioral and c-Fos Immunoreactivity Study

 , and
, and
Abstract
:1. Introduction
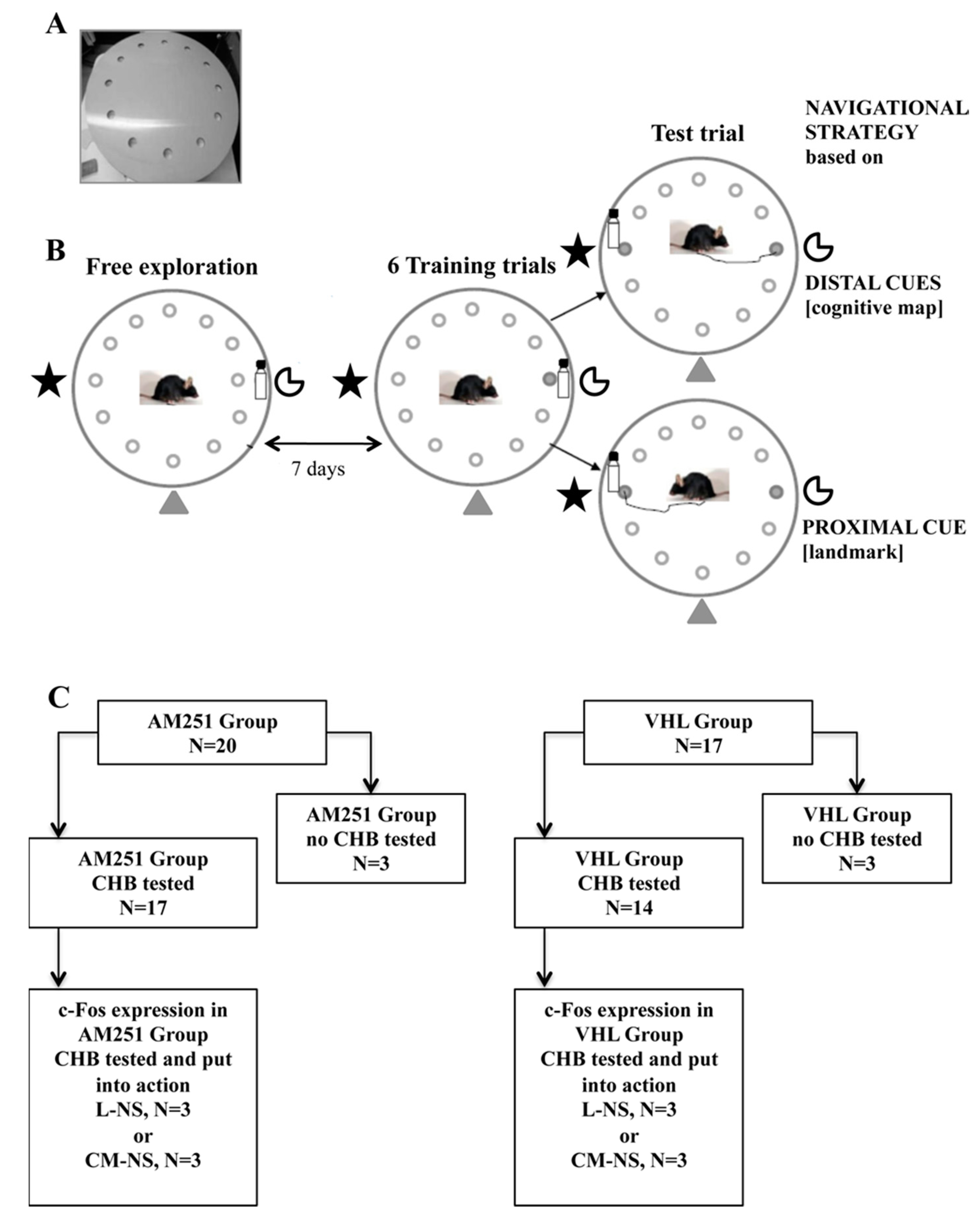
2. Results
2.1. CHB Behavioral Testing
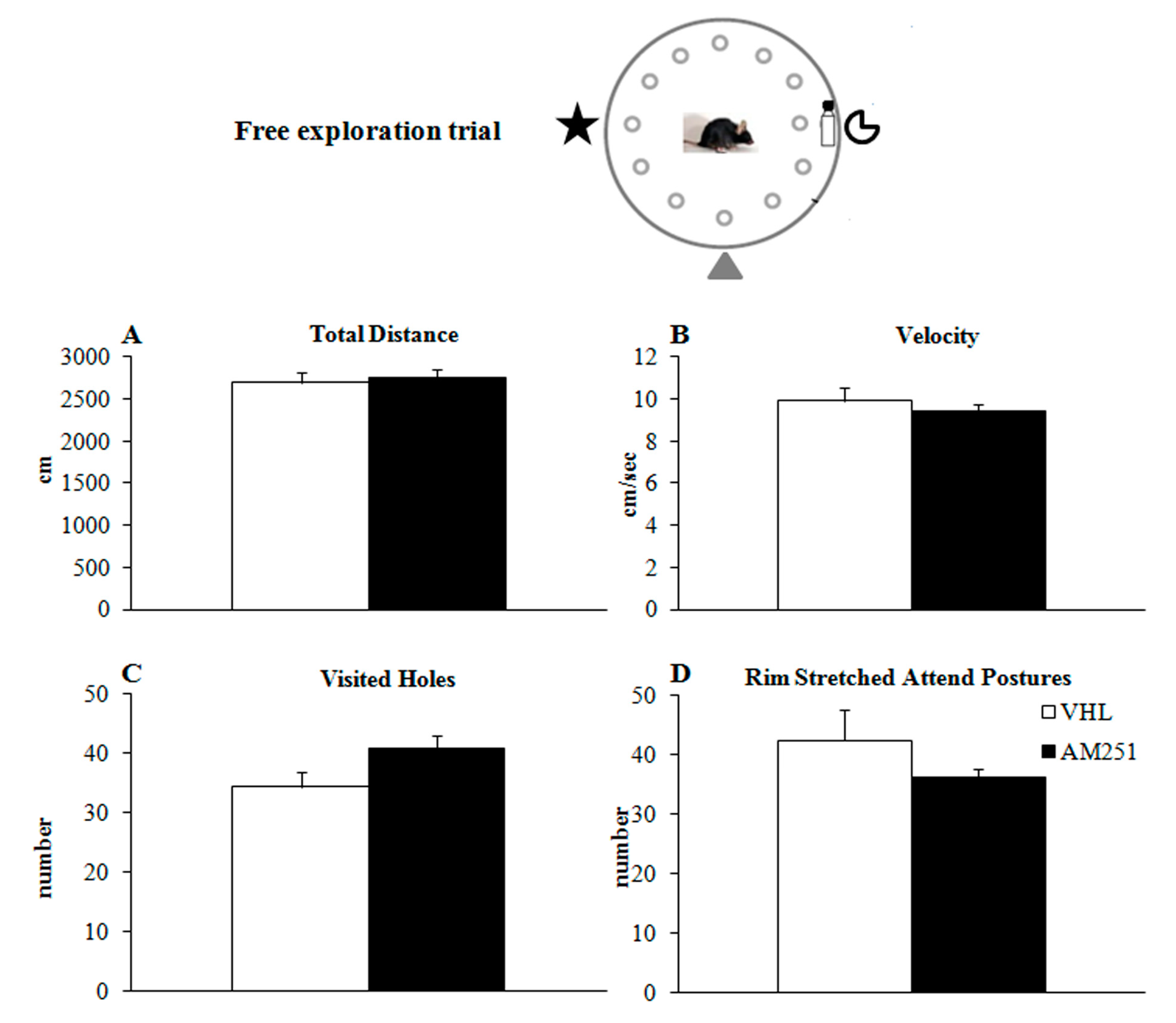
2.1.1. Free Exploration Trial
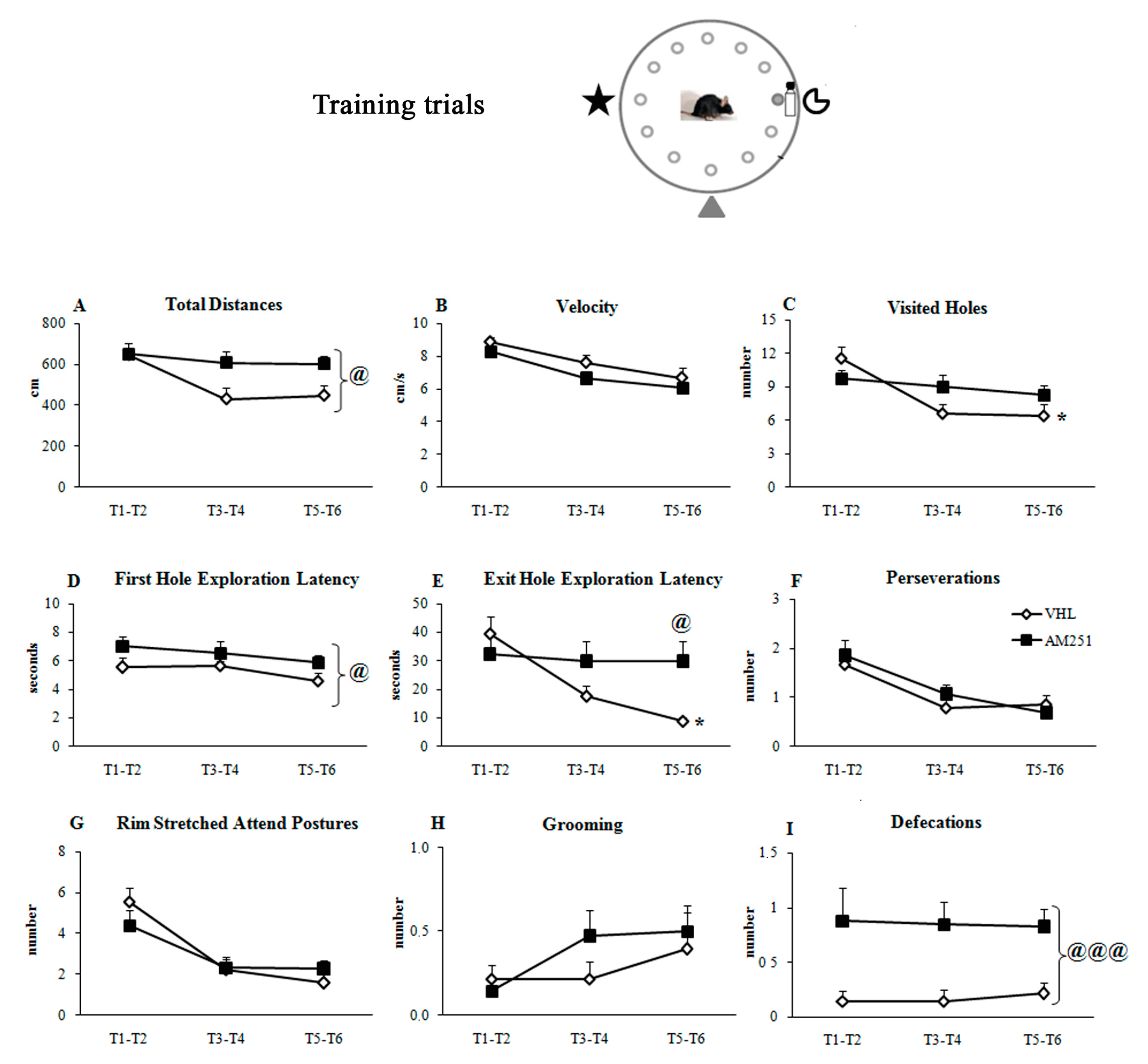
2.1.2. Training Trials
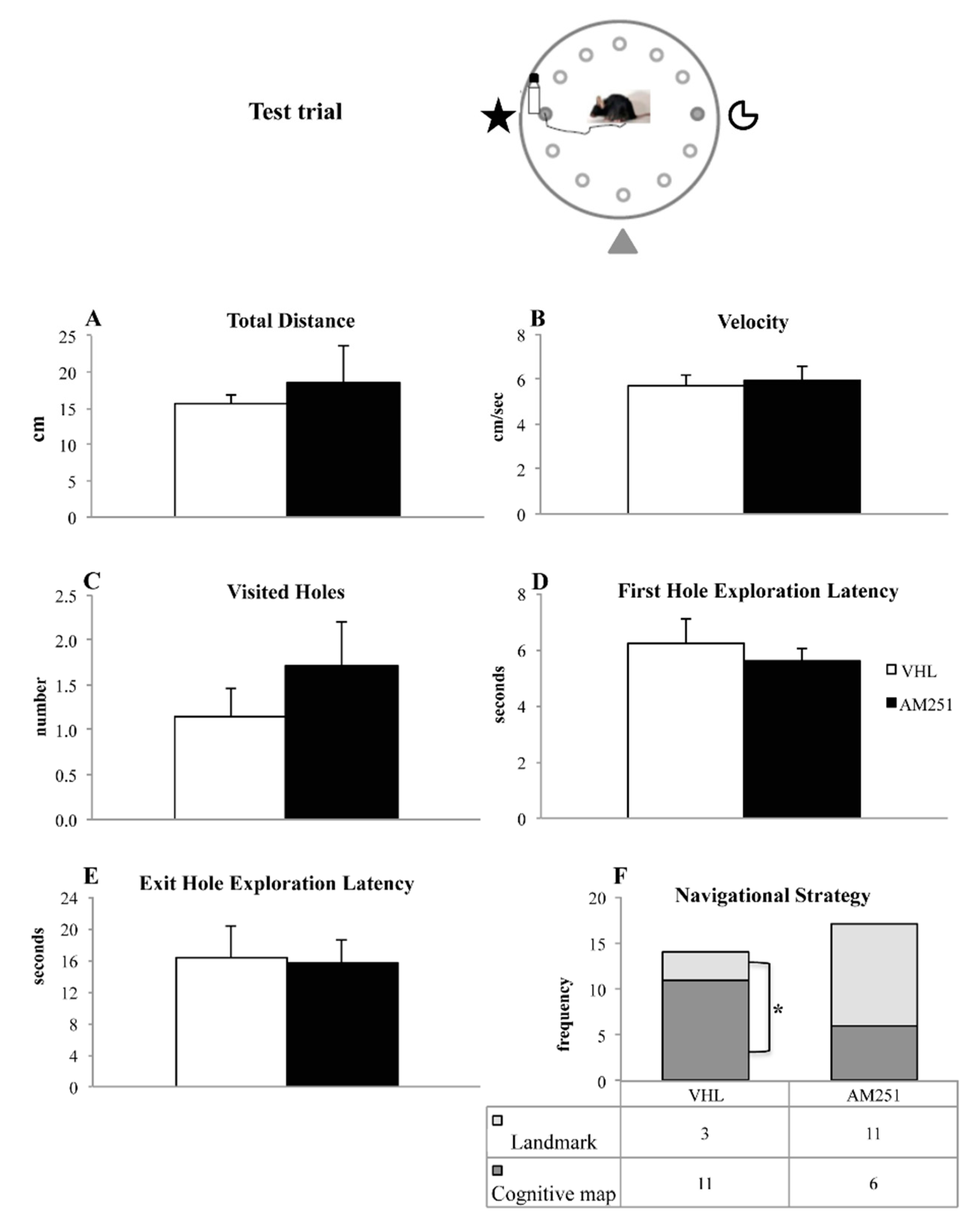
2.1.3. Test Trial
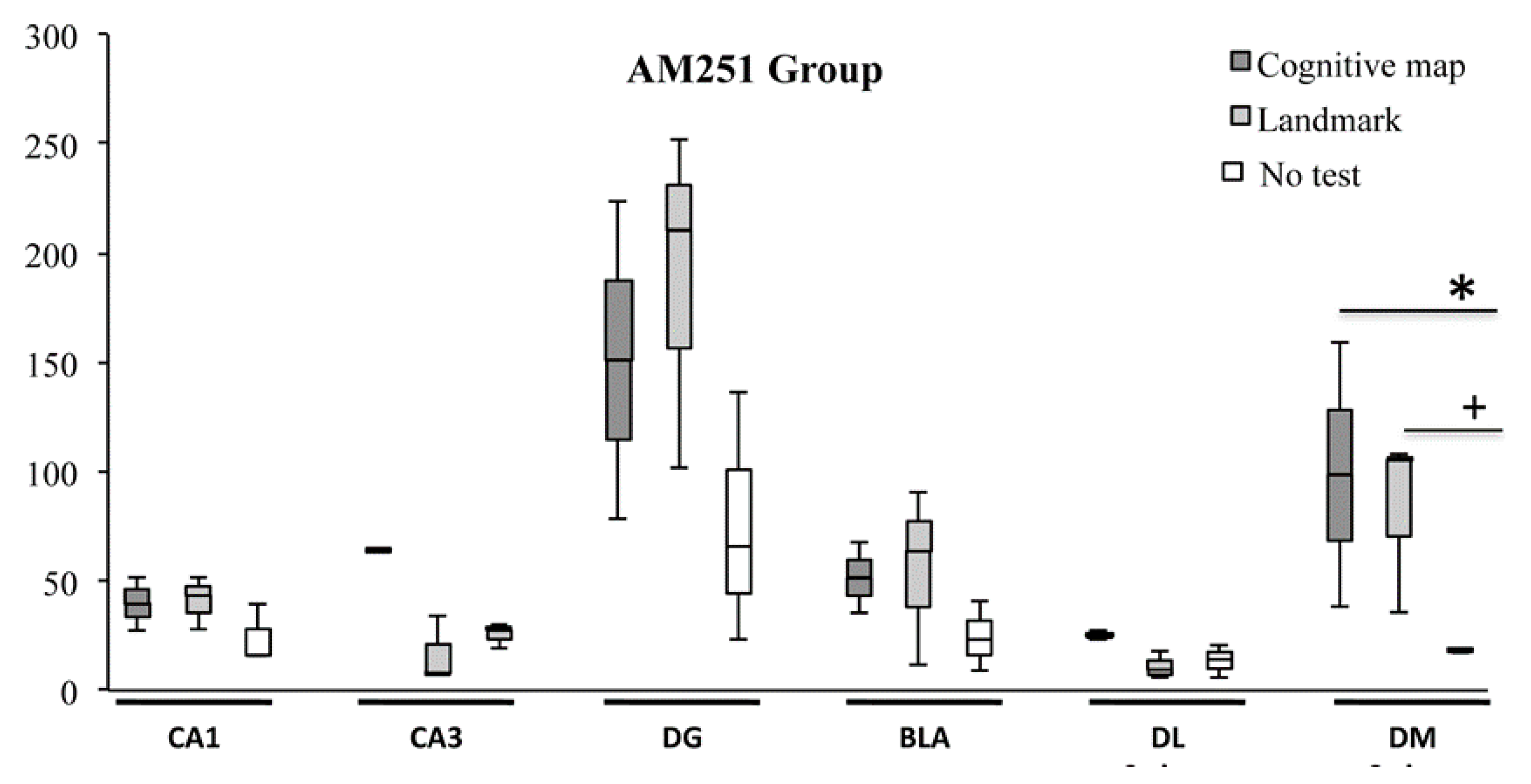
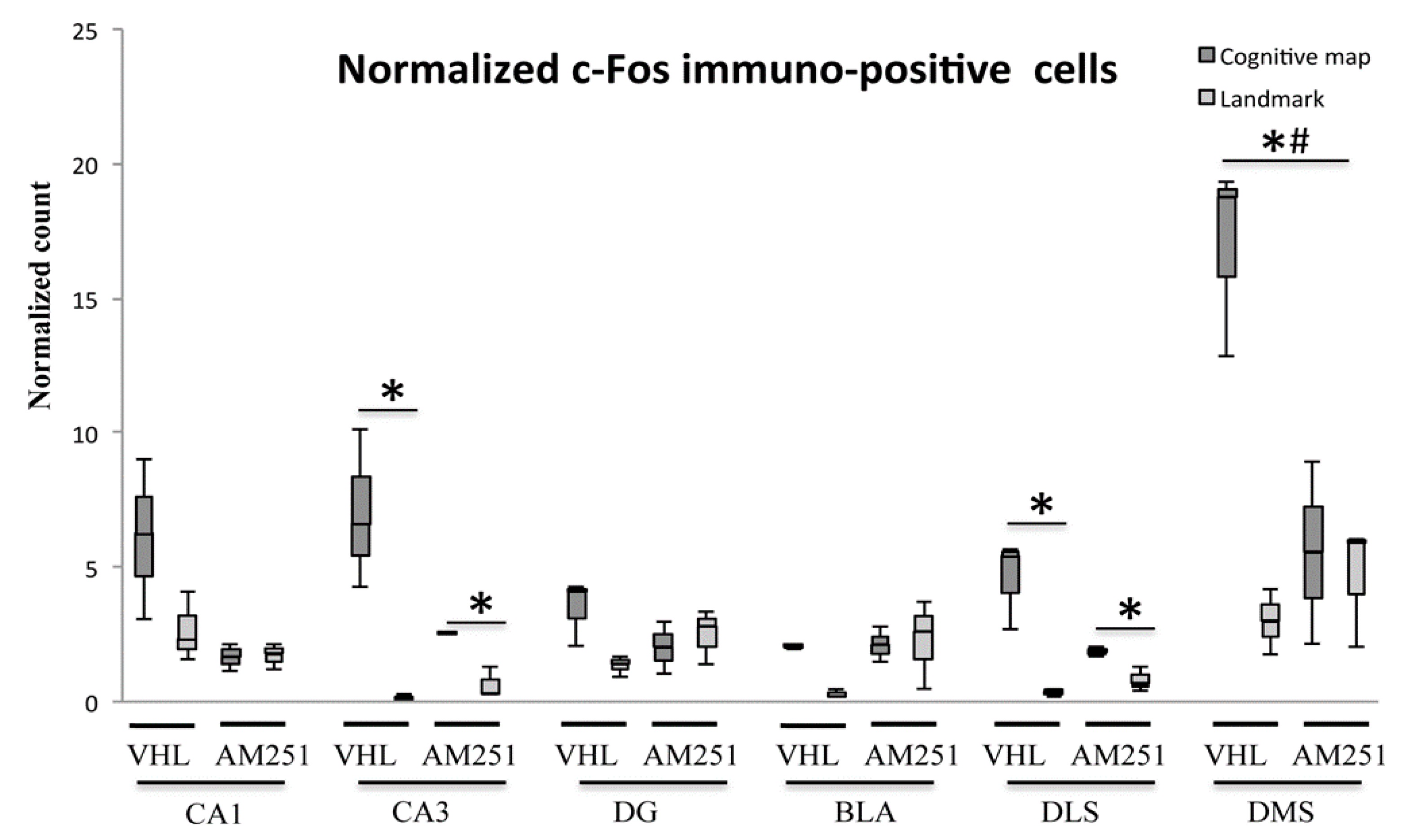
2.2. c-Fos Immunohistochemistry
3. Discussion
4. Material and Methods
4.1. Subjects
4.2. Drugs
4.3. CHB Testing
4.3.1. Apparatus
4.3.2. Procedures
4.3.3. Free Exploration Trial
4.3.4. Training Trials
4.3.5. Test Trial
4.4. Biochemical Analyses
4.4.1. Tissue Preparation
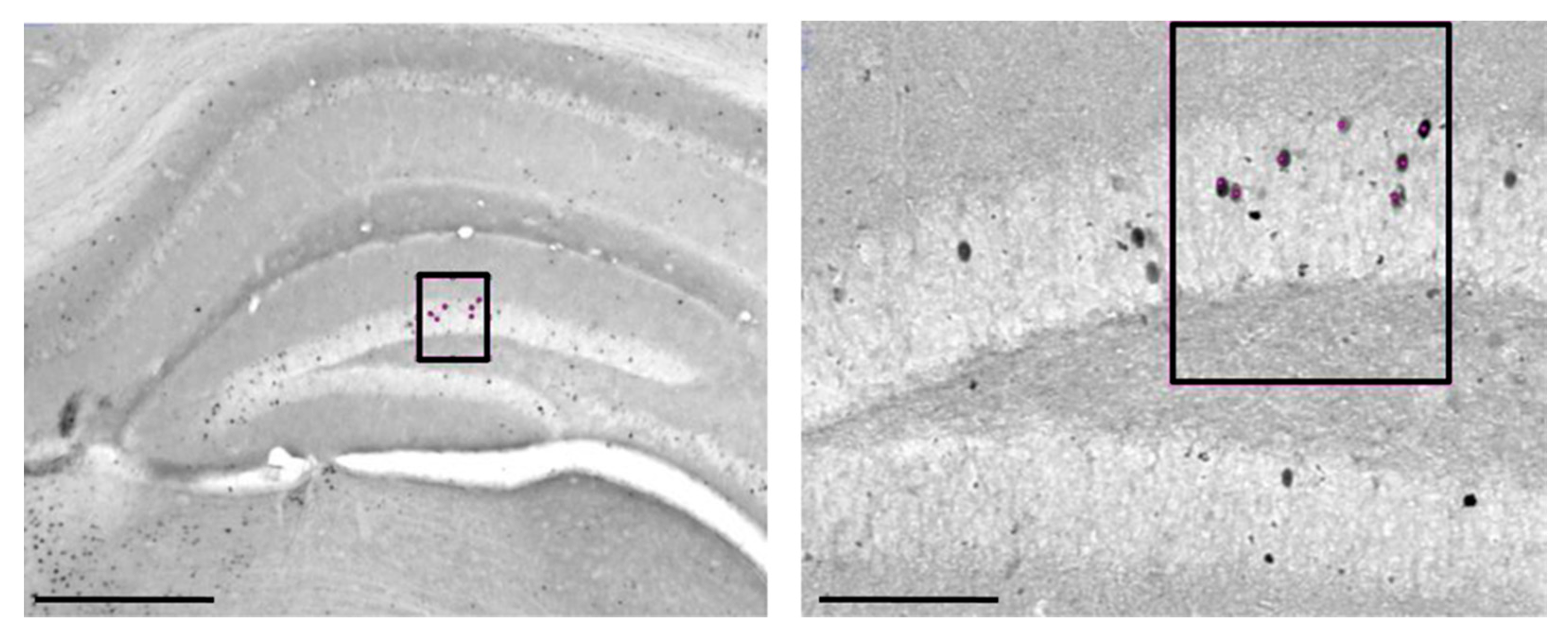
4.4.2. c-Fos Immunohistochemistry
4.4.3. Cell Counting
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Hinman, J.R.; Dannenberg, H.; Alexander, A.S.; Hasselmo, M.E. Neural mechanisms of navigation involving interactions of cortical and subcortical structures. J. Neurophysiol. 2018, 119, 2007–2029. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Wu, Y.; Liu, R.; Wang, J.; Huang, H.; Huang, R. Representation of human spatial navigation responding to input spatial information and output navigational strategies: An ALE meta-analysis. Neurosci. Biobehav. Rev. 2019, 103, 60–72. [Google Scholar] [CrossRef] [PubMed]
- Bocchi, A.; Palmiero, M.; Nori, R.; Verde, P.; Piccardi, L. Does spatial cognitive style affect how navigational strategy is planned? Exp. Brain Res. 2019, 237, 2523–2533. [Google Scholar] [CrossRef] [PubMed]
- Ekstrom, A.D.; Isham, E.A. Human spatial navigation: Representations across dimensions and scales. Curr. Opin. Behav. Sci. 2017, 17, 84–89. [Google Scholar] [CrossRef]
- Olson, C.R. Brain representation of object-centered space in monkeys and humans. Annu. Rev. Neurosci. 2003, 26, 331–354. [Google Scholar] [CrossRef] [Green Version]
- Siegel, A.W.; White, S.H. The development of spatial representations of large-scale environments. Adv. Child. Dev. Behav. 1975, 10, 9–55. [Google Scholar]
- Dezfouli, A.; Lingawi, N.W.; Balleine, B.W. Habits as action sequences: Hierarchical action control and changes in outcome value. Philos. Trans. R. Soc. B Biol. Sci. 2014, 369, 20130482. [Google Scholar] [CrossRef]
- Graybiel, A.M. Habits, Rituals, and the Evaluative Brain. Annu. Rev. Neurosci. 2008, 31, 359–387. [Google Scholar] [CrossRef] [Green Version]
- Bonicalzi, S.; Haggard, P. From Freedom from to Freedom to: New Perspectives on Intentional Action. Front. Psychol. 2019, 10, 1193. [Google Scholar] [CrossRef] [Green Version]
- Ruotolo, F.; Iachini, T.; Ruggiero, G.; van der Ham, I.J.M.; Postma, A. Frames of reference and categorical/coordinate spatial relations in a “what was where” task. Exp. Brain Res. 2016, 234, 2687–2696. [Google Scholar] [CrossRef] [Green Version]
- Eichenbaum, H. Time (and space) in the hippocampus. Curr. Opin. Behav. Sci. 2017, 17, 65–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dolan, R.J.; Dayan, P. Goals and Habits in the Brain. Neuron 2013, 80, 312–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elliott, A.E.; Packard, M.G. Intra-amygdala anxiogenic drug infusion prior to retrieval biases rats towards the use of habit memory. Neurobiol. Learn. Mem. 2008, 90, 616–623. [Google Scholar] [CrossRef] [PubMed]
- Schwabe, L.; Wolf, O.T. Stress Modulates the Engagement of Multiple Memory Systems in Classification Learning. J. Neurosci. 2012, 32, 11042–11049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malvaez, M.; Wassum, K.M. Regulation of habit formation in the dorsal striatum. Curr. Opin. Behav. Sci. 2018, 20, 67–74. [Google Scholar] [CrossRef]
- Packard, M.G.; Knowlton, B.J. Learning and Memory Functions of the Basal Ganglia. Annu. Rev. Neurosci. 2002, 25, 563–593. [Google Scholar] [CrossRef] [Green Version]
- Yin, H.H.; Knowlton, B.J.; Balleine, B.W. Lesions of dorsolateral striatum preserve outcome expectancy but disrupt habit formation in instrumental learning. Eur. J. Neurosci. 2004, 19, 181–189. [Google Scholar] [CrossRef] [PubMed]
- White, N.M.; Packard, M.G.; McDonald, R.J. Dissociation of memory systems: The story unfolds. Behav. Neurosci. 2013, 127, 813–834. [Google Scholar] [CrossRef] [PubMed]
- O’Keefe, J.; Nadel, L. The Hippocampus AS a Cognitive Map; Clarendon Press: Oxford, UK, 1978. [Google Scholar]
- Morris, R.G.M.; Garrud, P.; Rawlins, J.N.P.; O’Keefe, J. Place navigation impaired in rats with hippocampal lesions. Nature 1982, 297, 681–683. [Google Scholar] [CrossRef] [PubMed]
- Rudy, J.W.; Sutherland, R.J. Configural association theory and the hippocampal formation: An appraisal and reconfiguration. Hippocampus 1995, 5, 375–389. [Google Scholar] [CrossRef]
- Mizumori, S.J.Y.; Yeshenko, O.; Gill, K.M.; Davis, D.M. Parallel processing across neural systems: Implications for a multiple memory system hypothesis. Neurobiol. Learn. Mem. 2004, 82, 278–298. [Google Scholar] [CrossRef]
- Yeshenko, O.; Guazzelli, A.; Mizumori, S.J.Y. Context-dependent reorganization of spatial and movement representations by simultaneously recorded hippocampal and striatal neurons during performance of allocentric and egocentric tasks. Behav. Neurosci. 2004, 118, 751–769. [Google Scholar] [CrossRef] [Green Version]
- Deoliveiraalvares, L.; Deoliveira, L.; Camboim, C.; Diehl, F.; Genro, B.; Lanziotti, V.; Quillfeldt, J. Amnestic effect of intrahippocampal AM251, a CB1-selective blocker, in the inhibitory avoidance, but not in the open field habituation task, in rats. Neurobiol. Learn. Mem. 2005, 83, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Katona, I.; Rancz, E.A.; Acsady, L.; Ledent, C.; Mackie, K.; Hajos, N.; Freund, T.F. Distribution of CB1 cannabinoid receptors in the amygdala and their role in the control of GABAergic transmission. J. Neurosci. Off. J. Soc. Neurosci. 2001, 21, 9506–9518. [Google Scholar] [CrossRef]
- Laricchiuta, D.; Centonze, D.; Petrosini, L. Effects of endocannabinoid and endovanilloid systems on aversive memory extinction. Behav. Brain Res. 2013, 256, 101–107. [Google Scholar] [CrossRef]
- Laricchiuta, D.; Saba, L.; De Bartolo, P.; Caioli, S.; Zona, C.; Petrosini, L. Maintenance of aversive memories shown by fear extinction-impaired phenotypes is associated with increased activity in the amygdaloid-prefrontal circuit. Sci. Rep. 2016, 6, 21205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peterfi, Z.; Urban, G.M.; Papp, O.I.; Nemeth, B.; Monyer, H.; Szabo, G.; Erdelyi, F.; Mackie, K.; Freund, T.F.; Hajos, N.; et al. Endocannabinoid-Mediated Long-Term Depression of Afferent Excitatory Synapses in Hippocampal Pyramidal Cells and GABAergic Interneurons. J. Neurosci. 2012, 32, 14448–14463. [Google Scholar] [CrossRef] [PubMed]
- Katona, I.; Freund, T.F. Multiple functions of endocannabinoid signaling in the brain. Annu. Rev. Neurosci. 2012, 35, 529–558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marsicano, G.; Lutz, B. Expression of the cannabinoid receptor CB1 in distinct neuronal subpopulations in the adult mouse forebrain: CB1 expression in murine forebrain. Eur. J. Neurosci. 1999, 11, 4213–4225. [Google Scholar] [CrossRef] [PubMed]
- Katona, I. Endocannabinoid receptors: CNS localization of the CB1 cannabinoid receptor. Curr. Top. Behav. Neurosci. 2009, 1, 65–86. [Google Scholar]
- Piomelli, D. The molecular logic of endocannabinoid signalling. Nat. Rev. Neurosci. 2003, 4, 873–884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Marzo, V.; Fontana, A.; Cadas, H.; Schinelli, S.; Cimino, G.; Schwartz, J.-C.; Piomelli, D. Formation and inactivation of endogenous cannabinoid anandamide in central neurons. Nature 1994, 372, 686–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campos, A.C.; Ferreira, F.R.; Guimarães, F.S.; Lemos, J.I. Facilitation of endocannabinoid effects in the ventral hippocampus modulates anxiety-like behaviors depending on previous stress experience. Neuroscience 2010, 167, 238–246. [Google Scholar] [CrossRef] [PubMed]
- Hill, M.N.; Patel, S.; Carrier, E.J.; Rademacher, D.J.; Ormerod, B.K.; Hillard, C.J.; Gorzalka, B.B. Downregulation of Endocannabinoid Signaling in the Hippocampus Following Chronic Unpredictable Stress. Neuropsychopharmacology 2005, 30, 508–515. [Google Scholar] [CrossRef] [PubMed]
- Pietrzak, R.H.; Huang, Y.; Corsi-Travali, S.; Zheng, M.-Q.; Lin, S.; Henry, S.; Potenza, M.N.; Piomelli, D.; Carson, R.E.; Neumeister, A. Cannabinoid Type 1 Receptor Availability in the Amygdala Mediates Threat Processing in Trauma Survivors. Neuropsychopharmacology 2014, 39, 2519–2528. [Google Scholar] [CrossRef] [PubMed]
- De Chiara, V.; Angelucci, F.; Rossi, S.; Musella, A.; Cavasinni, F.; Cantarella, C.; Mataluni, G.; Sacchetti, L.; Napolitano, F.; Castelli, M.; et al. Brain-Derived Neurotrophic Factor Controls Cannabinoid CB1 Receptor Function in the Striatum. J. Neurosci. 2010, 30, 8127–8137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sciolino, N.R.; Bortolato, M.; Eisenstein, S.A.; Fu, J.; Oveisi, F.; Hohmann, A.G.; Piomelli, D. Social isolation and chronic handling alter endocannabinoid signaling and behavioral reactivity to context in adult rats. Neuroscience 2010, 168, 371–386. [Google Scholar] [CrossRef] [Green Version]
- Abush, H.; Akirav, I. Cannabinoids modulate hippocampal memory and plasticity. Hippocampus 2010, 20, 1126–1138. [Google Scholar] [CrossRef]
- Galanopoulos, A.; Polissidis, A.; Georgiadou, G.; Papadopoulou-Daifoti, Z.; Nomikos, G.G.; Pitsikas, N.; Antoniou, K. WIN55,212-2 impairs non-associative recognition and spatial memory in rats via CB1 receptor stimulation. Pharmacol. Biochem. Behav. 2014, 124, 58–66. [Google Scholar] [CrossRef]
- Rueda-Orozco, P.E.; Soria-Gomez, E.; Montes-Rodriguez, C.J.; Martínez-Vargas, M.; Galicia, O.; Navarro, L.; Prospero-García, O. A potential function of endocannabinoids in the selection of a navigation strategy by rats. Psychopharmacology (Berl.) 2008, 198, 565–576. [Google Scholar] [CrossRef]
- Kruk-Slomka, M.; Biala, G. CB1 receptors in the formation of the different phases of memory-related processes in the inhibitory avoidance test in mice. Behav. Brain Res. 2016, 301, 84–95. [Google Scholar] [CrossRef] [PubMed]
- Lichtman, A.H. SR 141716A enhances spatial memory as assessed in a radial-arm maze task in rats. Eur. J. Pharmacol. 2000, 404, 175–179. [Google Scholar] [CrossRef]
- Wise, L.E.; Shelton, C.C.; Cravatt, B.F.; Martin, B.R.; Lichtman, A.H. Assessment of anandamide’s pharmacological effects in mice deficient of both fatty acid amide hydrolase and cannabinoid CB1 receptors. Eur. J. Pharmacol. 2007, 557, 44–48. [Google Scholar] [CrossRef] [PubMed]
- Wolff, M.C.; Leander, J.D. SR141716A, a cannabinoid CB1 receptor antagonist, improves memory in a delayed radial maze task. Eur. J. Pharmacol. 2003, 477, 213–217. [Google Scholar] [CrossRef] [PubMed]
- Assini, F.L.; Nakamura, C.A.; Piermartiri, T.C.; Tasca, C.I.; Takahashi, R.N. Coadministration of cannabinoid CB1-receptor and adenosine A1-receptor antagonists improves the acquisition of spatial memory in mice: Participation of glutamatergic neurotransmission. Behav. Pharmacol. 2012, 23, 292–301. [Google Scholar] [CrossRef] [PubMed]
- Varvel, S.A.; Lichtman, A.H. Evaluation of CB1 Receptor Knockout Mice in the Morris Water Maze. J. Pharmacol. Exp. Ther. 2002, 301, 915–924. [Google Scholar] [CrossRef]
- Robinson, L.; McKillop-Smith, S.; Ross, N.L.; Pertwee, R.G.; Hampson, R.E.; Platt, B.; Riedel, G. Hippocampal endocannabinoids inhibit spatial learning and limit spatial memory in rats. Psychopharmacology (Berl.) 2008, 198, 551–563. [Google Scholar] [CrossRef]
- Morena, M.; Campolongo, P. The endocannabinoid system: An emotional buffer in the modulation of memory function. Neurobiol. Learn. Mem. 2014, 112, 30–43. [Google Scholar] [CrossRef]
- Darmani, N.A. Methods Evaluating Cannabinoid and Endocannabinoid Effects on Gastrointestinal Functions. In Marijuana and Cannabinoid Research; Humana Press: Totowa, NJ, USA, 2005; Volume 123, pp. 169–189. ISBN 978-1-59259-999-8. [Google Scholar]
- Fouquet, C.; Babayan, B.M.; Watilliaux, A.; Bontempi, B.; Tobin, C.; Rondi-Reig, L. Complementary Roles of the Hippocampus and the Dorsomedial Striatum during Spatial and Sequence-Based Navigation Behavior. PLoS ONE 2013, 8, e67232. [Google Scholar] [CrossRef] [Green Version]
- Berg, K.A.; Clarke, W.P. Making Sense of Pharmacology: Inverse Agonism and Functional Selectivity. Int. J. Neuropsychopharmacol. 2018, 21, 962–977. [Google Scholar] [CrossRef] [Green Version]
- Pertwee, R.G. Inverse agonism and neutral antagonism at cannabinoid CB1 receptors. Life Sci. 2005, 76, 1307–1324. [Google Scholar] [CrossRef] [PubMed]
- Sim-Selley, L.J.; Brunk, L.K.; Selley, D.E. Inhibitory effects of SR141716A on G-protein activation in rat brain. Eur. J. Pharmacol. 2001, 414, 135–143. [Google Scholar] [CrossRef]
- Barbieri, M.; Ossato, A.; Canazza, I.; Trapella, C.; Borelli, A.C.; Beggiato, S.; Rimondo, C.; Serpelloni, G.; Ferraro, L.; Marti, M. Synthetic cannabinoid JWH-018 and its halogenated derivatives JWH-018-Cl and JWH-018-Br impair Novel Object Recognition in mice: Behavioral, electrophysiological and neurochemical evidence. Neuropharmacology 2016, 109, 254–269. [Google Scholar] [CrossRef] [PubMed]
- Maione, S.; Costa, B.; Piscitelli, F.; Morera, E.; De Chiaro, M.; Comelli, F.; Boccella, S.; Guida, F.; Verde, R.; Ortar, G.; et al. Piperazinyl carbamate fatty acid amide hydrolase inhibitors and transient receptor potential channel modulators as “dual-target” analgesics. Pharmacol. Res. 2013, 76, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Newsom, R.J.; Garcia, R.J.; Stafford, J.; Osterlund, C.; O’Neill, C.E.; Day, H.E.W.; Campeau, S. Remote CB1 receptor antagonist administration reveals multiple sites of tonic and phasic endocannabinoid neuroendocrine regulation. Psychoneuroendocrinology 2020, 113, 104549. [Google Scholar] [CrossRef] [PubMed]
- McLaughlin, P.J.; Winston, K.M.; Swezey, L.A.; Vemuri, V.K.; Makriyannis, A.; Salamone, J.D. Detailed analysis of food-reinforced operant lever pressing distinguishes effects of a cannabinoid CB1 inverse agonist and dopamine D1 and D2 antagonists. Pharmacol. Biochem. Behav. 2010, 96, 75–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, M.N.; McLaughlin, R.J.; Bingham, B.; Shrestha, L.; Lee, T.T.Y.; Gray, J.M.; Hillard, C.J.; Gorzalka, B.B.; Viau, V. Endogenous cannabinoid signaling is essential for stress adaptation. Proc. Natl. Acad. Sci. USA 2010, 107, 9406–9411. [Google Scholar] [CrossRef] [Green Version]
- Henstridge, C.M.; Balenga, N.A.B.; Ford, L.A.; Ross, R.A.; Waldhoer, M.; Irving, A.J. The GPR55 ligand L-α-lysophosphatidylinositol promotes RhoA-dependent Ca 2+ signaling and NFAT activation. FASEB J. 2009, 23, 183–193. [Google Scholar] [CrossRef]
- Ruz-Maldonado, I.; Liu, B.; Atanes, P.; Pingitore, A.; Huang, G.C.; Choudhary, P.; Persaud, S.J. The cannabinoid ligands SR141716A and AM251 enhance human and mouse islet function via GPR55-independent signalling. Cell. Mol. Life Sci. 2020. [Google Scholar] [CrossRef] [Green Version]
- Ryberg, E.; Larsson, N.; Sjögren, S.; Hjorth, S.; Hermansson, N.-O.; Leonova, J.; Elebring, T.; Nilsson, K.; Drmota, T.; Greasley, P.J. The orphan receptor GPR55 is a novel cannabinoid receptor. Br. J. Pharmacol. 2007, 152, 1092–1101. [Google Scholar] [CrossRef]
- Walsh, S.K.; Hepburn, C.Y.; Keown, O.; Åstrand, A.; Lindblom, A.; Ryberg, E.; Hjorth, S.; Leslie, S.J.; Greasley, P.J.; Wainwright, C.L. Pharmacological profiling of the hemodynamic effects of cannabinoid ligands: A combined in vitro and in vivo approach. Pharmacol. Res. Perspect. 2015, 3, e00143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tudurí, E.; Imbernon, M.; Hernández-Bautista, R.J.; Tojo, M.; Fernø, J.; Diéguez, C.; Nogueiras, R. GPR55: A new promising target for metabolism? J. Mol. Endocrinol. 2017, 58, R191–R202. [Google Scholar] [CrossRef] [PubMed]
- Pertwee, R.G.; Howlett, A.C.; Abood, M.E.; Alexander, S.P.H.; Di Marzo, V.; Elphick, M.R.; Greasley, P.J.; Hansen, H.S.; Kunos, G.; Mackie, K.; et al. International Union of Basic and Clinical Pharmacology. LXXIX. Cannabinoid Receptors and Their Ligands: Beyond CB1 and CB2. Pharmacol. Rev. 2010, 62, 588–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marichal-Cancino, B.A.; Sánchez-Fuentes, A.; Méndez-Díaz, M.; Ruiz-Contreras, A.E.; Prospéro-García, O. Blockade of GPR55 in the dorsolateral striatum impairs performance of rats in a T-maze paradigm. Behav. Pharmacol. 2016, 27, 393–396. [Google Scholar] [CrossRef]
- Marichal-Cancino, B.A.; Fajardo-Valdez, A.; Ruiz-Contreras, A.E.; Méndez-Díaz, M.; Prospéro-García, O. Possible role of hippocampal GPR55 in spatial learning and memory in rats. Acta Neurobiol. Exp. 2018, 78, 41–50. [Google Scholar] [CrossRef] [Green Version]
- Kramar, C.; Loureiro, M.; Renard, J.; Laviolette, S.R. Palmitoylethanolamide Modulates GPR55 Receptor Signaling in the Ventral Hippocampus to Regulate Mesolimbic Dopamine Activity, Social Interaction, and Memory Processing. Cannabis Cannabinoid Res. 2017, 2, 8–20. [Google Scholar] [CrossRef]
- Hurst, K.; Badgley, C.; Ellsworth, T.; Bell, S.; Friend, L.; Prince, B.; Welch, J.; Cowan, Z.; Williamson, R.; Lyon, C.; et al. A putative lysophosphatidylinositol receptor GPR55 modulates hippocampal synaptic plasticity. Hippocampus 2017, 27, 985–998. [Google Scholar] [CrossRef]
- Rueda-Orozco, P.E.; Montes-Rodriguez, C.J.; Ruiz-Contreras, A.E.; Mendez-Diaz, M.; Prospero-Garcia, O. The effects of anandamide and oleamide on cognition depend on diurnal variations. Brain Res. 2017, 1672, 129–136. [Google Scholar] [CrossRef]
- Ragozzino, M.E. The Contribution of the Medial Prefrontal Cortex, Orbitofrontal Cortex, and Dorsomedial Striatum to Behavioral Flexibility. Ann. N. Y. Acad. Sci. 2007, 1121, 355–375. [Google Scholar] [CrossRef] [Green Version]
- Peak, J.; Hart, G.; Balleine, B.W. From learning to action: The integration of dorsal striatal input and output pathways in instrumental conditioning. Eur. J. Neurosci. 2019, 49, 658–671. [Google Scholar] [CrossRef]
- Moussa, R.; Poucet, B.; Amalric, M.; Sargolini, F. Contributions of dorsal striatal subregions to spatial alternation behavior. Learn. Mem. 2011, 18, 444–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hart, G.; Bradfield, L.A.; Balleine, B.W. Prefrontal Corticostriatal Disconnection Blocks the Acquisition of Goal-Directed Action. J. Neurosci. 2018, 38, 1311–1322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiener, S.I. Spatial and behavioral correlates of striatal neurons in rats performing a self-initiated navigation task. J. Neurosci. Off. J. Soc. Neurosci. 1993, 13, 3802–3817. [Google Scholar] [CrossRef]
- Wiener, S.I. Spatial, behavioral and sensory correlates of hippocampal CA1 complex spike cell activity: Implications for information processing functions. Prog. Neurobiol. 1996, 49, 335–361. [Google Scholar] [CrossRef]
- Schmitzer-Torbert, N.; Redish, A.D. Neuronal Activity in the Rodent Dorsal Striatum in Sequential Navigation: Separation of Spatial and Reward Responses on the Multiple T Task. J. Neurophysiol. 2004, 91, 2259–2272. [Google Scholar] [CrossRef] [Green Version]
- Khamassi, M.; Humphries, M.D. Integrating cortico-limbic-basal ganglia architectures for learning model-based and model-free navigation strategies. Front. Behav. Neurosci. 2012, 6, 79. [Google Scholar] [CrossRef] [Green Version]
- Hunnicutt, B.J.; Jongbloets, B.C.; Birdsong, W.T.; Gertz, K.J.; Zhong, H.; Mao, T. A comprehensive excitatory input map of the striatum reveals novel functional organization. eLife 2016, 5. [Google Scholar] [CrossRef]
- Hintiryan, H.; Foster, N.N.; Bowman, I.; Bay, M.; Song, M.Y.; Gou, L.; Yamashita, S.; Bienkowski, M.S.; Zingg, B.; Zhu, M.; et al. The mouse cortico-striatal projectome. Nat. Neurosci. 2016, 19, 1100–1114. [Google Scholar] [CrossRef]
- Steiner, H.; Tseng, K.-Y. Handbook of Basal Ganglia Structure and Function, 2nd ed.; Academic Press: Cambridge, MA, USA, 2016. [Google Scholar]
- Fernández-Cabrera, M.R.; Higuera-Matas, A.; Fernaud-Espinosa, I.; DeFelipe, J.; Ambrosio, E.; Miguéns, M. Selective effects of Δ9-tetrahydrocannabinol on medium spiny neurons in the striatum. PLoS ONE 2018, 13, e0200950. [Google Scholar] [CrossRef]
- Huang, C.C.; Lo, S.W.; Hsu, K.S. Presynaptic mechanisms underlying cannabinoid inhibition of excitatory synaptic transmission in rat striatal neurons. J. Physiol. 2001, 532, 731–748. [Google Scholar] [CrossRef]
- Köfalvi, A.; Rodrigues, R.J.; Ledent, C.; Mackie, K.; Vizi, E.S.; Cunha, R.A.; Sperlágh, B. Involvement of cannabinoid receptors in the regulation of neurotransmitter release in the rodent striatum: A combined immunochemical and pharmacological analysis. J. Neurosci. Off. J. Soc. Neurosci. 2005, 25, 2874–2884. [Google Scholar] [CrossRef] [PubMed]
- Eisenstein, S.A.; Clapper, J.R.; Holmes, P.V.; Piomelli, D.; Hohmann, A.G. A role for 2-arachidonoylglycerol and endocannabinoid signaling in the locomotor response to novelty induced by olfactory bulbectomy. Pharmacol. Res. 2010, 61, 419–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Umathe, S.N.; Manna, S.S.S.; Utturwar, K.S.; Jain, N.S. Endocannabinoids mediate anxiolytic-like effect of acetaminophen via CB1 receptors. Prog. Neuropsychopharmacol. Biol. Psychiatry 2009, 33, 1191–1199. [Google Scholar] [CrossRef] [PubMed]
- Laricchiuta, D.; Musella, A.; Rossi, S.; Centonze, D. Behavioral and electrophysiological effects of endocannabinoid and dopaminergic systems on salient stimuli. Front. Behav. Neurosci. 2014, 8, 183. [Google Scholar] [CrossRef] [Green Version]
- Conversi, D.; Orsini, C.; Cabib, S. Distinct patterns of Fos expression induced by systemic amphetamine in the striatal complex of C57BL/6JICo and DBA/2JICo inbred strains of mice. Brain Res. 2004, 1025, 59–66. [Google Scholar] [CrossRef]
- Paxinos, G.; Franklin, K.B.J. Paxinos and Franklin’s the Mouse Brain in Stereotaxic Coordinates, 5th ed.; Academic Press: Cambridge, MA, USA, 2019. [Google Scholar]
- Albouy, G.; King, B.R.; Maquet, P.; Doyon, J. Hippocampus and striatum: Dynamics and interaction during acquisition and sleep-related motor sequence memory consolidation. Hippocampus 2013, 23, 985–1004. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Effect (Freedom Degrees) | Total Distances | Velocity | Visited Holes | First Hole Latency | Exit Hole Latency | Perseverations | Rim Stretched Attend Postures | Grooming | Defecations |
|---|---|---|---|---|---|---|---|---|---|
| Group F(1,29) p | 4.01 0.05 * | 2.42 0.13 | 0.80 0.37 | 6.27 0.02 * | 2.42 0.13 | 1.16 0.69 | 0.05 0.83 | 0.66 0.42 | 16.29 0.0004 * |
| Trials F(2,29) p | 6.40 0.003 * | 25.67 ≤0.0001 * | 10.50 0.0001 * | 1.65 0.20 | 5.20 0.008 * | 14.42 ≤0.0001 * | 21.17 ≤0.0001 * | 1.92 0.15 | 0.01 0.99 |
| Interaction F(2,58) p | 2.63 0.08 | 0.26 0.77 | 4.20 0.02 * | 0.11 0.89 | 3.70 0.03 * | 0.71 0.49 | 1.64 0.20 | 0.70 0.50 | 0.07 0.93 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Laricchiuta, D.; Balsamo, F.; Fabrizio, C.; Panuccio, A.; Termine, A.; Petrosini, L. CB1 Activity Drives the Selection of Navigational Strategies: A Behavioral and c-Fos Immunoreactivity Study. Int. J. Mol. Sci. 2020, 21, 1072. https://doi.org/10.3390/ijms21031072
Laricchiuta D, Balsamo F, Fabrizio C, Panuccio A, Termine A, Petrosini L. CB1 Activity Drives the Selection of Navigational Strategies: A Behavioral and c-Fos Immunoreactivity Study. International Journal of Molecular Sciences. 2020; 21(3):1072. https://doi.org/10.3390/ijms21031072
Chicago/Turabian StyleLaricchiuta, Daniela, Francesca Balsamo, Carlo Fabrizio, Anna Panuccio, Andrea Termine, and Laura Petrosini. 2020. "CB1 Activity Drives the Selection of Navigational Strategies: A Behavioral and c-Fos Immunoreactivity Study" International Journal of Molecular Sciences 21, no. 3: 1072. https://doi.org/10.3390/ijms21031072