
Oclacitinib, a Janus Kinase Inhibitor, Reduces the Frequency of IL-4- and IL-10-, but Not IFN-γ-, Producing Murine CD4+ and CD8+ T Cells and Counteracts the Induction of Type 1 Regulatory T Cells


Abstract
:1. Introduction
2. Results
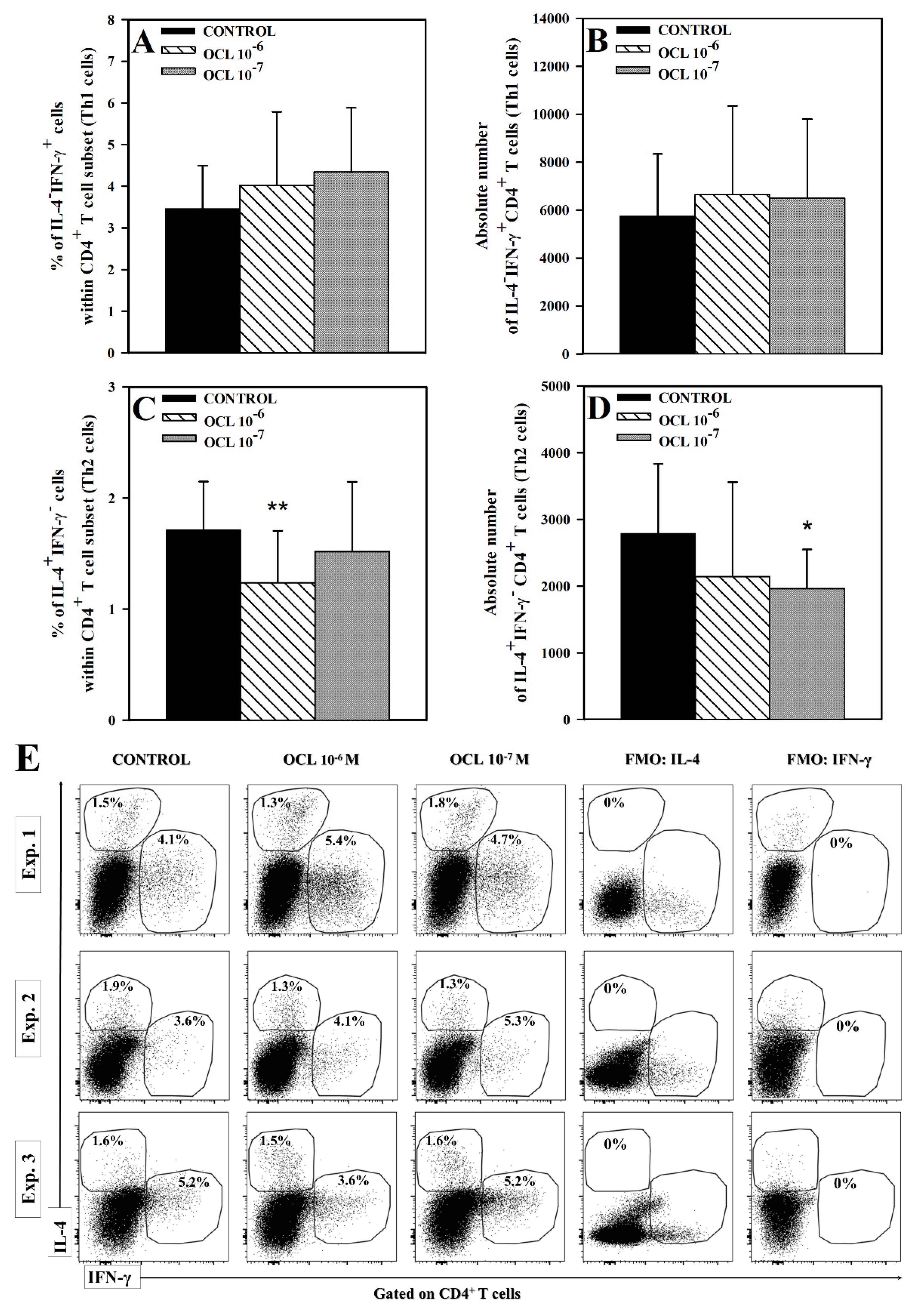
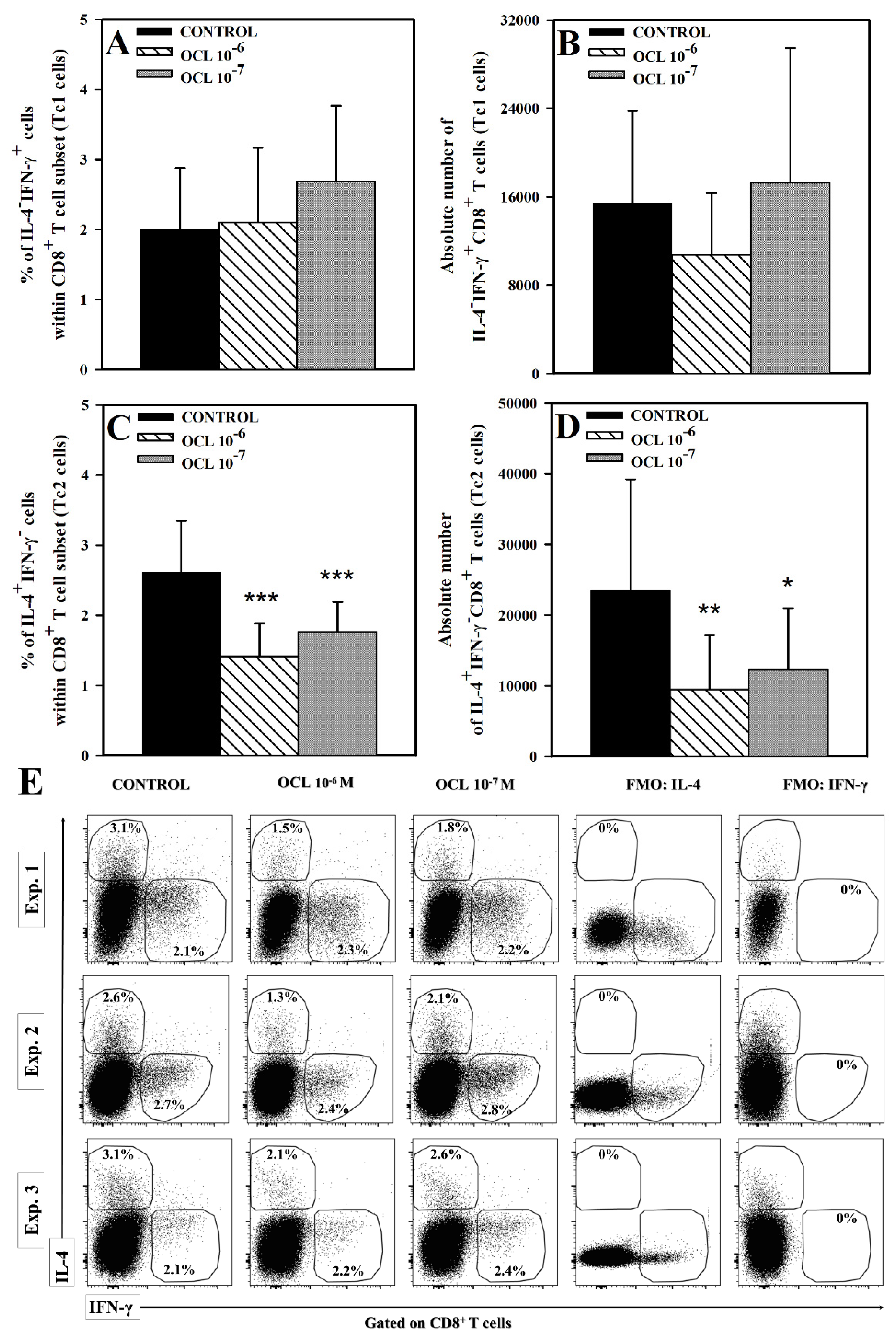
2.1. OCL Reduces the Frequency of IL-4-, but Not IFN-γ- and IL-17-, Producing CD4+ and CD8+ T Cells
2.2. OCL Reduces the Percentage and Absolute Counts of IL-10+CD4+ and IL-10+CD8+ T Cells
2.3. OCL Reduces the Percentage and Absolute Count of CD49b+CD223+CD4+ T Cells
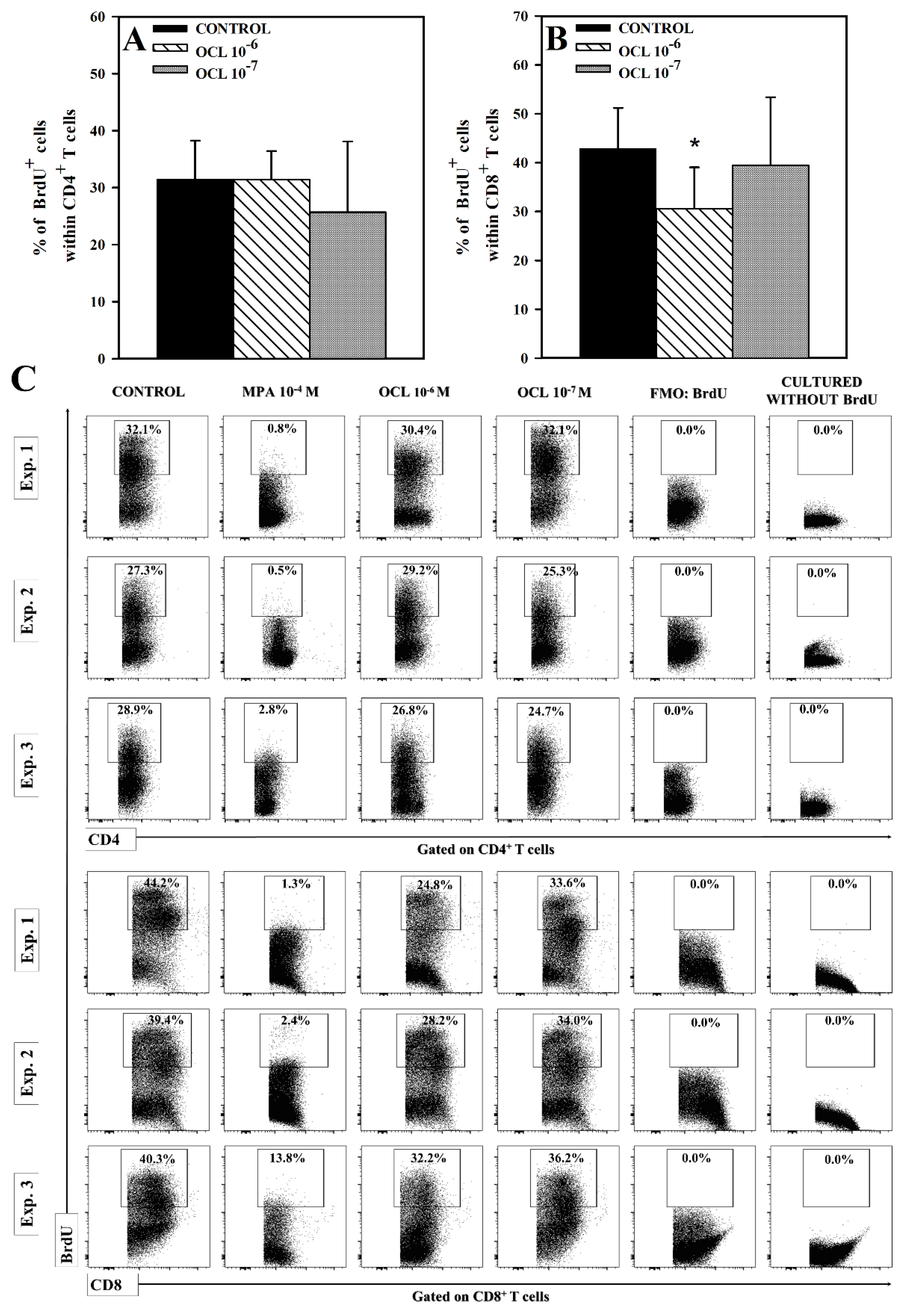
2.4. OCL Reduces the Number of Proliferating CD8+, but Not CD4+, T Cells
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Isolation of Lymphocytes
4.2.1. Spleen
4.2.2. Lungs
4.3. In-Vitro Stimulation and Culture Conditions
4.4. Flow Cytometry
4.4.1. Extracellular Staining
4.4.2. Intracellular Staining for Determination of IL-4-, IL-10-, IL-17- and IFN-γ-Producing Cells and Tr1 Cells
4.4.3. Intracellular Staining for Determination of Proliferating Cells
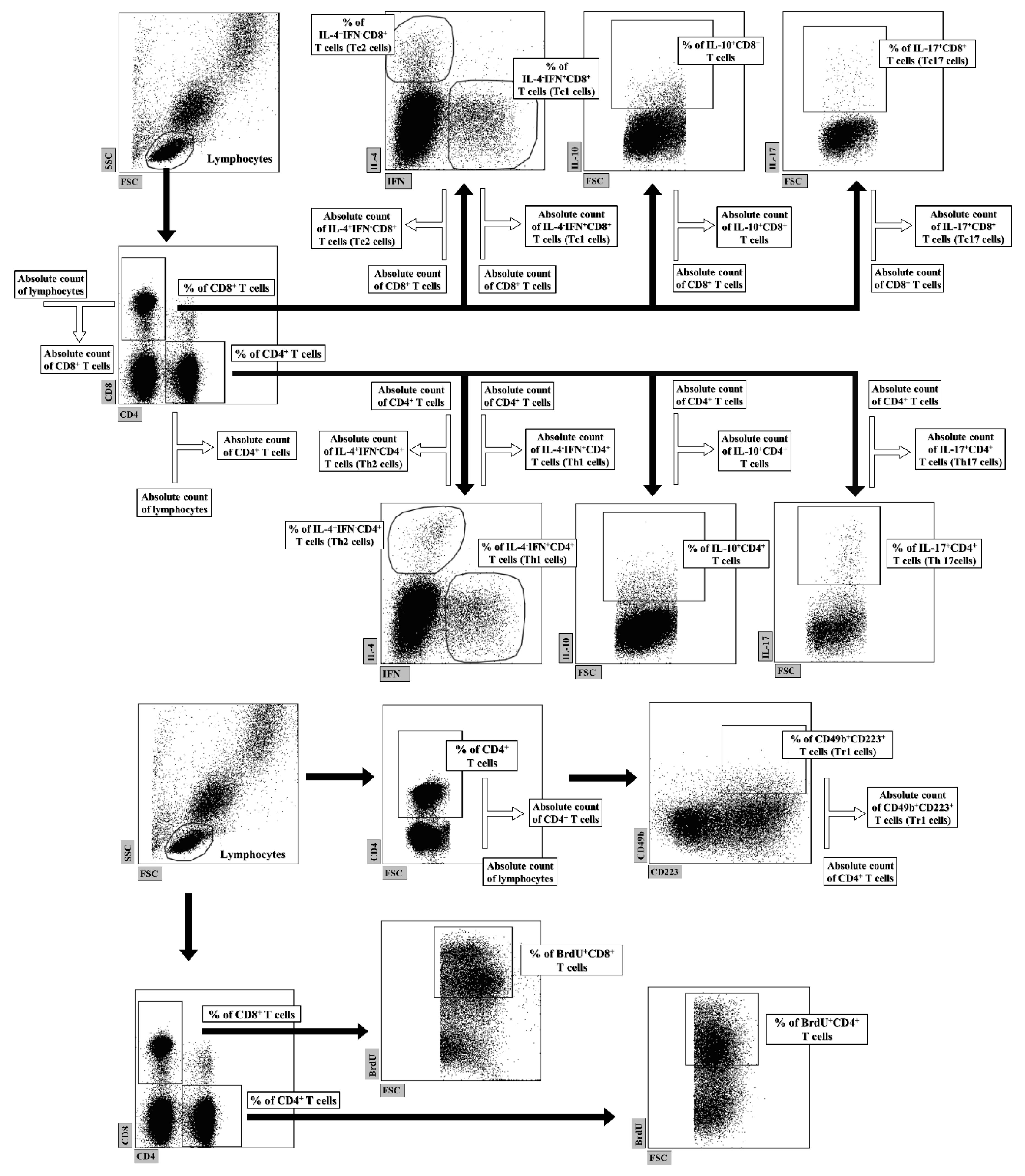
4.5. FACS Acquisition and Data Analysis
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Gonzales, A.J.; Bowman, J.W.; Fici, G.J.; Zhang, M.; Mann, D.W.; Mitton-Fry, M. Oclacitinib (APOQUEL(®)) is a novel Janus kinase inhibitor with activity against cytokines involved in allergy. J. Vet. Pharmacol. Ther. 2014, 37, 317–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiricozzi, A.; Pitocco, R.; Saraceno, R.; Nistico, S.P.; Giunta, A.; Chimenti, S. New topical treatments for psoriasis. Expert Opin. Pharmacother. 2014, 15, 461–470. [Google Scholar] [CrossRef]
- Wang, H.; Brown, J.; Gao, S.; Liang, S.; Jotwani, R.; Zhou, H.; Suttles, J.; Scott, D.A.; Lamont, R.J. The Role of JAK-3 in Regulating TLR-Mediated Inflammatory Cytokine Production in Innate Immune Cells. J. Immunol. 2013, 191, 1164–1174. [Google Scholar] [CrossRef] [Green Version]
- Hodge, J.A.; Kawabata, T.T.; Krishnaswami, S.; Clark, J.D.; Telliez, J.B.; Dowty, M.E.; Menon, S.; Lamba, M.; Zwillich, S. The mechanism of action of tofacitinib—An oral Janus kinase inhibitor for the treatment of rheumatoid arthritis. Clin. Exp. Rheumatol. 2016, 34, 318–328. [Google Scholar] [PubMed]
- Hammitzsch, A.; Chen, L.; de Wit, J.; Al-Mossawi, M.H.; Ridley, A.; Sekine, T.; Simone, D.; Doig, K.; Skapenko, A.; Bowness, P. Inhibiting ex-vivo Th17 responses in Ankylosing Spondylitis by targeting Janus kinases. Sci. Rep. 2018, 8, 15645. [Google Scholar] [CrossRef] [Green Version]
- Ong, P.Y.; Leung, D.Y. Immune dysregulation in atopic dermatitis. Curr. Allergy Asthma Rep. 2006, 6, 384–389. [Google Scholar] [CrossRef]
- Hijnen, D.; Knol, E.F.; Gent, Y.Y.; Giovannone, B.; Beijn, S.J.; Kupper, T.S.; Bruijnzeel-Koomen, C.A.; Clark, R.A. CD8(+) T cells in the lesional skin of atopic dermatitis and psoriasis patients are an important source of IFN-γ, IL-13, IL-17, and IL-22. J. Investig. Dermatol. 2013, 133, 973–979. [Google Scholar] [CrossRef] [Green Version]
- Carmi-Levy, I.; Homey, B.; Soumelis, V. A modular view of cytokine networks in atopic dermatitis. Clin. Rev. Allergy Immunol. 2011, 41, 245–253. [Google Scholar] [CrossRef]
- Gonzales, A.J.; Fleck, T.J.; Humphrey, W.R.; Galvan, B.A.; Aleo, M.M.; Mahabir, S.P.; Tena, J.K.; Greenwood, K.G.; McCall, R.B. IL-31-induced pruritus in dogs: A novel experimental model to evaluate anti-pruritic effects of canine therapeutics. Vet. Dermatol. 2016, 27, 34-e10. [Google Scholar] [CrossRef]
- Arcique, M.A.; Bajwa, J. Atopic dermatitis in humans and dogs. Can. Vet. J. 2020, 61, 82–84. [Google Scholar] [PubMed]
- Olivry, T.; Dean, G.A.; Tompkins, M.B.; Dow, J.L.; Moore, P.F. Toward a canine model of atopic dermatitis: Amplification of cytokine-gene transcripts in the skin of atopic dogs. Exp. Dermatol. 1999, 8, 204–211. [Google Scholar] [CrossRef]
- Nuttall, T.J.; Knight, P.A.; McAleese, S.M.; Lamb, J.R.; Hill, P.B. Expression of Th1, Th2 and immunosuppressive cytokine gene transcripts in canine atopic dermatitis. Clin. Exp. Allergy 2002, 32, 789–795. [Google Scholar] [CrossRef]
- Schlotter, Y.M.; Rutten, V.P.; Riemers, F.M.; Knol, E.F.; Willemse, T. Lesional skin in atopic dogs shows a mixed Type-1 and Type-2 immune responsiveness. Vet. Immunol. Immunopathol. 2011, 143, 20–26. [Google Scholar] [CrossRef]
- Chaudhary, S.K.; Singh, S.K.; Kumari, P.; Kanwal, S.; Soman, S.P.; Choudhury, S.; Garg, S.K. Alterations in circulating concentrations of IL-17, IL-31 and total IgE in dogs with atopic dermatitis. Vet. Dermatol. 2019, 30, 383-e114. [Google Scholar] [CrossRef] [PubMed]
- Arican, O.; Aral, M.; Sasmaz, S.; Ciragil, P. Serum levels of TNF-alpha, IFN-gamma, IL-6, IL-8, IL-12, IL-17, and IL-18 in patients with active psoriasis and correlation with disease severity. Mediat. Inflamm. 2005, 2005, 273–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olivry, T.; Naydan, D.K.; Moore, P.F. Characterization of the cutaneous inflammatory infiltrate in canine atopic dermatitis. Am. J. Dermatopathol. 1997, 19, 477–486. [Google Scholar] [CrossRef] [PubMed]
- Sinke, J.D.; Thepen, T.; Bihari, I.C.; Rutten, V.P.; Willemse, T. Immunophenotyping of skin-infiltrating T-cell subsets in dogs with atopic dermatitis. Vet. Immunol. Immunopathol. 1997, 57, 13–23. [Google Scholar] [CrossRef]
- Majewska, A.; Gajewska, M.; Dembele, K.; Maciejewski, H.; Prostek, A.; Jank, M. Lymphocytic, cytokine and transcriptomic profiles in peripheral blood of dogs with atopic dermatitis. BMC Vet. Res. 2016, 12, 174. [Google Scholar] [CrossRef] [Green Version]
- Boyman, O.; Werfel, T.; Akdis, C.A. The suppressive role of IL-10 in contact and atopic dermatitis. J. Allergy Clin. Immunol. 2012, 129, 160–161. [Google Scholar] [CrossRef]
- Koning, H.; Neijens, H.J.; Baert, M.R.; Oranje, A.P.; Savelkoul, H.F. T cells subsets and cytokines in allergic and non-allergic children. II. Analysis and IL-5 and IL-10 mRNA expression and protein production. Cytokine 1997, 9, 427–436. [Google Scholar] [CrossRef]
- Maeda, S.; Tsuchida, H.; Marsella, R. Allergen challenge decreases mRNA expression of regulatory cytokines in whole blood of high-IgE beagles. Vet. Dermatol. 2007, 18, 422–426. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Chen, G.; Zhang, M.Q.; Xiong, X.Z.; Liu, H.J.; Xin, J.B.; Zhang, J.C.; Wu, J.H.; Meng, Z.J.; Sun, S.W. Imbalance between subsets of CD8(+) peripheral blood T cells in patients with chronic obstructive pulmonary disease. PeerJ 2016, 4, e2301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tousa, S.; Semitekolou, M.; Morianos, I.; Banos, A.; Trochoutsou, A.I.; Brodie, T.M.; Poulos, N.; Samitas, K.; Kapasa, M.; Konstantopoulos, D.; et al. Activin-A co-opts IRF4 and AhR signaling to induce human regulatory T cells that restrain asthmatic responses. Proc. Natl. Acad. Sci. USA 2017, 114, E2891–E2900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chihara, N.; Madi, A.; Karwacz, K.; Awasthi, A.; Kuchroo, V.K. Differentiation and Characterization of Tr1 Cells. Curr. Protoc. Immunol. 2016, 113, 3.27.1–3.27.10. [Google Scholar] [CrossRef] [Green Version]
- Barrat, F.J.; Cua, D.J.; Boonstra, A.; Richards, D.F.; Crain, C.; Savelkoul, H.F.; de Waal-Malefyt, R.; Coffman, R.L.; Hawrylowicz, C.M.; O’Garra, A. In vitro generation of interleukin 10-producing regulatory CD4(+) T cells is induced by immunosuppressive drugs and inhibited by T helper type 1 (Th1)- and Th2-inducing cytokines. J. Exp. Med. 2002, 195, 603–616. [Google Scholar] [CrossRef]
- Zhang, M.; Mathews Griner, L.A.; Ju, W.; Duveau, D.Y.; Guha, R.; Petrus, M.N.; Wen, B.; Maeda, M.; Shinn, P.; Ferrer, M.; et al. Selective targeting of JAK/STAT signaling is potentiated by Bcl-xL blockade in IL-2–dependent adult T-cell leukemia. Proc. Natl. Acad. Sci. USA 2015, 112, 12480–12485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seif, F.; Khoshmirsafa, M.; Aazami, H.; Mohsenzadegan, M.; Sedighi, G.; Bahar, M. The role of JAK-STAT signaling pathway and its regulators in the fate of T helper cells. Cell Commun. Signal. 2017, 15, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Honda, T.; Egen, J.G.; Lämmermann, T.; Kastenmüller, W.; Torabi-Parizi, P.; Germain, R.N. Tuning of antigen sensitivity by T cell receptor-dependent negative feedback controls T cell effector function in inflamed tissues. Immunity 2014, 40, 235–247. [Google Scholar] [CrossRef] [Green Version]
- Jasiecka-Mikołajczyk, A.; Jaroszewski, J.J.; Maślanka, T. Oclacitinib depletes canine CD4+ and CD8+ T cells in vitro. Res. Vet. Sci. 2018, 121, 124–129. [Google Scholar] [CrossRef]
- Carter, L.L.; Murphy, K.M. Lineage-specific requirement for signal transducer and activator of transcription (Stat)4 in interferon gamma production from CD4(+) versus CD8(+) T cells. J. Exp. Med. 1999, 189, 1355–1360. [Google Scholar] [CrossRef]
- Kaplan, M.H.; Wurster, A.L.; Grusby, M.J. A signal transducer and activator of transcription (Stat)4-independent pathway for the development of T helper type 1 cells. J. Exp. Med. 1998, 188, 1191–1196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulling, P.M.; Olson, K.C.; Hamele, C.E.; Toro, M.F.; Tan, S.F.; Feith, D.J.; Loughran, T.P., Jr. Dysregulation of the IFN-γ-STAT1 signaling pathway in a cell line model of large granular lymphocyte leukemia. PLoS ONE 2018, 13, e0193429. [Google Scholar] [CrossRef]
- Yang, X.P.; Ghoreschi, K.; Steward-Tharp, S.M.; Rodriguez-Canales, J.; Zhu, J.; Grainger, J.R.; Hirahara, K.; Sun, H.W.; Wei, L.; Vahedi, G.; et al. Opposing regulation of the locus encoding IL-17 through direct, reciprocal actions of STAT3 and STAT5. Nat. Immunol. 2011, 12, 247–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiricozzi, A.; Maurelli, M.; Peris, K.; Girolomoni, G. Targeting IL-4 for the Treatment of Atopic Dermatitis. Immunotargets Ther. 2020, 9, 151–156. [Google Scholar] [CrossRef]
- Czarnowicki, T.; Gonzalez, J.; Shemer, A.; Malajian, D.; Xu, H.; Zheng, X.; Khattri, S.; Gilleaudeau, P.; Sullivan-Whalen, M.; Suárez-Fariñas, M.; et al. Severe atopic dermatitis is characterized by selective expansion of circulating TH2/TC2 and TH22/TC22, but not TH17/TC17, cells within the skin-homing T-cell population. J. Allergy Clin. Immunol. 2015, 136, 104–115. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, M.H.; Wurster, A.L.; Smiley, S.T.; Grusby, M.J. Stat6-dependent and -independent pathways for IL-4 production. J. Immun. 1999, 163, 6536–6540. [Google Scholar]
- Jankovic, D.; Kugler, D.G.; Sher, A. IL-10 production by CD4+ effector T cells: A mechanism for self-regulation. Mucosal Immunol. 2010, 3, 239–246. [Google Scholar] [CrossRef]
- Tsuji-Takayama, K.; Suzuki, M.; Yamamoto, M.; Harashima, A.; Okochi, A.; Otani, T.; Inoue, T.; Sugimoto, A.; Toraya, T.; Takeuchi, M.; et al. The production of IL-10 by human regulatory T cells is enhanced by IL-2 through a STAT5-responsive intronic enhancer in the IL-10 locus. J. Immunol. 2008, 181, 3897–3905. [Google Scholar] [CrossRef] [Green Version]
- Groux, H.; O’Garra, A.; Bigler, M.; Rouleau, M.; Antonenko, S.; de Vries, J.E.; Roncarolo, M.G. A CD4+ T-cell subset inhibits antigen-specific T-cell responses and prevents colitis. Nature 1997, 389, 737–742. [Google Scholar] [CrossRef]
- Brockmann, L.; Gagliani, N.; Steglich, B.; Giannou, A.D.; Kempski, J.; Pelczar, P.; Geffken, M.; Mfarrej, B.; Huber, F.; Herkel, J.; et al. IL-10 Receptor Signaling Is Essential for TR1 Cell Function In Vivo. J. Immun. 2017, 198, 1130–1141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pot, C.; Apetoh, L.; Awasthi, A.; Kuchroo, V.K. Induction of regulatory Tr1 cells and inhibition of T(H)17 cells by IL-27. Semin Immunol. 2011, 23, 438–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amado, T.; Amorim, A.; Enguita, F.J.; Romero, P.V.; Inácio, D.; de Miranda, M.P.; Winter, S.J.; Simas, J.P.; Krueger, A.; Schmolka, N.; et al. MicroRNA-181a regulates IFN-γ expression in effector CD8 + T cell differentiation. J. Mol. Med. (Berl.) 2020, 98, 309–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lückel, C.; Picard, F.S.R.; Huber, M. Tc17 biology and function: Novel concepts. Eur. J. Immunol. 2020, 50, 1257–1267. [Google Scholar] [CrossRef]
- Gottlieb, S.L.; Martin, D.H.; Xu, F.; Byrne, G.I.; Brunham, R.C. Summary: The natural history and immunobiology of Chlamydia trachomatis genital infection and implications for Chlamydia control. J. Infect. Dis. 2010, 201, S190–S204. [Google Scholar] [CrossRef] [Green Version]
- Banovic, F.; Tarigo, J.; Gordon, H.; Barber, J.P.; Gogal, R.M., Jr. Immunomodulatory in vitro effects of oclacitinib on canine T-cell proliferation and cytokine production. Vet. Dermatol. 2019, 30, 17-e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghoreschi, K.; Jesson, M.I.; Li, X.; Lee, J.L.; Ghosh, S.; Alsup, J.W.; Warner, J.D.; Tanaka, M.; Steward-Tharp, S.M.; Gadina, M.; et al. Modulation of innate and adaptive immune responses by tofacitinib (CP-690,550). J. Immunol. 2011, 186, 4234–4243. [Google Scholar] [CrossRef] [Green Version]
- Collard, W.T.; Hummel, B.D.; Fielder, A.F.; King, V.L.; Boucher, J.F.; Mullins, M.A.; Malpas, P.B.; Stegemann, M.R. The pharmacokinetics of oclacitinib maleate, a Janus kinase inhibitor, in the dog. J. Vet. Pharmacol. Ther. 2014, 37, 279–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Marker | Fluorochrome | Clone | Isotype | Evaluated Parameters |
|---|---|---|---|---|
| CD4 | FITC | H129.19 | IgG2a, κ | The percentage and absolute count of:
|
| CD8a | APC-Cy7 | 53-6.7 | IgG2a, κ | |
| IL-4 | PE-CF594 | 11B11 | IgG1 | |
| IL-10 | APC | JES5-16E3 | IgG2b | |
| IFN-γ | AF700 | XMG1.2 | IgG1,κ | |
| CD4 | FITC | H129.19 | IgG2a, κ | The percentage and absolute count of:
|
| CD8a | APC-Cy7 | 53-6.7 | IgG2a, κ | |
| IL-17 | PerCP-Cy5.5 | TC11-18H10 | IgG1, λ | |
| CD4 | FITC | H129.19 | IgG2a, κ | The percentage and absolute count of:
|
| CD49b | PE-CF594 | DX5 | IgM, κ | |
| CD223 | BB700 | C9B7W | IgG1, κ | |
| CD4 | FITC | H129.19 | IgG2a, κ | The percentage of:
|
| CD8a | APC-Cy7 | 53-6.7 | IgG2a, κ | |
| BrdU | APC | BU1/75 | IgG2a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jasiecka-Mikołajczyk, A.; Jaroszewski, J.J.; Maślanka, T. Oclacitinib, a Janus Kinase Inhibitor, Reduces the Frequency of IL-4- and IL-10-, but Not IFN-γ-, Producing Murine CD4+ and CD8+ T Cells and Counteracts the Induction of Type 1 Regulatory T Cells. Molecules 2021, 26, 5655. https://doi.org/10.3390/molecules26185655
Jasiecka-Mikołajczyk A, Jaroszewski JJ, Maślanka T. Oclacitinib, a Janus Kinase Inhibitor, Reduces the Frequency of IL-4- and IL-10-, but Not IFN-γ-, Producing Murine CD4+ and CD8+ T Cells and Counteracts the Induction of Type 1 Regulatory T Cells. Molecules. 2021; 26(18):5655. https://doi.org/10.3390/molecules26185655
Chicago/Turabian StyleJasiecka-Mikołajczyk, Agnieszka, Jerzy J. Jaroszewski, and Tomasz Maślanka. 2021. "Oclacitinib, a Janus Kinase Inhibitor, Reduces the Frequency of IL-4- and IL-10-, but Not IFN-γ-, Producing Murine CD4+ and CD8+ T Cells and Counteracts the Induction of Type 1 Regulatory T Cells" Molecules 26, no. 18: 5655. https://doi.org/10.3390/molecules26185655