Oxymatrine Inhibits Influenza A Virus Replication and Inflammation via TLR4, p38 MAPK and NF-κB Pathways

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
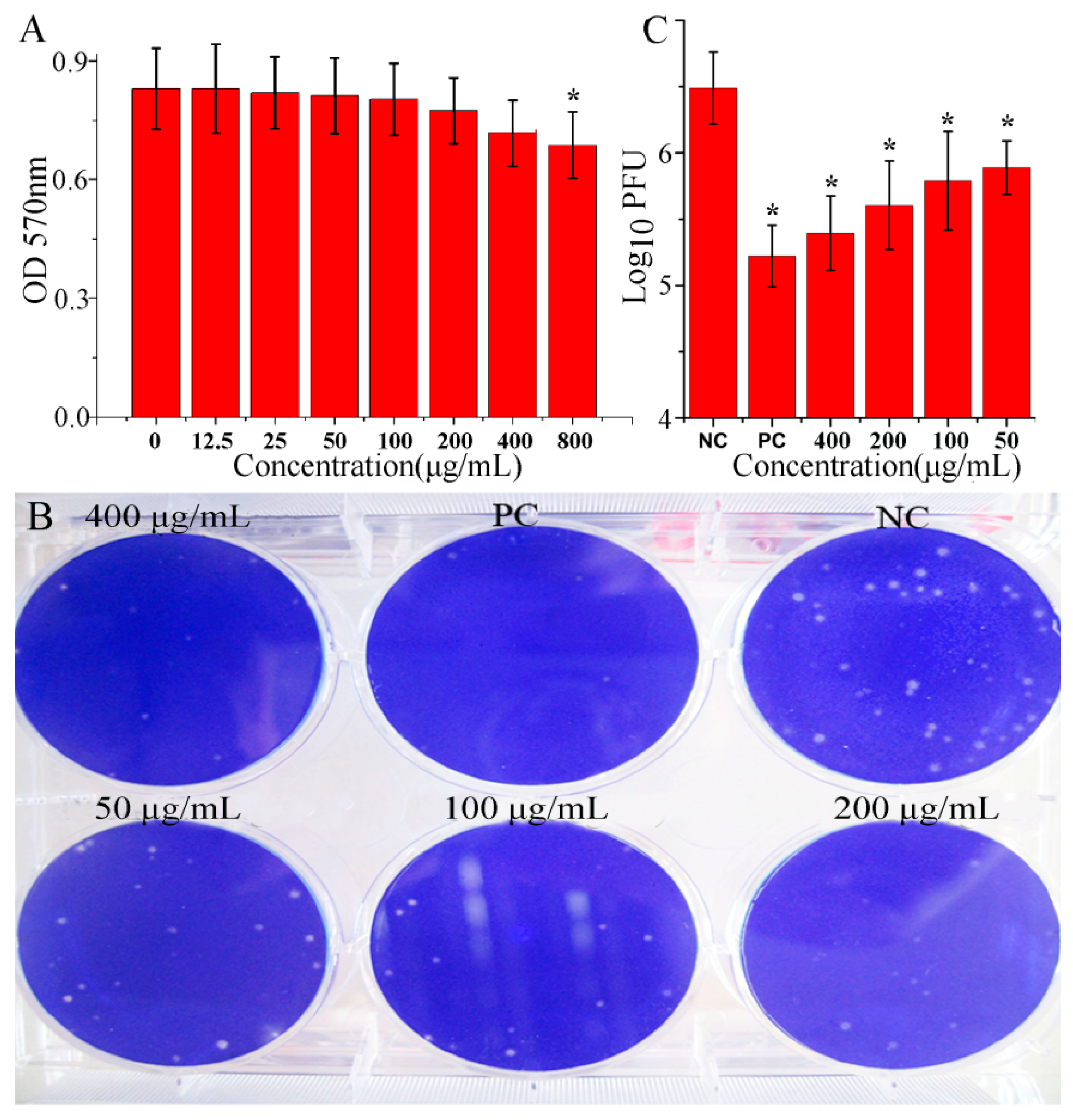
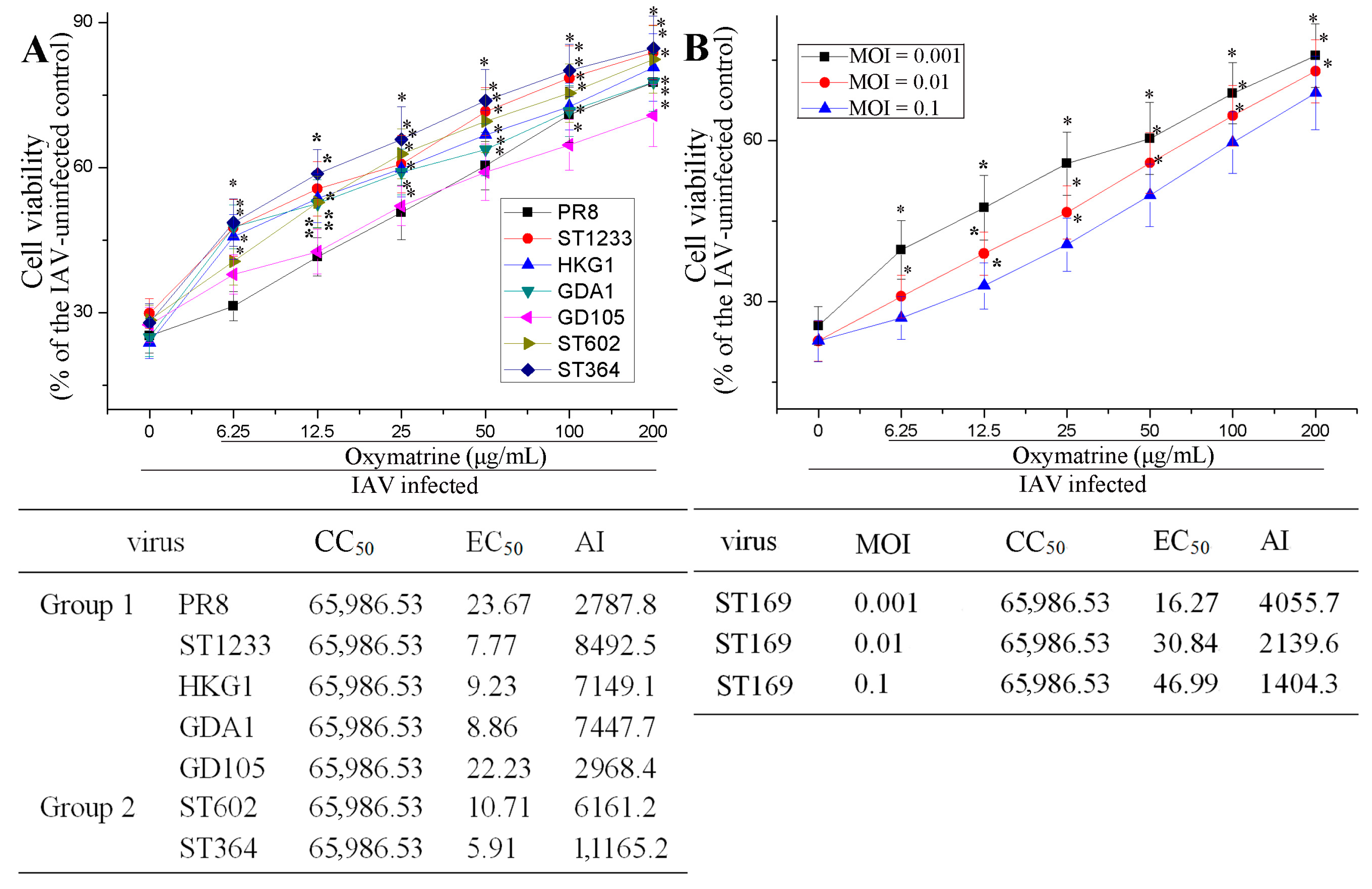
2.1. OMT Could Inhibit IAV Replication In Vitro
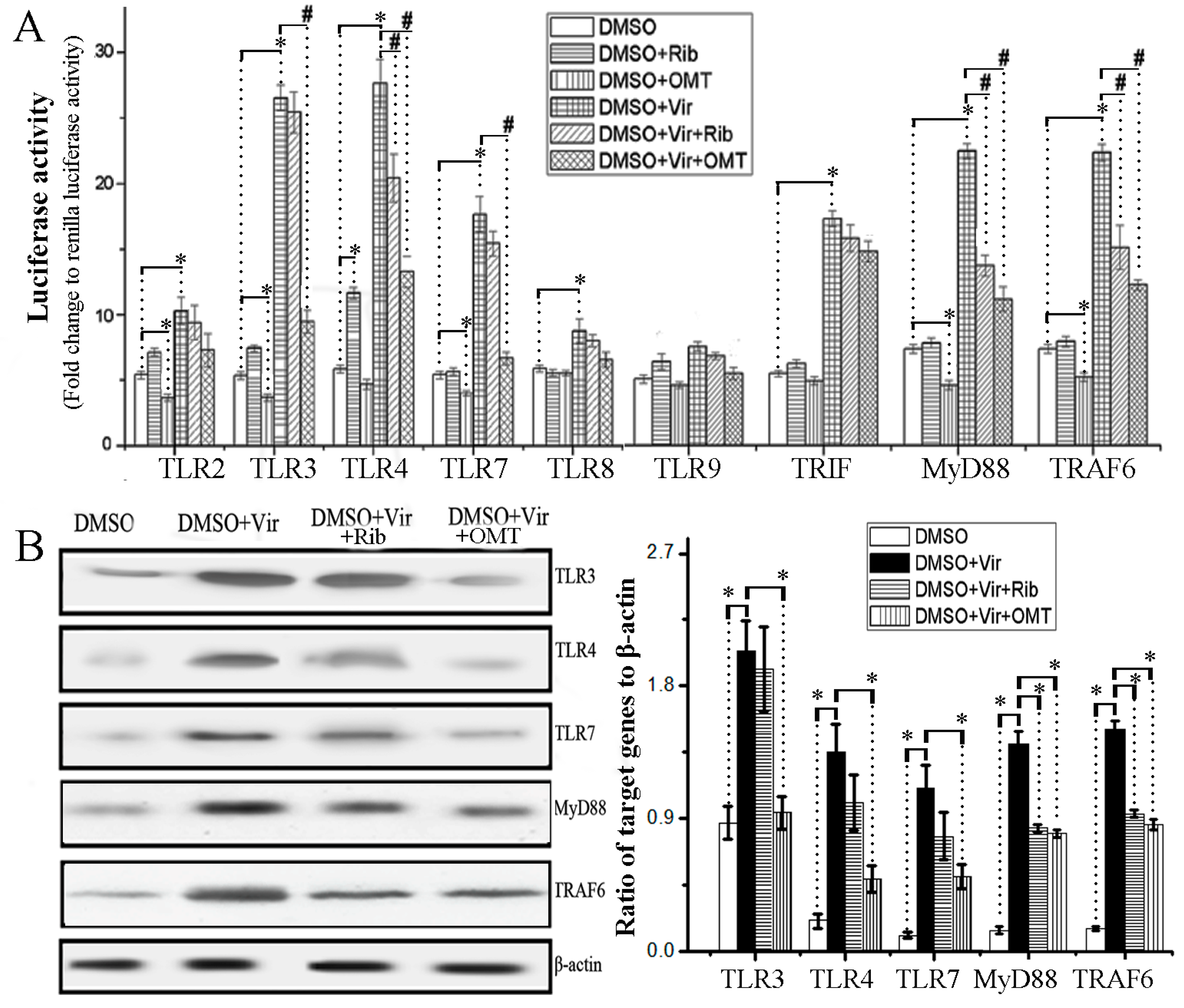
2.2. OMT Could Inhibit IAV-Induced Activation of TLR3/4/7-Myd88-TRAF6 Pathway
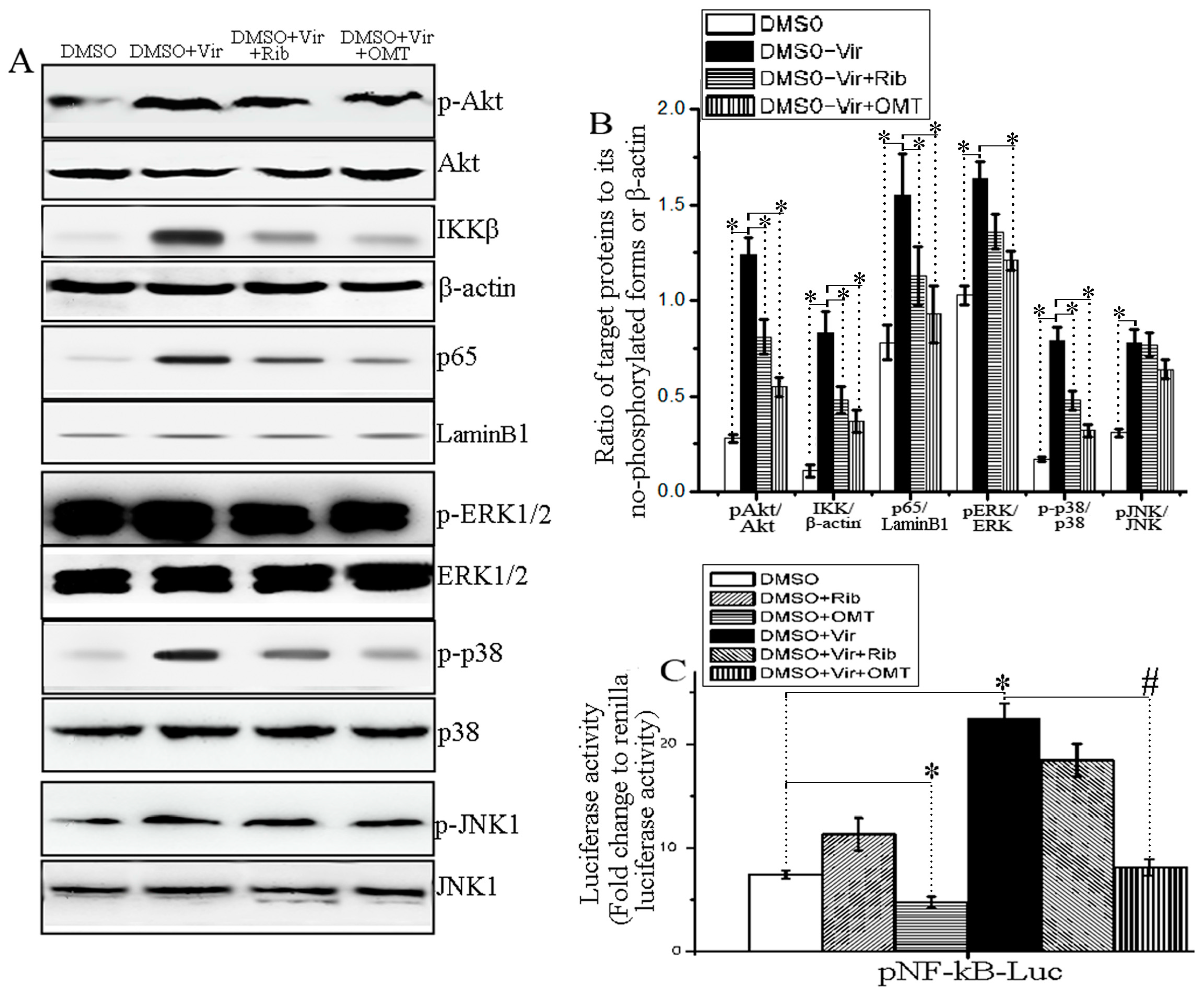
2.3. OMT Could Inhibit IAV-Induced Activation of Akt, ERK1/2, p38 MAPK and NF-κB Pathways
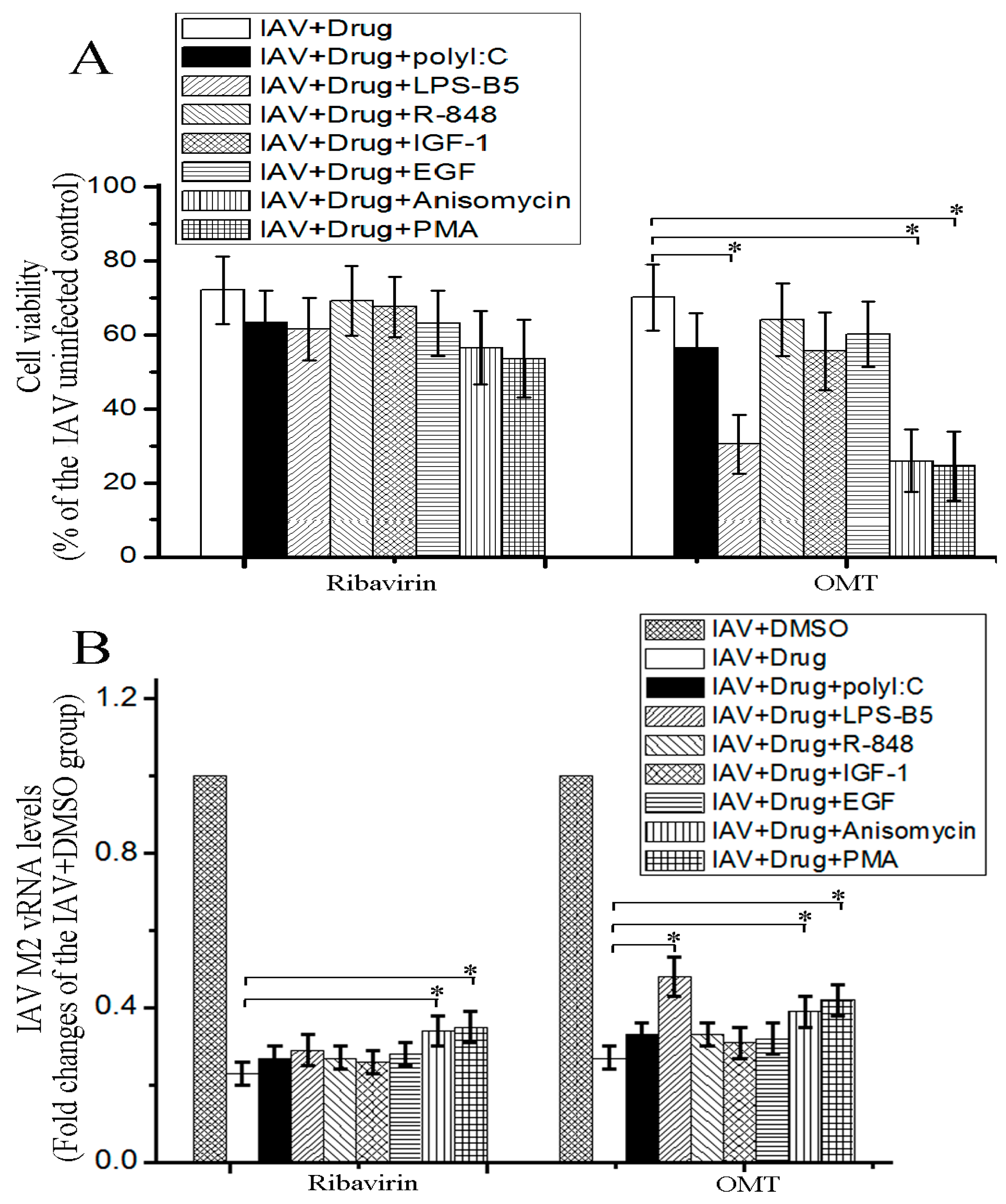
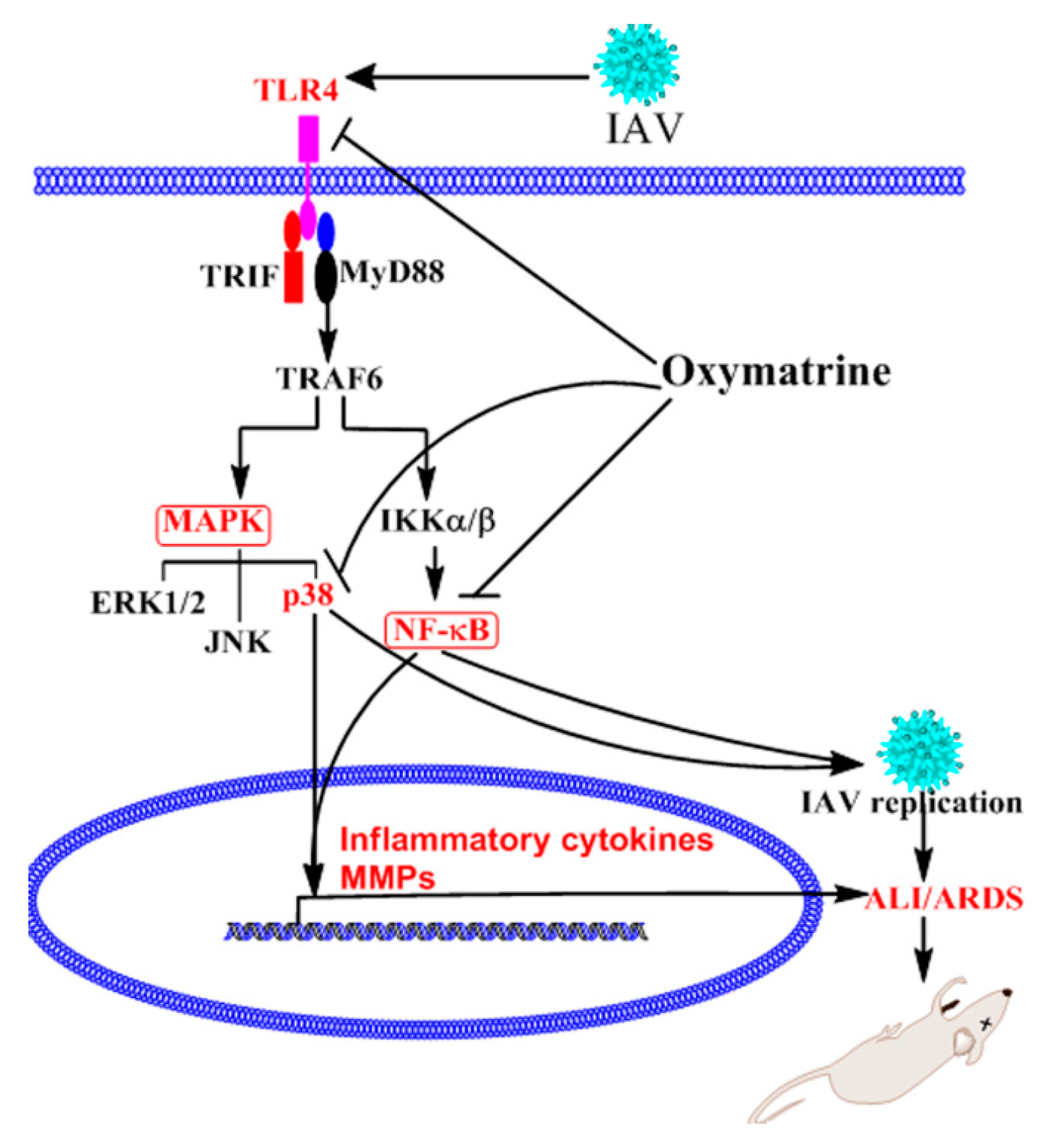
2.4. OMT Might Exert Its Anti-IAV Activity via TLR4, p38 MAPK and NF-κB Signal Pathways
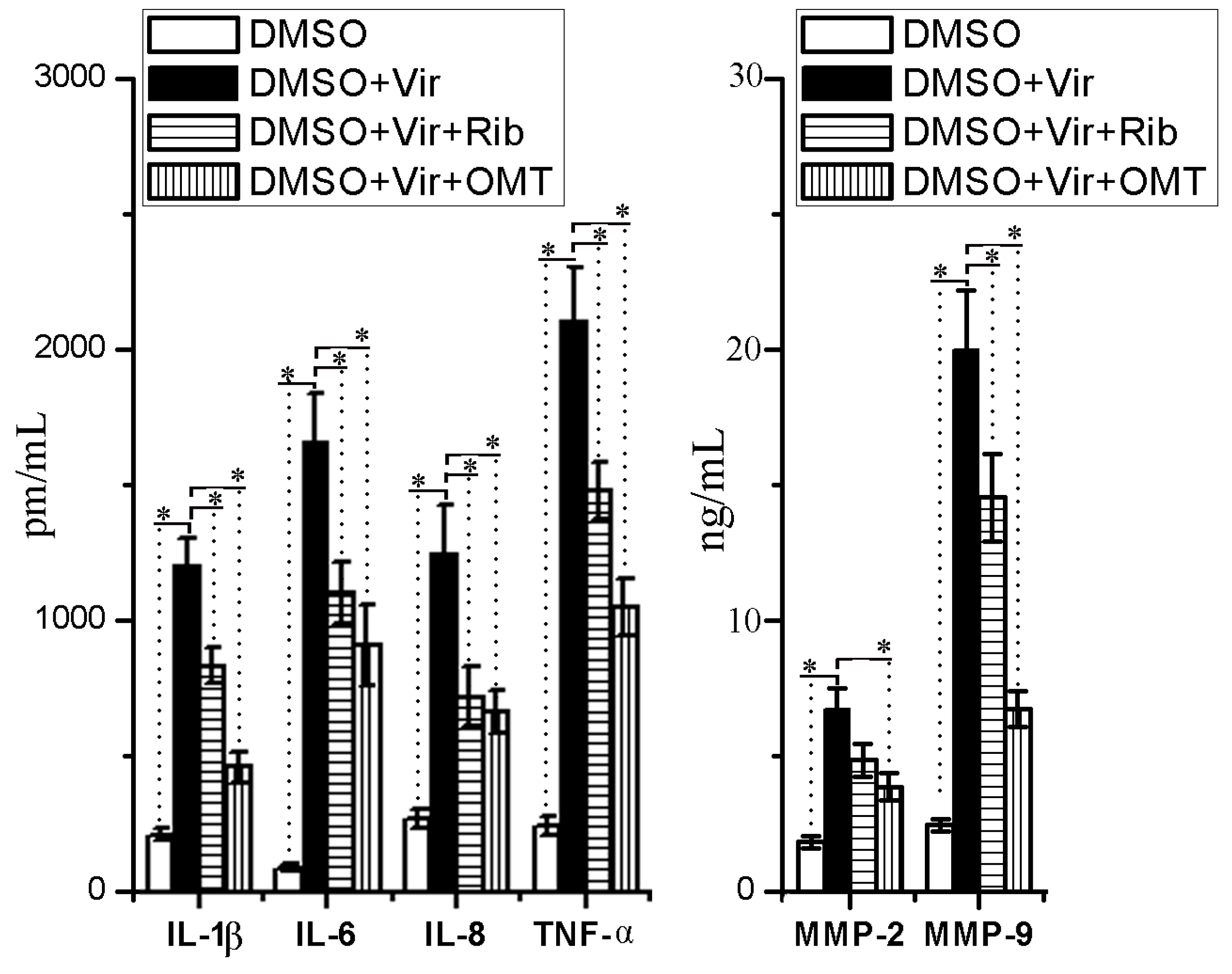
2.5. OMT Could Inhibit IAV-Induced Expressions of Proinflammatory Cytokines and MMP-2/-9
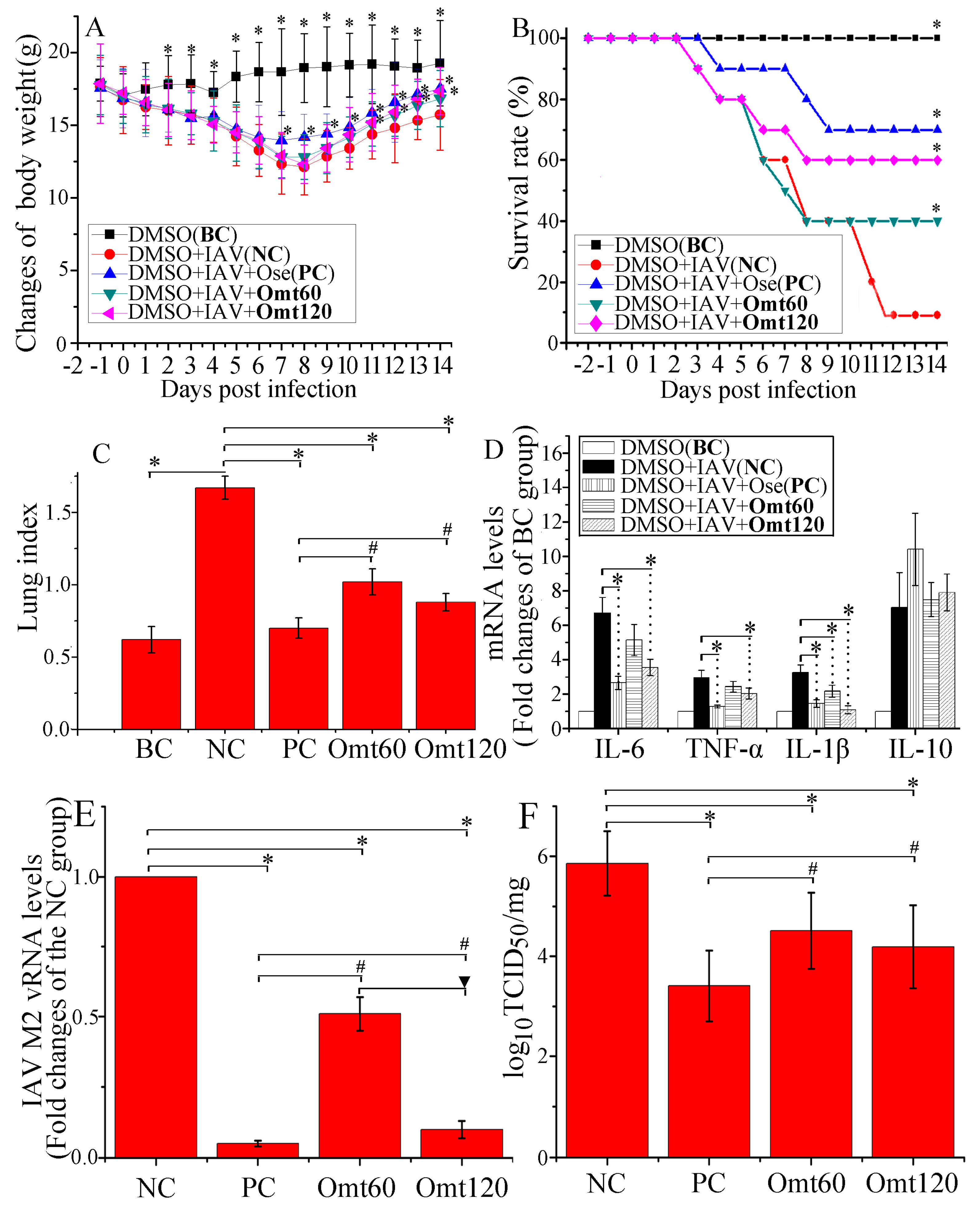
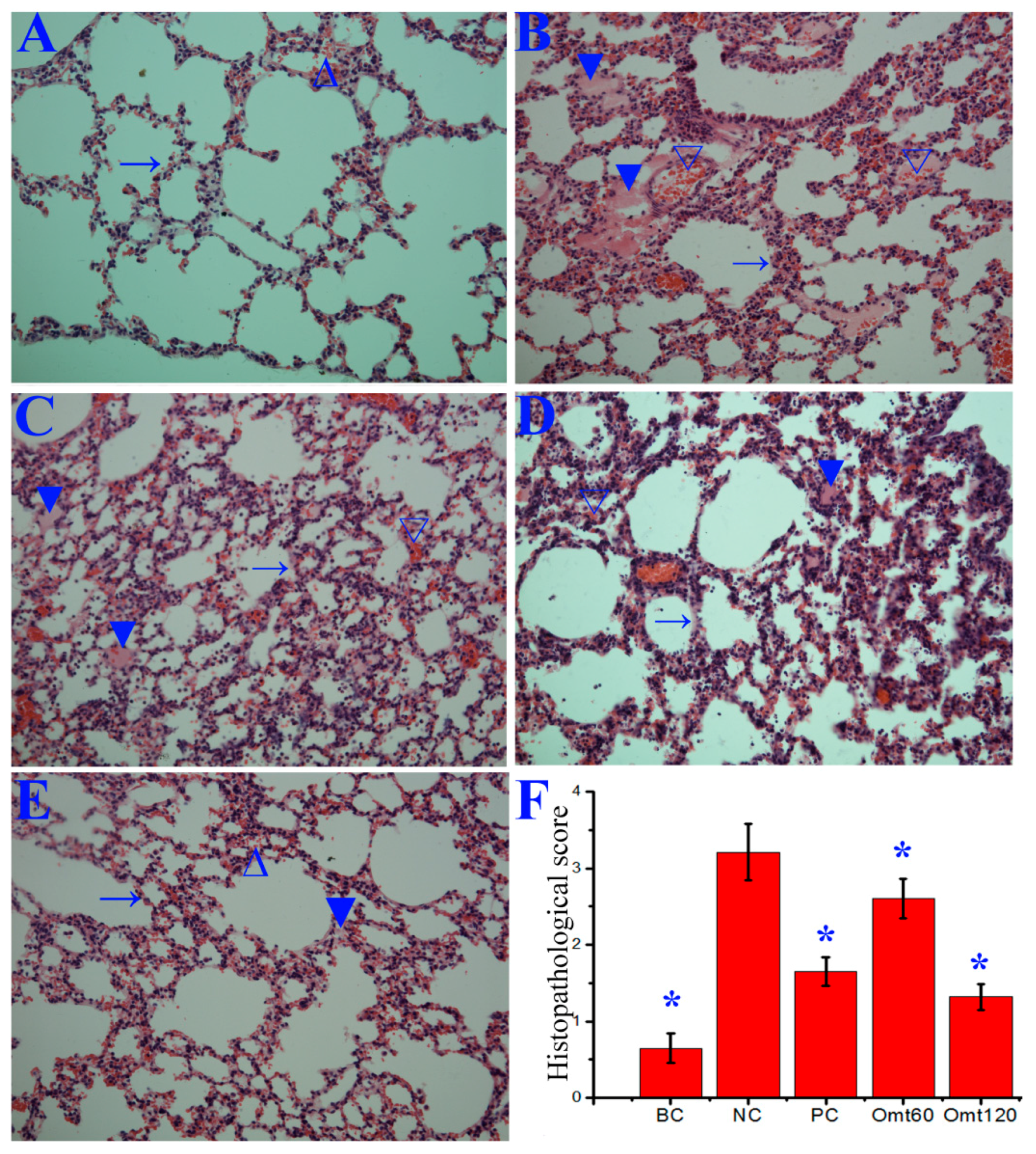
2.6. OMT Could Ameliorate Lung Inflammation and Improved Pulmonary Histopathological Changes after IAV Infection in Mice
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Viruses and Cells
4.3. Plasmid Construction
4.4. Transfection and Luciferase Assay
4.5. Plaque Formation, Plaque Inhibition and Time-of-Addition Assays
4.6. TCID50 Assay and Antiviral Assay by the Sulforhodamine B (SRB) Method
4.7. Quantitative Real-Time RT-PCR
4.8. Western Blotting Assay
4.9. ELISA Assay
4.10. In Vivo Test
- Blank control (BC, n = 16): Mice were not infected with IAV (PR8) but shammed intranasally with VGM medium in a 50 μL volumes, and treated with DMSO (0.5% (v/v)) by oral gavage.
- Negative control (NC, n = 16): Mice were infected intranasally with 10× MLD50 of IAV (PR8) in a 50 μL volumes, and treated with DMSO (0.5% (v/v)) by oral gavage.
- Positive control (PC, n = 16): Mice were infected intranasally with 10× MLD50 of IAV (PR8) viruses in a 50 μL volumes, and treated with oseltamivir (10 mg/kg/day) by oral gavage.
- OMT-treated groups (OMT60 and OMT120, each group n = 16): Mice were infected intranasally with 10× MLD50 of IAV (PR8) viruses in a 50 μL volumes, and treated with OMT (60 mg/kg/day, 120 mg/kg/day) by oral gavage, respectively.
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Sheu, C.C.; Gong, M.N.; Zhai, R.; Chen, F.; Bajwa, E.K.; Clardy, P.F.; Gallagher, D.C.; Thompson, B.T.; Christiani, D.C. Clinical characteristics and outcomes of sepsis-related vs non-sepsis-related ARDS. Chest 2010, 138, 559–567. [Google Scholar] [CrossRef] [PubMed]
- Kuszewski, K.; Brydak, L. The epidemiology and history of influenza. Biomed. Pharmacother. 2000, 54, 188–195. [Google Scholar] [CrossRef]
- Gaur, P.; Munjhal, A.; Lal, S.K. Influenza virus and cell signaling pathways. Med. Sci. Monit. 2011, 17, 148–154. [Google Scholar] [CrossRef]
- Lee, N.; Wong, C.K.; Hui, D.S.; Lee, S.K.; Wong, R.Y.; Ngai, K.L.; Chan, M.C.; Chu, Y.J.; Ho, A.W.; Lui, G.C.; et al. Role of human Toll-like receptors in naturally occurring influenza A infections. Influenza Other Respir. Viruses 2013, 7, 666–675. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Chen, H.; Sun, Y.; Chen, F. Antiviral role of toll-like receptors and cytokines against the new 2009 H1N1 virus infection. Mol. Biol. Rep. 2011, 39, 1163–1172. [Google Scholar] [CrossRef] [PubMed]
- Song, B.M.; Kang, Y.M.; Kim, H.S.; Seo, S.H. Induction of inflammatory cytokines and toll-like receptors in human normal respiratory epithelial cells infected with seasonal H1N1, 2009 pandemic H1N1, seasonal H3N2, and highly pathogenic H5N1 influenza virus. Viral Immunol. 2011, 24, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.Y.; Yano, M.; Kido, H. Effects of inhibitors of Toll-like receptors, protease-activated receptor-2 signalings and trypsin on influenza A virus replication and upregulation of cellular factors in cardiomyocytes. J. Med. Investig. 2011, 58, 19–28. [Google Scholar] [CrossRef]
- Zhang, R.; Ai, X.; Duan, Y.; Xue, M.; He, W.; Wang, C.; Xu, T.; Xu, M.; Liu, B.; Li, C.; et al. Kaempferol ameliorates H9N2 swine influenza virus-induced acute lung injury by inactivation of TLR4/MyD88-mediated NF-kappaB and MAPK signaling pathways. Biomed. Pharmacother. 2017, 89, 660–672. [Google Scholar] [CrossRef] [PubMed]
- Mauad, T.; Hajjar, L.A.; Callegari, G.D.; da Silva, L.F.; Schout, D.; Galas, F.R.; Alves, V.A.; Malheiros, D.M.; Auler, J.O., Jr.; Ferreira, A.F.; et al. Lung pathology in fatal novel human influenza A (H1N1) infection. Am. J. Respir. Crit. Care Med. 2010, 181, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Li, C.R.; Yang, J.; Jia, S.L.; Wang, G.B.; He, Y.X.; Zu, Y.; Yang, W.G.; Fu, D. Changes of immune function in patients with 2009 influenza A (H1N1). Zhonghua Er Ke Za Zhi 2010, 48, 947–953. [Google Scholar] [PubMed]
- Guillot, L.; le Goffic, R.; Bloch, S.; Escriou, N.; Akira, S.; Chignard, M.; Si-Tahar, M. Involvement of toll-like receptor 3 in the immune response of lung epithelial cells to double-stranded RNA and influenza A virus. J. Biol. Chem. 2005, 280, 5571–5580. [Google Scholar] [CrossRef] [PubMed]
- Le Goffic, R.; Balloy, V.; Lagranderie, M.; Alexopoulou, L.; Escriou, N.; Flavell, R.; Chignard, M.; Si-Tahar, M. Detrimental contribution of the Toll-like receptor (TLR)3 to influenza A virus-induced acute pneumonia. PLoS Pathog. 2006, 2, e53. [Google Scholar] [CrossRef] [PubMed]
- Marchant, D.; Singhera, G.K.; Utokaparch, S.; Hackett, T.L.; Boyd, J.H.; Luo, Z.; Si, X.; Dorscheid, D.R.; McManus, B.M.; Hegele, R.G. Toll Like Receptor 4 mediated p38 Mitogen Activated Protein Kinase Activation is a Determinant of Respiratory Virus Entry and Tropism. J. Virol. 2010, 84, 11359–11373. [Google Scholar] [CrossRef] [PubMed]
- Imai, Y.; Kuba, K.; Neely, G.G.; Yaghubian-Malhami, R.; Perkmann, T.; van Loo, G.; Ermolaeva, M.; Veldhuizen, R.; Leung, Y.H.; Wang, H.; et al. Identification of oxidative stress and Toll-like receptor 4 signaling as a key pathway of acute lung injury. Cell 2008, 133, 235–249. [Google Scholar] [CrossRef] [PubMed]
- Shirey, K.A.; Lai, W.; Scott, A.J.; Lipsky, M.; Mistry, P.; Pletneva, L.M.; Karp, C.L.; McAlees, J.; Gioannini, T.L.; Weiss, J.; et al. The TLR4 antagonist Eritoran protects mice from lethal influenza infection. Nature 2013, 497, 498–502. [Google Scholar] [CrossRef] [PubMed]
- Pang, I.K.; Pillai, P.S.; Iwasaki, A. Efficient influenza A virus replication in the respiratory tract requires signals from TLR7 and RIG-I. Proc. Natl. Acad. Sci. USA 2013, 110, 13910–13915. [Google Scholar] [CrossRef] [PubMed]
- Bradley, L.M.; Douglass, M.F.; Chatterjee, D.; Akira, S.; Baaten, B.J. Matrix metalloprotease 9 mediates neutrophil migration into the airways in response to influenza virus-induced toll-like receptor signaling. PLoS Pathog. 2012, 8, e1002641. [Google Scholar] [CrossRef] [PubMed]
- Hirata, N.; Suizu, F.; Matsuda-Lennikov, M.; Edamura, T.; Bala, J.; Noguchi, M. Inhibition of Akt kinase activity suppresses entry and replication of influenza virus. Biochem. Biophys. Res. Commun. 2014, 450, 891–898. [Google Scholar] [CrossRef] [PubMed]
- Pleschka, S.; Wolff, T.; Ehrhardt, C.; Hobom, G.; Planz, O.; Rapp, U.R.; Ludwig, S. Influenza virus propagation is impaired by inhibition of the Raf/MEK/ERK signalling cascade. Nat. Cell Biol. 2001, 3, 301–305. [Google Scholar] [CrossRef] [PubMed]
- Nacken, W.; Ehrhardt, C.; Ludwig, S. Small molecule inhibitors of the c-Jun N-terminal kinase (JNK) possess antiviral activity against highly pathogenic avian and human pandemic influenza A viruses. Biol. Chem. 2012, 393, 525–534. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Xin, Z.T.; Liang, Y.; Ly, H.; Liang, Y. NF-κB signaling differentially regulates influenza virus RNA synthesis. J. Virol. 2008, 82, 9880–9889. [Google Scholar] [CrossRef] [PubMed]
- Ling, J.Y.; Zhang, G.Y.; Cui, Z.J.; Zhang, C.K. Supercritical fluid extraction of quinolizidine alkaloids from Sophora flavescens Ait and purification by high-speed counter-current chromatography. J. Chromatogr. A 2007, 1145, 123–127. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.L.; Yao, L.; Rao, S.Y.; Gong, Z.N.; Zhang, S.Q.; Yu, S.Q. Attenuation of acute lung injury in mice by oxymatrine is associated with inhibition of phosphorylated p38 mitogen-activated protein kinase. J. Eethnopharmacol. 2005, 98, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Wang, X.; Wang, X.; Hou, X.; Teng, P.; Jiang, Y.; Zhang, L.; Yang, X.; Tian, J.; Li, G.; et al. Oxymatrine protects against myocardial injury via inhibition of JAK2/STAT3 signaling in rat septic shock. Mol. Med. Rep. 2013, 7, 1293–1299. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.Q.; Yu, W.H.; Hu, Y.Y.; Zhang, Z.Y.; Huang, M. Oxymatrine reduces neuronal cell apoptosis by inhibiting Toll-like receptor 4/nuclear factor κB-dependent inflammatory responses in traumatic rat brain injury. Inflamm. Res. 2011, 60, 533–539. [Google Scholar] [CrossRef] [PubMed]
- Murray, J.L.; McDonald, N.J.; Sheng, J.; Shaw, M.W.; Hodge, T.W.; Rubin, D.H.; O’Brien, W.A.; Smee, D.F. Inhibition of influenza A virus replication by antagonism of a PI3K-AKT-mTOR pathway member identified by gene-trap insertional mutagenesis. Antivir. Chem. Chemother. 2012, 22, 205–215. [Google Scholar] [CrossRef] [PubMed]
- Marjuki, H.; Alam, M.I.; Ehrhardt, C.; Wagner, R.; Planz, O.; Klenk, H.D.; Ludwig, S.; Pleschka, S. Membrane accumulation of influenza A virus hemagglutinin triggers nuclear export of the viral genome via protein kinase Calpha-mediated activation of ERK signaling. J. Biol. Chem. 2006, 281, 16707–16715. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.; Li, L.; Zhang, X.; Liu, Y.; Yang, C.; Yang, Y.; Yin, J. Oxymatrine downregulates TLR4, TLR2, MyD88, and NF-κB and protects rat brains against focal ischemia. Mediat. Inflamm. 2009, 2009, 704706. [Google Scholar] [CrossRef] [PubMed]
- Wen, J.B.; Zhu, F.Q.; Chen, W.G.; Jiang, L.P.; Chen, J.; Hu, Z.P.; Huang, Y.J.; Zhou, Z.W.; Wang, G.L.; Lin, H.; et al. Oxymatrine improves intestinal epithelial barrier function involving NF-κB-mediated signaling pathway in CCl4-induced cirrhotic rats. PLoS ONE 2014, 9, e106082. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.Q.; Du, Q.; Yu, W.H.; Zhang, Z.Y.; Zhu, Q.; Che, Z.H.; Chen, F.; Wang, H.; Chen, J. Anti-inflammatory Effects of Oxymatrine Through Inhibition of Nuclear Factor-κB and Mitogen-activated Protein Kinase Activation in Lipopolysaccharide-induced BV2 Microglia Cells. Iran. J. Pharm. Res. 2013, 12, 165–174. [Google Scholar] [PubMed]
- Ge, H.; Wang, Y.F.; Xu, J.; Gu, Q.; Liu, H.B.; Xiao, P.G.; Zhou, J.; Liu, Y.; Yang, Z.; Su, H. Anti-influenza agents from Traditional Chinese Medicine. Nat. Prod. Rep. 2010, 27, 1758–1780. [Google Scholar] [CrossRef] [PubMed]
- National Health and Family Planning Commission of the People’s Republic of China. Guidelines for Diagnosis and Treatment of Influenza (2011 Edition). Available online: http://www.nhfpc.gov.cn/zwgk/wtwj/201304/7fd812ef8dc54291af5060957eea304c.shtml (accessed on 17 February 2011).
- National Health and Family Planning Commission of the People’s Republic of China. Diagnosis and Treatment of Influenza (2018 Edition). Available online: http://www.nhfpc.gov.cn/yzygj/s3594q/201801/5737c258bb0c4a0493cb4f65fcf11be1.shtml (accessed on 8 January 2018).
- Lu, L.G.; Zeng, M.D.; Mao, Y.M.; Li, J.Q.; Wan, M.B.; Li, C.Z.; Chen, C.W.; Fu, Q.C.; Wang, J.Y.; She, W.M.; et al. Oxymatrine therapy for chronic hepatitis B: A randomized double-blind and placebo-controlled multi-center trial. World J. Gastroenterol. 2003, 9, 2480–2483. [Google Scholar] [CrossRef] [PubMed]
- Marjuki, H.; Gornitzky, A.; Marathe, B.M.; Ilyushina, N.A.; Aldridge, J.R.; Desai, G.; Webby, R.J.; Webster, R.G. Influenza A virus-induced early activation of ERK and PI3K mediates V-ATPase-dependent intracellular pH change required for fusion. Cell. Microbiol. 2011, 13, 587–601. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Le, T.Q.; Chida, J.; Cisse, Y.; Yano, M.; Kido, H. Mechanisms of matrix metalloproteinase-9 upregulation and tissue destruction in various organs in influenza A virus infection. J. Med. Investig. 2010, 57, 26–34. [Google Scholar] [CrossRef]
- Lee, Y.H.; Lai, C.L.; Hsieh, S.H.; Shieh, C.C.; Huang, L.M.; Wu-Hsieh, B.A. Influenza A virus induction of oxidative stress and MMP-9 is associated with severe lung pathology in a mouse model. Virus Res. 2013, 178, 411–422. [Google Scholar] [CrossRef] [PubMed]
- Guo, B.; Zhang, T.; Su, J.; Wang, K.; Li, X. Oxymatrine targets EGFR(p-Tyr845) and inhibits EGFR-related signaling pathways to suppress the proliferation and invasion of gastric cancer cells. Cancer Chemother. Pharmacol. 2015, 75, 353–363. [Google Scholar] [CrossRef] [PubMed]
- Chan, M.H.; Chan, P.K.; Griffith, J.F.; Chan, I.H.; Lit, L.C.; Wong, C.K.; Antonio, G.E.; Liu, E.Y.; Hui, D.S.; Suen, M.W.; et al. Steroid-induced osteonecrosis in severe acute respiratory syndrome: A retrospective analysis of biochemical markers of bone metabolism and corticosteroid therapy. Pathology 2006, 38, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.P.; Zhao, X.F.; Zeng, J.; Wan, Q.Y.; Yang, J.C.; Li, W.Z.; Chen, X.X.; Wang, G.F.; Li, K.S. Drug Screening for Autophagy Inhibitors Based on the Dissociation of Beclin1-Bcl2 Complex Using BiFC Technique and Mechanism of Eugenol on Anti-Influenza A Virus Activity. PLoS ONE 2013, 8, e61026. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.P.; Chen, J.; Bei, Y.F.; Han, B.X.; Wang, S. Influence of borneol on primary mice oral fibroblasts: A penetration enhancer may be used in oral submucous fibrosis. J. Oral Pathol. Med. 2009, 38, 276–281. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.; Wang, G.; Li, W.; Zhang, L.; Yang, J.; Zhao, X.; Chen, X.; Xu, Y.; Li, K. High-Throughput Screening for AntiInfluenza A Virus Drugs and Study of the Mechanism of Procyanidin on Influenza A VirusInduced Autophagy. J. Biomol. Screen. 2012, 17, 605–617. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.J.; Lim, C.H.; Song, J.H.; Baek, S.H.; Kwon, D.H. Antiviral activity of raoulic acid from Raoulia australis against Picornaviruses. Phytomedicine 2009, 16, 35–39. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.P.; Wu, L.Q.; Li, R.; Zhao, X.F.; Wan, Q.Y.; Chen, X.X.; Li, W.Z.; Wang, G.F.; Li, K.S. Identification of 23-(s)-2-amino-3-phenylpropanoyl-silybin as an antiviral agent for influenza a virus infection in vitro and in vivo. Antimicrob. Agents Chemother. 2013, 57, 4433–4443. [Google Scholar] [CrossRef] [PubMed]
- McGrath, J.C.; Lilley, E. Implementing guidelines on reporting research using animals (ARRIVE etc.): New requirements for publication in BJP. Br. J. Pharmacol. 2015, 172, 3189–3193. [Google Scholar] [CrossRef] [PubMed]
- Zhou, R.; Xu, Q.; Xu, Y.; Xiong, A.; Wang, Y.; Ma, P. Oxymatrine attenuated isoproterenol-induced heart failure in rats via regulation of COX-2/PGI2 pathway. Biomed. Pharmacother. 2016, 84, 1359–1366. [Google Scholar] [CrossRef] [PubMed]
- Bae, H.B.; Jeong, C.W.; Li, M.; Kim, H.S.; Kwak, S.H. Effects of urinary trypsin inhibitor on lipopolysaccharide-induced acute lung injury in rabbits. Inflammation 2012, 35, 176–182. [Google Scholar] [CrossRef] [PubMed]









© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dai, J.-P.; Wang, Q.-W.; Su, Y.; Gu, L.-M.; Deng, H.-X.; Chen, X.-X.; Li, W.-Z.; Li, K.-S. Oxymatrine Inhibits Influenza A Virus Replication and Inflammation via TLR4, p38 MAPK and NF-κB Pathways. Int. J. Mol. Sci. 2018, 19, 965. https://doi.org/10.3390/ijms19040965
Dai J-P, Wang Q-W, Su Y, Gu L-M, Deng H-X, Chen X-X, Li W-Z, Li K-S. Oxymatrine Inhibits Influenza A Virus Replication and Inflammation via TLR4, p38 MAPK and NF-κB Pathways. International Journal of Molecular Sciences. 2018; 19(4):965. https://doi.org/10.3390/ijms19040965
Chicago/Turabian StyleDai, Jian-Ping, Qian-Wen Wang, Yun Su, Li-Ming Gu, Hui-Xiong Deng, Xiao-Xuan Chen, Wei-Zhong Li, and Kang-Sheng Li. 2018. "Oxymatrine Inhibits Influenza A Virus Replication and Inflammation via TLR4, p38 MAPK and NF-κB Pathways" International Journal of Molecular Sciences 19, no. 4: 965. https://doi.org/10.3390/ijms19040965