
The Small Molecule PPARγ Agonist GL516 Induces Feeding-Stimulatory Effects in Hypothalamus Cells Hypo-E22 and Isolated Hypothalami


 , ,
, ,  and
and 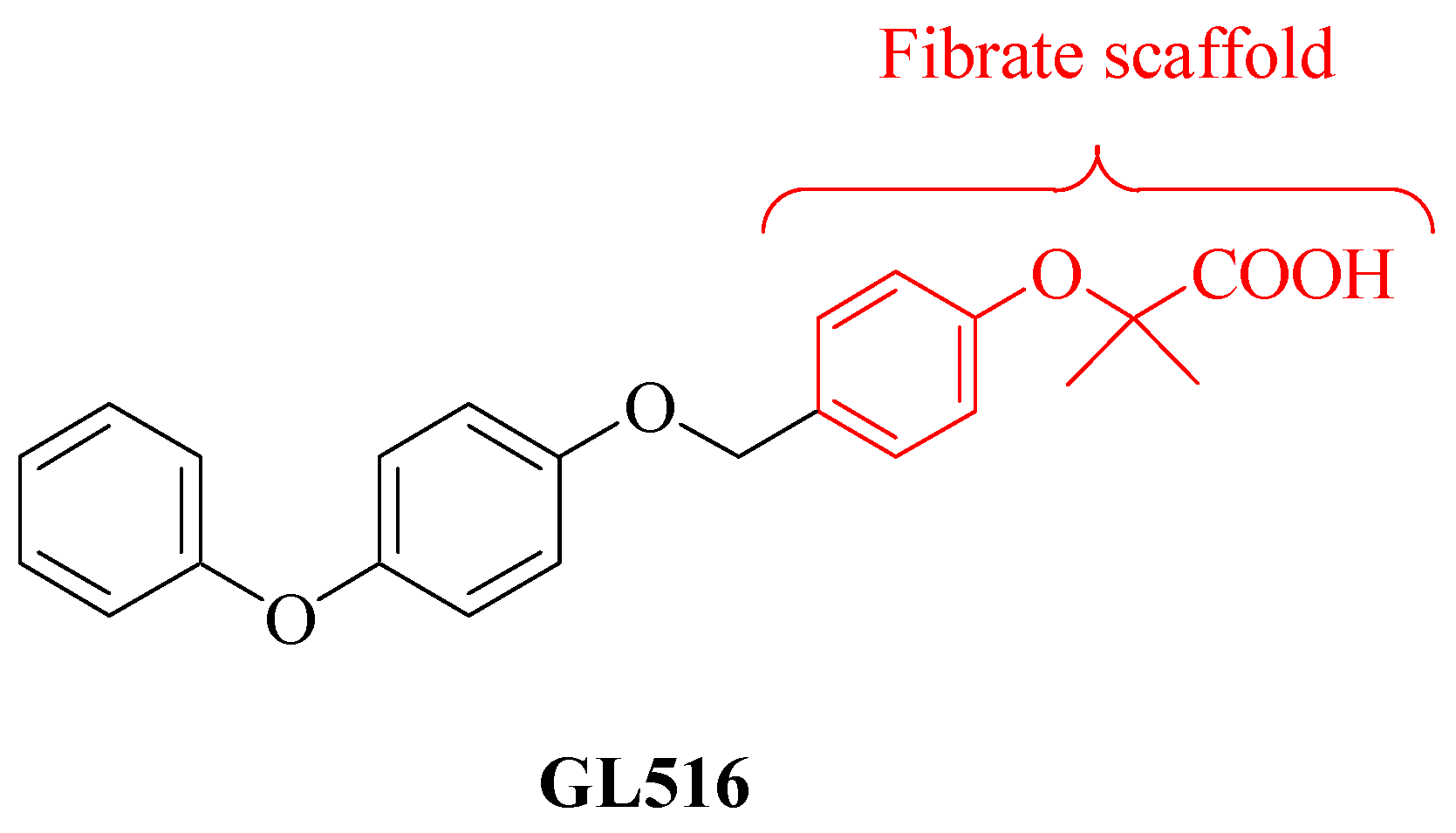{kind=link}
{kind=link}
{kind=link}
{kind=link}
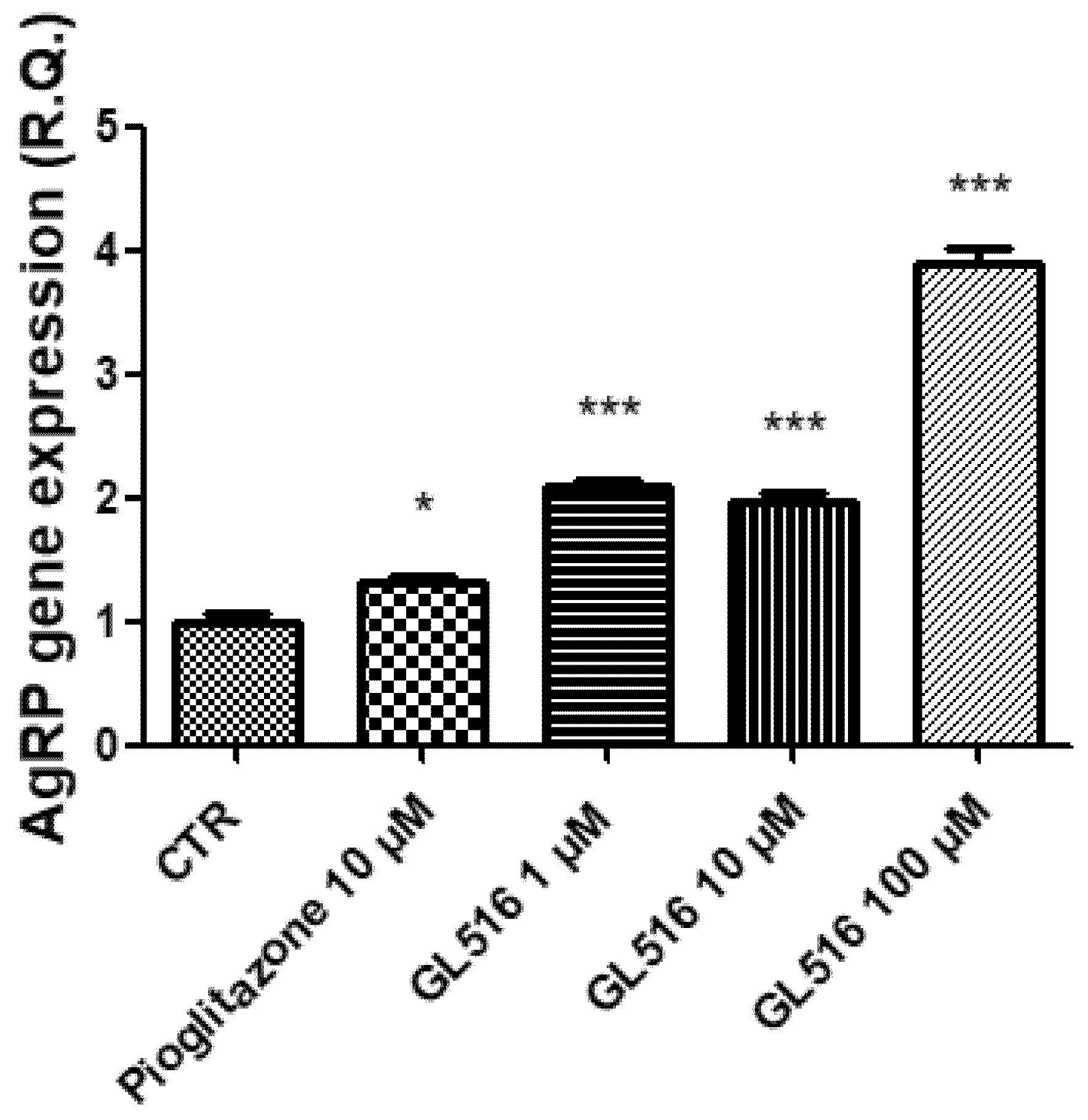{kind=link}
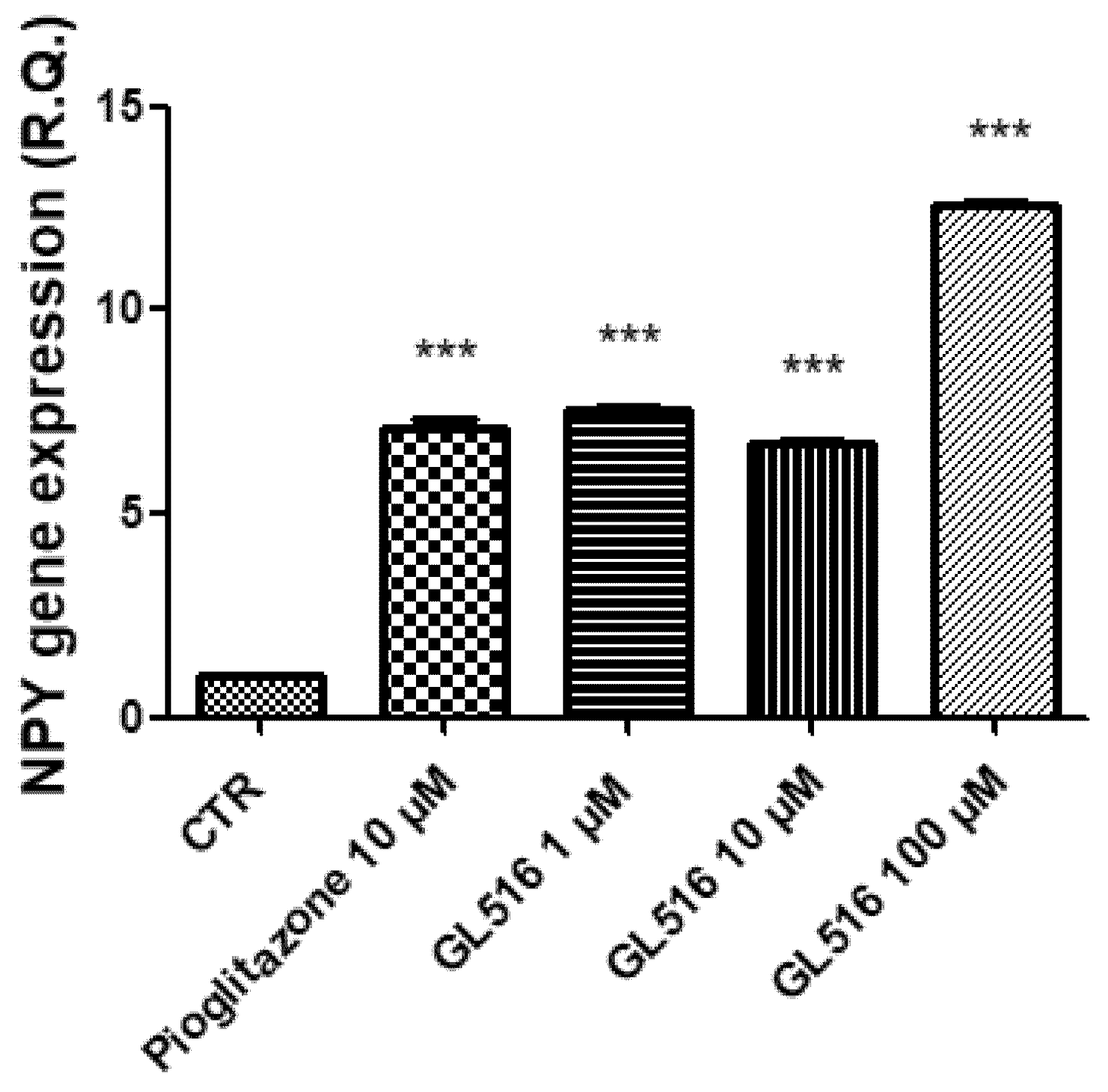{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
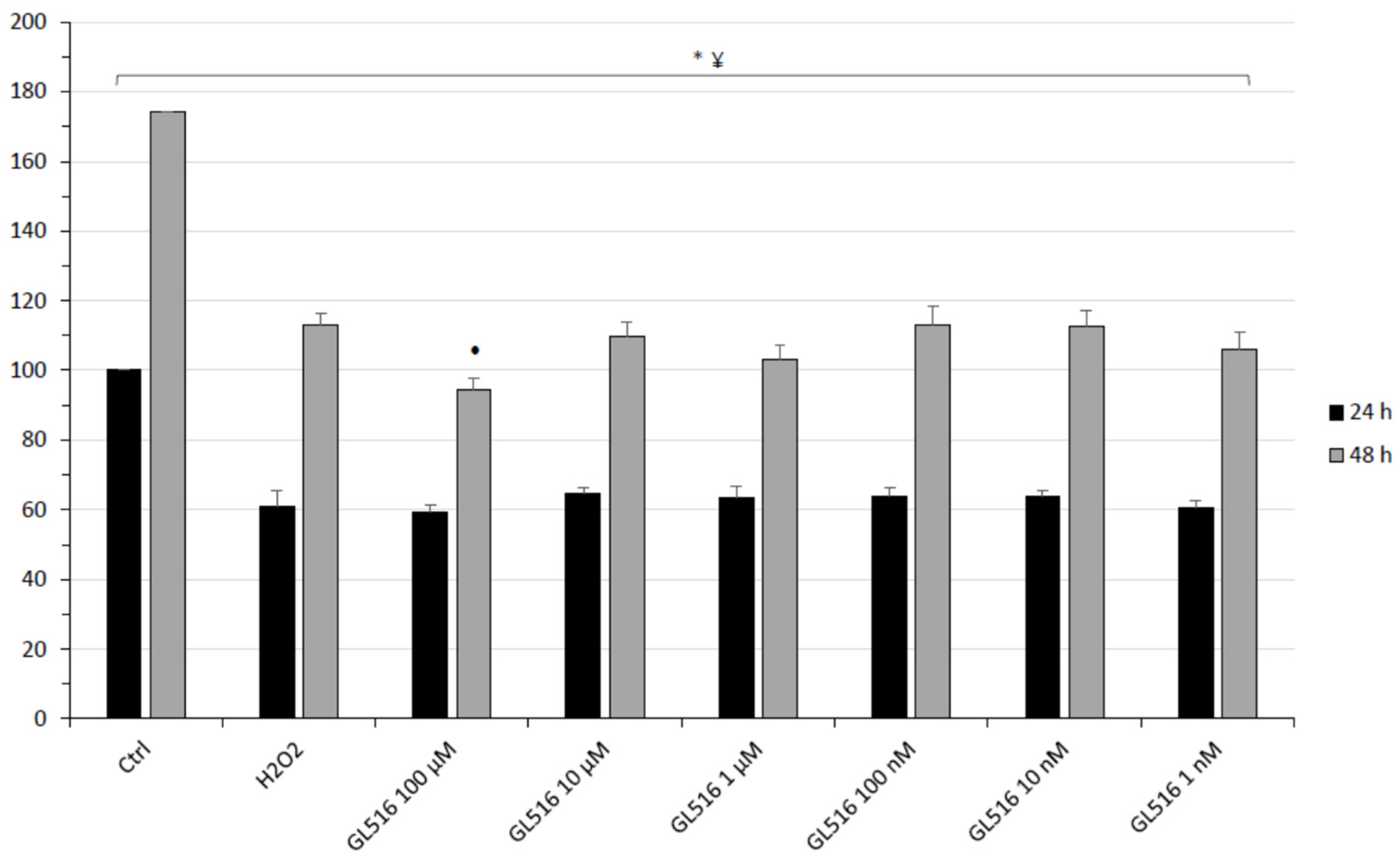
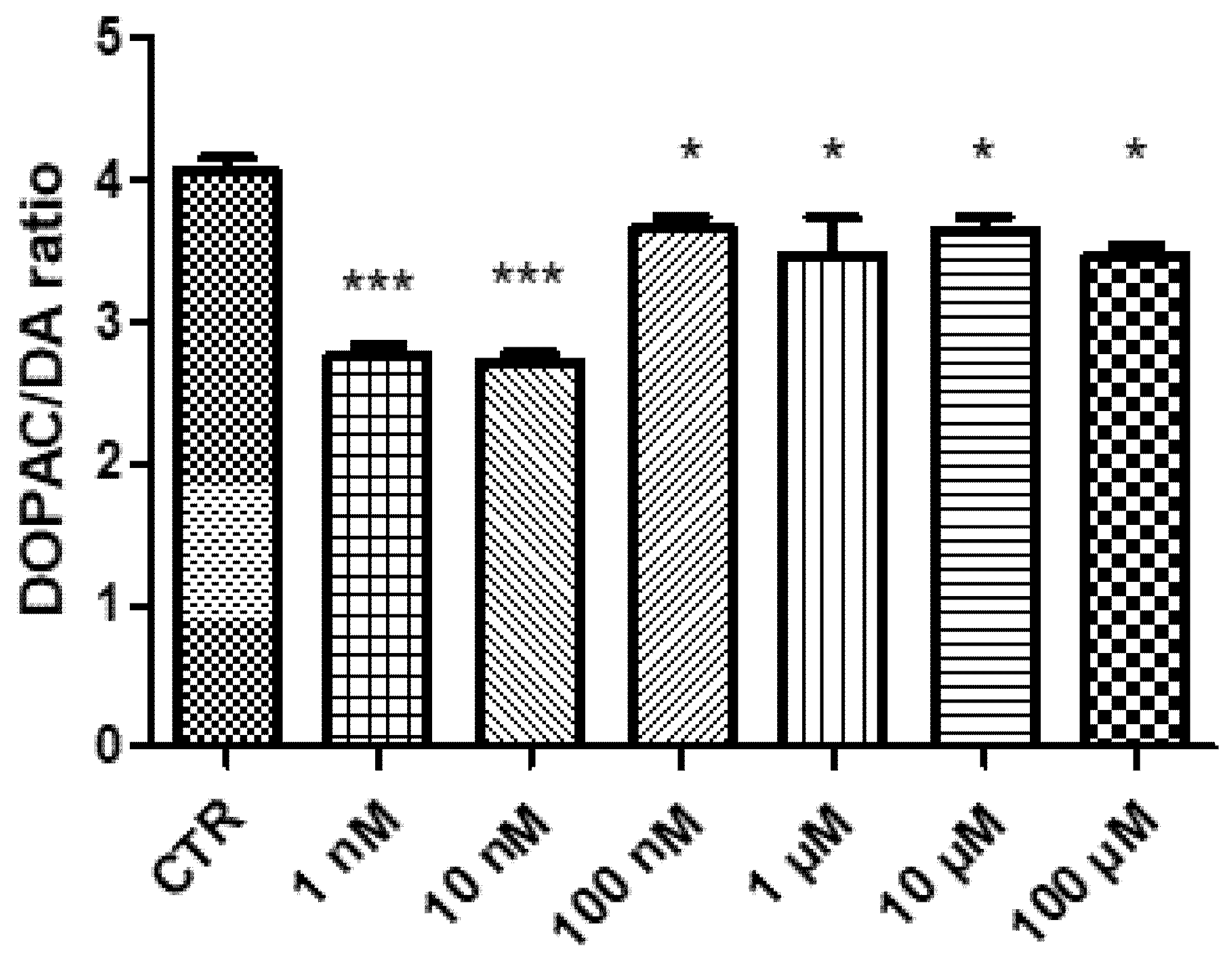
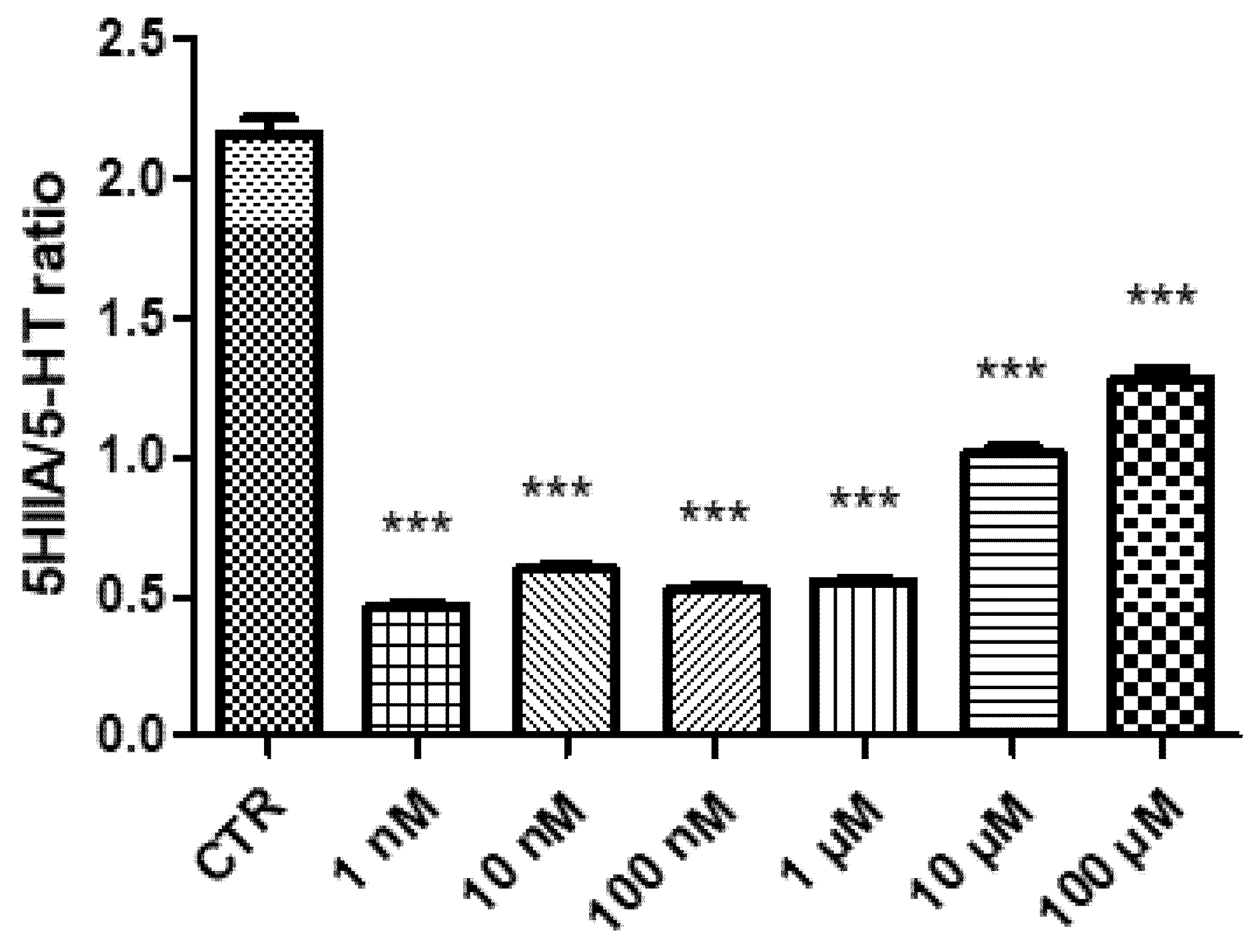
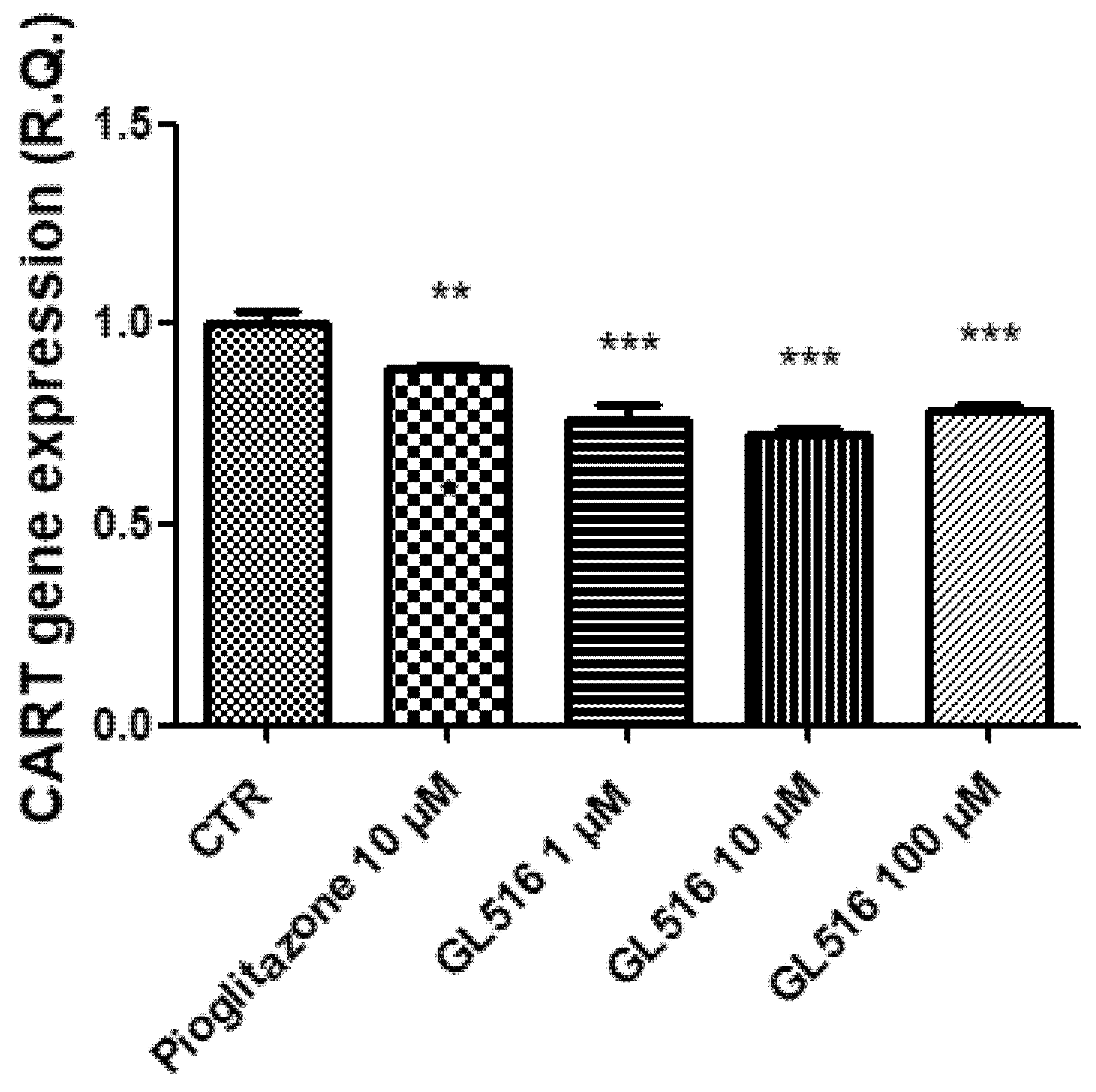
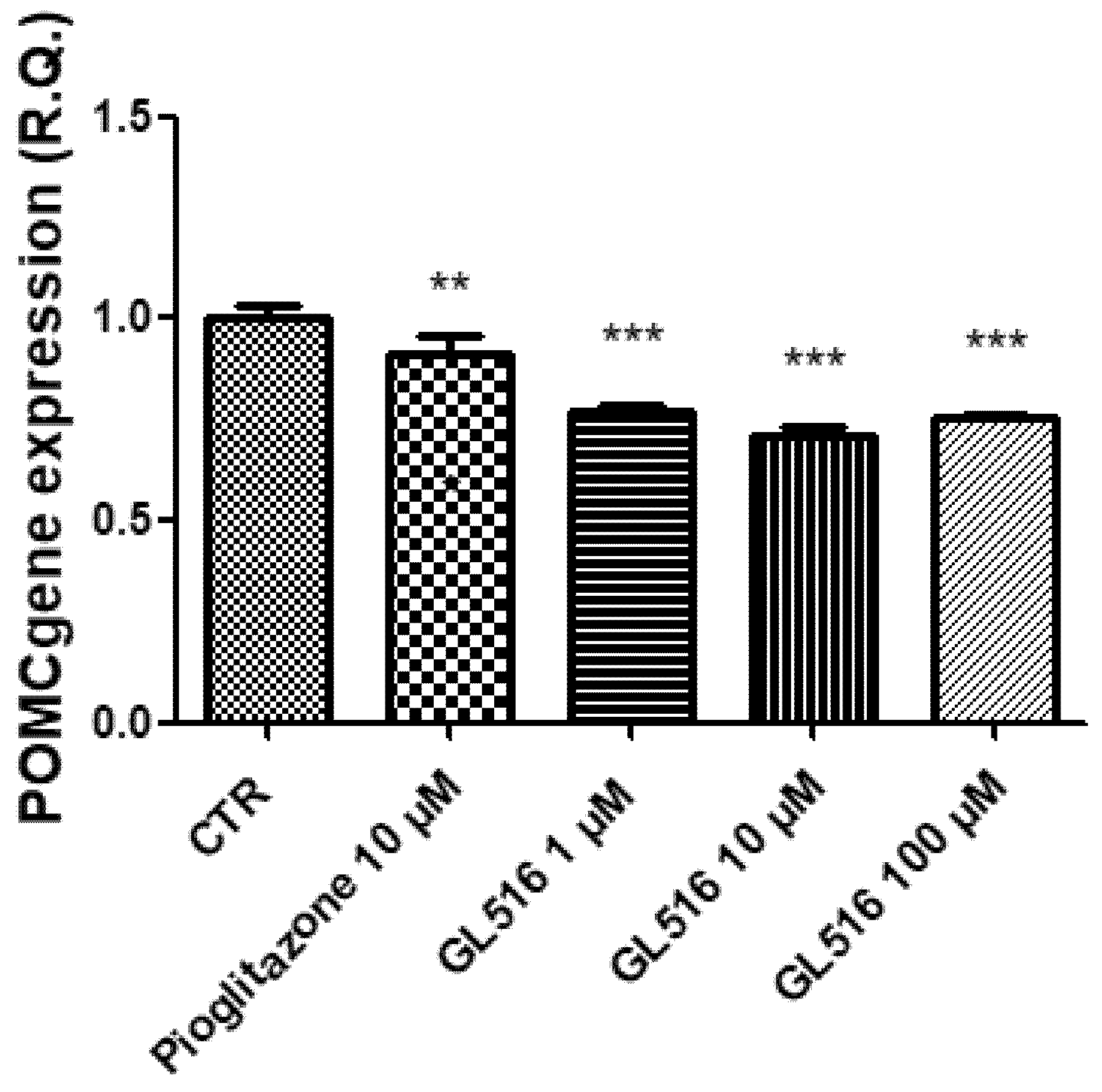
2. Results and Discussion
3. Materials and Methods
3.1. Hypothalamic HypoE22 cells: Evaluation of Neuromodulatory Effects
3.2. Quantitative Determination of Dopamine and Serotonin
3.3. Ex Vivo Studies
3.4. RNA Extraction, Reverse Transcription, and Real-Time Reverse Transcription Polymerase Chain Reaction (RT-PCR)
3.5. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Kota, B.P.; Huang, T.H.-W.; Roufogalis, B.D. An overview on biological mechanisms of PPARs. Pharmacol. Res. 2005, 51, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Wahli, W.; Michalik, L. PPARs at the crossroads of lipid signaling and inflammation. Trends Endocrinol. Metab. 2012, 23, 351–363. [Google Scholar] [CrossRef] [PubMed]
- Moreno, S.; Farioli-Vecchioli, S.; Cerù, M.P. Immunolocalization of peroxisome proliferator-activated receptors and retinoid X receptors in the adult rat CNS. Neuroscience 2004, 123, 131–145. [Google Scholar] [CrossRef] [PubMed]
- Bernardo, A.; Minghetti, L. Regulation of Glial Cell Functions by PPAR-γ Natural and Synthetic Agonists. PPAR Res. 2008, 2008, 864140. [Google Scholar] [CrossRef] [Green Version]
- Chawla, A. Control of macrophage activation and function by PPARs. Circ. Res. 2010, 106, 1559–1569. [Google Scholar] [CrossRef]
- Sanjay, S.A.; Lee, H.-J. Role of Phytoconstituents as PPAR Agonists: Implications for Neurodegenerative Disorders. Biomedicines 2021, 9, 1914. [Google Scholar] [CrossRef]
- Behl, T.; Madaan, P.; Sehgal, A.; Singh, S.; Sharma, N.; Bhatia, S.; Al-Harrasi, A.; Chigurupati, S.; Alrashdi, I.; Bungau, S.G. Elucidating the Neuroprotective Role of PPARs in Parkinson’s Disease: A Neoteric and Prospective Target. Int. J. Mol. Sci. 2021, 22, 10161. [Google Scholar] [CrossRef]
- Lim, J.; Kim, H.I.; Bang, Y.; Choi, H.J. Peroxisome proliferator-activated receptor gamma: A novel therapeutic target for cognitive impairment and mood disorders that functions via the regulation of adult neurogenesis. Arch. Pharmacal Res. 2021, 44, 553–563. [Google Scholar] [CrossRef]
- Villapol, S. Roles of Peroxisome Proliferator-Activated Receptor Gamma on Brain and Peripheral Inflammation. Cell. Mol. Neurobiol. 2018, 38, 121–132. [Google Scholar] [CrossRef]
- Cai, W.; Yang, T.; Liu, H.; Han, L.; Zhang, K.; Hu, X.; Zhang, X.; Yin, K.-J.; Gao, Y.; Bennett, M.V.L.; et al. Peroxisome proliferator-activated receptor γ (PPARγ): A master gatekeeper in CNS injury and repair. Prog. Neurobiol. 2018, 163, 27–58. [Google Scholar] [CrossRef]
- Mannan, A.; Garg, N.; Singh, T.G.; Kang, H.K. Peroxisome Proliferator-Activated Receptor-Gamma (PPAR-ɣ): Molecular Effects and Its Importance as a Novel Therapeutic Target for Cerebral Ischemic Injury. Neurochem. Res. 2021, 46, 2800–2831. [Google Scholar] [CrossRef] [PubMed]
- Ngo, S.T.; Mi, J.D.; Henderson, R.D.; McCombe, P.A.; Steyn, F.J. Exploring targets and therapies for amyotrophic lateral sclerosis: Current insights into dietary interventions. Degener. Neurol. Neuromuscul. Dis. 2017, 7, 95–108. [Google Scholar] [CrossRef] [Green Version]
- Gorges, M.; Vercruysse, P.; Müller, H.P.; Huppertz, H.J.; Rosenbohm, A.; Nagel, G.; Weydt, P.; Petersén, Å.; Ludolph, A.C.; Kassubek, J.; et al. Hypothalamic atrophy is related to body mass index and age at onset in amyotrophic lateral sclerosis. J. Neurol. Neurosurg. Psychiatry 2017, 88, 1033–1041. [Google Scholar] [CrossRef]
- Ngo, S.T.; van Eijk, R.P.A.; Chachay, V.; van den Berg, L.H.; McCombe, P.A.; Henderson, R.D.; Steyn, F.J. Loss of appetite is associated with a loss of weight and fat mass in patients with amyotrophic lateral sclerosis. Amyotroph. Lateral Scler. Front. Degener. 2019, 20, 497–505. [Google Scholar] [CrossRef] [Green Version]
- Garretson, J.T.; Teubner, B.J.; Grove, K.L.; Vazdarjanova, A.; Ryu, V.; Bartness, T.J. Peroxisome proliferator-activated receptor γ controls ingestive behavior, agouti-related protein, and neuropeptide Y mRNA in the arcuate hypothalamus. J. Neurosci. 2015, 35, 4571–4581. [Google Scholar] [CrossRef]
- Lu, M.; Sarruf, D.A.; Talukdar, S.; Sharma, S.; Li, P.; Bandyopadhyay, G.; Nalbandian, S.; Fan, W.; Gayen, J.R.; Mahata, S.K.; et al. Brain PPAR-γ promotes obesity and is required for the insulin-sensitizing effect of thiazolidinediones. Nat. Med. 2011, 17, 618–622. [Google Scholar] [CrossRef] [Green Version]
- Ryan, K.K.; Li, B.; Grayson, B.E.; Matter, E.K.; Woods, S.C.; Seeley, R.J. A role for central nervous system PPAR-γ in the regulation of energy balance. Nat. Med. 2011, 17, 623–626. [Google Scholar] [CrossRef]
- Sarruf, D.A.; Yu, F.; Nguyen, H.T.; Williams, D.L.; Printz, R.L.; Niswender, K.D.; Schwartz, M.W. Expression of peroxisome proliferator-activated receptor-gamma in key neuronal subsets regulating glucose metabolism and energy homeostasis. Endocrinology 2009, 150, 707–712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matias, J.A.; Gilbert, E.R.; Denbow, D.M.; Cline, M.A. Effects of intracerebroventricular injection of rosiglitazone on appetite-associated parameters in chicks. Gen. Comp. Endocrinol. 2017, 246, 99–104. [Google Scholar] [PubMed] [Green Version]
- Banks, W.A.; Kastin, A.J.; Huang, W.; Jaspan, J.B.; Maness, L.M. Leptin enters the brain by a saturable system independent of insulin. Peptides 1996, 17, 305–311. [Google Scholar] [CrossRef]
- Kalra, S.P.; Dube, M.G.; Pu, S.; Xu, B.; Horvath, T.L.; Kalra, P.S. Interacting appetite-regulating pathways in the hypothalamic regulation of body weight. Endocr. Rev. 1999, 20, 68–100. [Google Scholar] [PubMed]
- Kastin, A.J.; Pan, W.; Maness, L.M.; Banks, W.A. Peptides crossing the blood–brain barrier: Some unusual observations. Brain Res. 1999, 848, 96–100. [Google Scholar] [CrossRef]
- Valassi, E.; Scacchi, M.; Cavagnini, F. Neuroendocrine control of food intake. Nutr. Metab. Cardiovasc. Dis. 2008, 18, 158–168. [Google Scholar] [CrossRef] [PubMed]
- Brunetti, L.; Di Nisio, C.; Orlando, G.; Ferrante, C.; Vacca, M. The regulation of feeding: A cross talk between peripheral and central signalling. Int. J. Immunopathol. Pharmacol. 2005, 18, 201–212. [Google Scholar] [CrossRef] [PubMed]
- Giampietro, L.; D’Angelo, A.; Giancristofaro, A.; Ammazzalorso, A.; De Filippis, B.; Fantacuzzi, M.; Linciano, P.; Maccallini, C.; Amoroso, R. Synthesis and structure–activity relationships of fibrate-based analogues inside PPARs. Bioorg. Med. Chem. Lett. 2012, 22, 7662–7666. [Google Scholar] [CrossRef]
- Giampietro, L.; Laghezza, A.; Cerchia, C.; Florio, R.; Recinella, L.; Capone, F.; Ammazzalorso, A.; Bruno, I.; De Filippis, B.; Fantacuzzi, M.; et al. Novel Phenyldiazenyl Fibrate Analogues as PPAR α/γ/δ Pan-Agonists for the Amelioration of Metabolic Syndrome. ACS Med. Chem. Lett. 2019, 10, 545–551. [Google Scholar] [CrossRef]
- Giampietro, L.; Ammazzalorso, A.; Giancristofaro, A.; Lannutti, F.; Bettoni, G.; De Filippis, B.; Fantacuzzi, M.; Maccallini, C.; Petruzzelli, M.; Morgano, A.; et al. Synthesis and Biological Evaluation of 2-Heteroarylthioalkanoic Acid Analogues of Clofibric Acid as Peroxisome Proliferator-Activated Receptor α Agonists. J. Med. Chem. 2009, 52, 6224–6232. [Google Scholar] [CrossRef]
- Giampietro, L.; Ammazzalorso, A.; Amoroso, R.; De Filippis, B. Development of Fibrates as Important Scaffolds in Medicinal Chemistry. ChemMedChem 2019, 14, 1051–1066. [Google Scholar] [CrossRef] [Green Version]
- Ammazzalorso, A.; Amoroso, R.; Bettoni, G.; De Filippis, B.; Fantacuzzi, M.; Giampietro, L.; Maccallini, C.; Tricca, M.L. Candida rugosa lipase-catalysed kinetic resolution of 2-substituted-aryloxyacetic esters with dimethylsulfoxide and isopropanol as additives. Chirality 2008, 20, 115–118. [Google Scholar] [CrossRef]
- Moore, R.A.; Derry, S.; Wiffen, P.J.; Straube, S.; Aldington, D.J. Overview review: Comparative efficacy of oral ibuprofen and paracetamol (acetaminophen) across acute and chronic pain conditions. Eur. J. Pain 2015, 19, 1213–1223. [Google Scholar] [CrossRef] [PubMed]
- Sashidhara, K.V.; Dodda, R.P.; Sonkar, R.; Palnati, G.R.; Bhatia, G. Design and synthesis of novel indole-chalcone fibrates as lipid lowering agents. Eur. J. Med. Chem. 2014, 81, 499–509. [Google Scholar] [CrossRef] [PubMed]
- Amoroso, R.; Leporini, L.; Cacciatore, I.; Marinelli, L.; Ammazzalorso, A.; Bruno, I.; De Filippis, B.; Fantacuzzi, M.; Maccallini, C.; Menghini, L.; et al. Synthesis, Characterization and Evaluation of Gemfibrozil-stilbene Hybrid as Antioxidant Agent. Lett. Drug Des. Discov. 2018, 15, 1230–1238. [Google Scholar] [CrossRef]
- Ammazzalorso, A.; Amoroso, R.; Baraldi, M.; Bettoni, G.; Braghiroli, D.; De Filippis, B.; Giampietro, L.; Tricca, M.L.; Vezzalini, F. Synthesis and antiplatelet activity of thioaryloxyacids analogues of clofibric acid. Eur. J. Med. Chem. 2005, 40, 918–921. [Google Scholar] [CrossRef] [PubMed]
- Giampietro, L.; Gallorini, M.; De Filippis, B.; Amoroso, R.; Cataldi, A.; di Giacomo, V. PPAR-γ agonist GL516 reduces oxidative stress and apoptosis occurrence in a rat astrocyte cell line. Neurochem. Int. 2019, 126, 239–245. [Google Scholar] [CrossRef]
- Orlando, G.; Leone, S.; Ferrante, C.; Chiavaroli, A.; Mollica, A.; Stefanucci, A.; Macedonio, G.; Dimmito, M.P.; Leporini, L.; Menghini, L.; et al. Effects of Kisspeptin-10 on Hypothalamic Neuropeptides and Neurotransmitters Involved in Appetite Control. Molecules 2018, 24, 3071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- di Giacomo, V.; Chiavaroli, A.; Orlando, G.; Cataldi, A.; Rapino, M.; Di Valerio, V.; Leone, S.; Brunetti, L.; Menghini, L.; Recinella, L.; et al. Neuroprotective and Neuromodulatory Effects Induced by Cannabidiol and Cannabigerol in Rat Hypo-E22 cells and Isolated Hypothalamus. Antioxidants 2020, 13, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonvento, G.; Bolaños, J.P. Astrocyte-neuron metabolic cooperation shapes brain activity. Cell Metab. 2021, 33, 1546–1564. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.; Saver, J.L.; Liao, H.W.; Lin, C.H.; Ovbiagele, B. Pioglitazone for Secondary Stroke Prevention: A Systematic Review and Meta-Analysis. Stroke 2017, 48, 388–393. [Google Scholar] [CrossRef]
- Saunders, A.M.; Burns, D.K.; Gottschalk, W.K. Reassessment of Pioglitazone for Alzheimer’s Disease. Front. Neurosci. 2021, 15, 666958. [Google Scholar] [CrossRef]
- Recinella, L.; Leone, S.; Ferrante, C.; Chiavaroli, A.; Shohreh, R.; Di Nisio, C.; Vacca, M.; Orlando, G.; Salvatori, R.; Brunetti, L. Effects of growth hormone-releasing hormone gene targeted ablation on ghrelin-induced feeding. Growth Horm. IGF Res. 2017, 37, 40–46. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chiavaroli, A.; di Giacomo, V.; De Filippis, B.; Cataldi, A.; Ferrante, C.; Giampietro, L. The Small Molecule PPARγ Agonist GL516 Induces Feeding-Stimulatory Effects in Hypothalamus Cells Hypo-E22 and Isolated Hypothalami. Molecules 2022, 27, 4882. https://doi.org/10.3390/molecules27154882
Chiavaroli A, di Giacomo V, De Filippis B, Cataldi A, Ferrante C, Giampietro L. The Small Molecule PPARγ Agonist GL516 Induces Feeding-Stimulatory Effects in Hypothalamus Cells Hypo-E22 and Isolated Hypothalami. Molecules. 2022; 27(15):4882. https://doi.org/10.3390/molecules27154882
Chicago/Turabian StyleChiavaroli, Annalisa, Viviana di Giacomo, Barbara De Filippis, Amelia Cataldi, Claudio Ferrante, and Letizia Giampietro. 2022. "The Small Molecule PPARγ Agonist GL516 Induces Feeding-Stimulatory Effects in Hypothalamus Cells Hypo-E22 and Isolated Hypothalami" Molecules 27, no. 15: 4882. https://doi.org/10.3390/molecules27154882